全尺度人工濕地中植物多樣性對(duì)生產(chǎn)力與多樣性效應(yīng)的影響
2014-02-07 06:31:46朱四喜王鳳友張建民蘇春花
生態(tài)環(huán)境學(xué)報(bào) 2014年1期
朱四喜, 常 杰, 葛 瀅, 王 海,, 王鳳友 張建民, 蘇春花
1. 貴州民族大學(xué)化學(xué)與環(huán)境科學(xué)學(xué)院,貴州 貴陽(yáng) 550025;2. 浙江大學(xué)生命科學(xué)學(xué)院,浙江 杭州 310058;3. 紹興文理學(xué)院生命科學(xué)學(xué)院,浙江 紹興 31200;4. 中國(guó)水電顧問集團(tuán)貴陽(yáng)勘測(cè)設(shè)計(jì)研究院,貴州 貴陽(yáng) 550081
全尺度人工濕地中植物多樣性對(duì)生產(chǎn)力與多樣性效應(yīng)的影響
朱四喜1,2*, 常 杰2, 葛 瀅2, 王 海2,3, 王鳳友1, 張建民4, 蘇春花1
1. 貴州民族大學(xué)化學(xué)與環(huán)境科學(xué)學(xué)院,貴州 貴陽(yáng) 550025;2. 浙江大學(xué)生命科學(xué)學(xué)院,浙江 杭州 310058;3. 紹興文理學(xué)院生命科學(xué)學(xué)院,浙江 紹興 31200;4. 中國(guó)水電顧問集團(tuán)貴陽(yáng)勘測(cè)設(shè)計(jì)研究院,貴州 貴陽(yáng) 550081
在中國(guó)東南部的全尺度復(fù)合垂直流人工濕地中開展2年的植物多樣性實(shí)驗(yàn),以研究植物多樣性(包括植物物種豐富度和植物組成)對(duì)群落生產(chǎn)力與多樣性效應(yīng)(即互補(bǔ)效應(yīng)、選擇效應(yīng)和凈多樣性效應(yīng))的影響及其產(chǎn)生機(jī)制。結(jié)果表明,2007年物種豐富度與群落生產(chǎn)力呈線形正相關(guān),而2008年顯著的單峰格局,其關(guān)系式為:y=-0.213x2+3.455x+15.192(R=0.215)。2008年物種豐富度與互補(bǔ)效應(yīng)呈顯著地線形負(fù)相關(guān),而2007年呈單峰格局,其關(guān)系式為:y=-0.389x2+6.974x-10.707(R=0.247),而且2007年與2008年的互補(bǔ)效應(yīng)與生產(chǎn)力都呈顯著的正相關(guān),表明互補(bǔ)效應(yīng)對(duì)生產(chǎn)力的提高有重要作用。然而,2007年與2008年物種豐富度與選擇效應(yīng)之間均沒有顯著相關(guān)性,且選擇效應(yīng)與群落生產(chǎn)力之間也沒有顯著相關(guān)性,表明選擇效應(yīng)對(duì)生產(chǎn)力的提高作用不顯著。2007年與2008年中物種組成對(duì)生產(chǎn)力、互補(bǔ)效應(yīng)、選擇效應(yīng)與凈多樣性效應(yīng)均有顯著影響,說明人工濕地的植物配置對(duì)其生態(tài)系統(tǒng)功能的維持尤為重要。2008年物種豐富度與凈多樣性效應(yīng)呈極顯著地線形負(fù)相關(guān),而2007年呈顯著單峰格局,其關(guān)系式為:y=-0.329 x2+5.968 x-12.659(R=0.234),這種趨勢(shì)主要是由于植物多樣性-生態(tài)系統(tǒng)功能關(guān)系的影響因素(如物種的競(jìng)爭(zhēng)力和生態(tài)位)在2年中有所變化。同時(shí),2007年與2008年的多樣性凈效應(yīng)與生產(chǎn)力都呈顯著正相關(guān)關(guān)系,表明生產(chǎn)力與多樣性凈效應(yīng)的變化趨勢(shì)是同步的。與抽樣效應(yīng)假說不同的是,本實(shí)驗(yàn)中單種最高產(chǎn)物種(蘆竹)在混種時(shí)沒有表現(xiàn)出高產(chǎn),主要是由于生長(zhǎng)的分配、資源的競(jìng)爭(zhēng)力與環(huán)境的變化等。
植物多樣性;人工濕地;多樣性效應(yīng);生產(chǎn)力;植物組成;單峰格局
全球生物多樣性正在急劇下降,物種的消失會(huì)對(duì)生態(tài)系統(tǒng)功能產(chǎn)生損害(Hedlund等,2003;賈力和趙娜,2012;馮劍豐等,2009)。許多研究一直關(guān)注生物多樣性對(duì)生態(tài)系統(tǒng)(如陸地植物群落(多數(shù)為草地群落)、模式微生物生態(tài)系統(tǒng)和水生生態(tài)系統(tǒng))功能的影響(Maestre和Reynolds,2006;鄭克舉等,2013;馬姜明等,2013),但是這些限制了我們對(duì)生態(tài)系統(tǒng)功能作用全面認(rèn)識(shí),所以有必要對(duì)其他類型的自然群落進(jìn)行研究以得到具有更普遍意義上的結(jié)論(Giller等,2004;馬克平,2013)。
許多草地多樣性實(shí)驗(yàn)表明物種豐富度與生產(chǎn)力呈線形正相關(guān)、負(fù)相關(guān)、無(wú)相關(guān),或單峰格局(Huston等,2000;Tilman,1999;Fridley,2001;Wang等,2013;Zhu等,2010),對(duì)以上的解釋主要有2種機(jī)制:選擇效應(yīng)與互補(bǔ)效應(yīng)(Hector,1998;Tilman,1997)。在選擇效應(yīng)占主導(dǎo)的群落中,單個(gè)物種的作用(競(jìng)爭(zhēng)優(yōu)勢(shì)種或關(guān)鍵種)對(duì)系統(tǒng)功能的影響最大(Hector,1998),而在互補(bǔ)效應(yīng)占主導(dǎo)地位的群落中,不同種間的相互促進(jìn)作用對(duì)系統(tǒng)功能的影響更大(Tilman,1997)。然而,選擇效應(yīng)與互補(bǔ)效應(yīng)并不是相互排斥的,有可能它們共同作用影響生產(chǎn)力。
在人工濕地系統(tǒng)中,有關(guān)生物多樣性與生態(tài)系統(tǒng)功能的關(guān)系在近期得到了廣泛研究(Zhu等,2010,2012a,2012b;Zhang等,2010a,2010b)。人工濕地系統(tǒng)提供了重要的生態(tài)系統(tǒng)服務(wù),且它可能依賴于植物多樣性對(duì)生產(chǎn)力與營(yíng)養(yǎng)固持的作用(Zhu等,2010,2012a,2012b)。同時(shí),人工濕地系統(tǒng)在世界范圍內(nèi)其面積在不斷增長(zhǎng),對(duì)于陸地生態(tài)系統(tǒng)的功能與結(jié)構(gòu)具有重要的意義(Zhu等,2010,2012a,2012b;Zhang等,2010a,2010b;Liu等,2009)。因而,本文在中國(guó)東南部的全尺度復(fù)合垂直流人工濕地中,用本地優(yōu)勢(shì)種開展了植物多樣性實(shí)驗(yàn),以研究在高營(yíng)養(yǎng)條件下物種豐富度與物種組成對(duì)群落生產(chǎn)力與生物多樣性效應(yīng)(即互補(bǔ)效應(yīng)、選擇效應(yīng)與凈多樣性效應(yīng))的影響及其產(chǎn)生機(jī)制。
1 材料與方法
1.1 研究樣地與實(shí)驗(yàn)設(shè)計(jì)
實(shí)驗(yàn)樣地位于中國(guó)東南部的浙江省舟山市朱家尖鎮(zhèn)(29o53''N, 122o23'E)的人工濕地,系用于處理朱家尖南沙風(fēng)景區(qū)賓館與餐館等生活污水的全尺度復(fù)合垂直流結(jié)構(gòu)的人工濕地,面積約0.20 hm2。將實(shí)驗(yàn)樣地劃分為164個(gè)小區(qū)(每個(gè)小區(qū)面積約為3.0 m×3.0 m,表1)。
從人工濕地物種庫(kù)中選擇了16種植物,按照與特定生態(tài)系統(tǒng)功能(主要是生產(chǎn)力和養(yǎng)分利用)相關(guān)的植物功能特征,參照Tilman 等(2009)的方法,分為4個(gè)不同功能群,即C3禾草植物(蘆葦Phragmites australis、蘆竹Arundo donax)、C4禾草植物(白茅Imperata cylindrical、荻Triarrhena sacchariflora、芒Miscanthus sinensis、菩提子Coix lacryma-jobi、山類蘆Neyraudia montana、斑茅Saccharum arundinaceum)、豆科植物(杭子梢Campylotropis macrocarpa、馬棘Indigofera pseudotinctoria、胡枝子Lespedeza bicolor、傘房決明Cassia tora)和闊葉草本植物(千屈菜Lythrum salicaria、風(fēng)車草Cyperus alternifolius、美人蕉Canna indica、再力花Thalia dealbata),均為本地優(yōu)勢(shì)種(Zhu等,2010,2012a,2012b)。
1.2 地上生物量的測(cè)定
2007年與2008年9月底,進(jìn)行了植物收割工作。其中,植物樣品的收割樣方設(shè)計(jì)見表1。在每個(gè)小區(qū)中,隨機(jī)收割0.5 m × 0.3 m條塊,留茬10 cm。植物樣品帶回實(shí)驗(yàn)室后,放入烘箱內(nèi),先105 ℃殺青60 min,再65 ℃烘至恒質(zhì)量,取出信封對(duì)各種植物樣品進(jìn)行稱量,得到干質(zhì)量,計(jì)算每個(gè)小區(qū)的群落生產(chǎn)力(即地上生物量, 或簡(jiǎn)稱為生產(chǎn)力)(Zhu等,2010,2012a,2012b)。
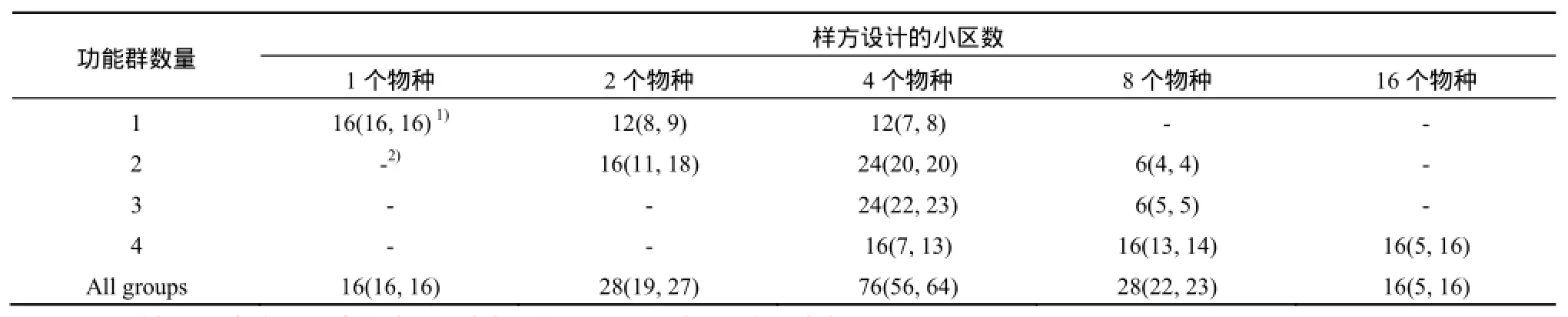
表1 舟山人工濕地多樣性實(shí)驗(yàn)中在2007年與2008年的樣方設(shè)計(jì)Table1 Plots design of the biodiversity experiment in the constructed wetland in Zhoushan in 2007 and 2008
1.3 多樣性效應(yīng)計(jì)算
按照Loreau和Hector的方法(Loreau和Hertor,2001;Lorea,1998;Hector和Hooper,2002),將混播群落中生產(chǎn)力的增加量(凈多樣性效應(yīng)ΔY)分解為選擇效應(yīng)和互補(bǔ)效應(yīng),其計(jì)算公式為:
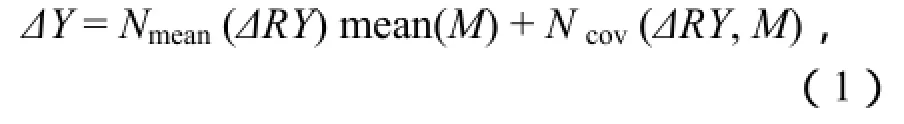
式中,ΔY為凈多樣性效應(yīng),ΔY = Yo-YE,Yo為混播群落的實(shí)際產(chǎn)量,YE為以單播產(chǎn)量為基礎(chǔ)計(jì)算出的混播群落期望產(chǎn)量,M為混播群落中各物種單產(chǎn)的平均產(chǎn)量,ΔRY 為混播群落中各物種相對(duì)產(chǎn)量的變化量(各物種的實(shí)際相對(duì)產(chǎn)量與其期望相對(duì)產(chǎn)量之差),N為混播群落的物種數(shù),Nmean(ΔRY·M)為互補(bǔ)效應(yīng)值,Ncov(ΔRY, M)為選擇效應(yīng)值。式(1)的基本前提是零假設(shè),即多樣性對(duì)生產(chǎn)力無(wú)影響,即ΔY=0,當(dāng)多樣性對(duì)混播群落生產(chǎn)力產(chǎn)生正效應(yīng)時(shí),ΔY > 0,而當(dāng)多樣性對(duì)生產(chǎn)力產(chǎn)生負(fù)效應(yīng)時(shí),則ΔY<0。
1.4 統(tǒng)計(jì)分析
本研究的生物多樣性試驗(yàn)控制了物種豐富度與物種組成這2個(gè)變量;使用SPSS軟件對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析,即利用一般線性模型(General Linear Model)對(duì)數(shù)據(jù)進(jìn)行方差分析(One–way ANOVA),分別按小區(qū)、有無(wú)C3禾草植物、C4禾草植物、豆科植物與闊葉草本植物、物種豐富度、物種組成、生物多樣性效應(yīng)(包括凈多樣性效應(yīng)、互補(bǔ)效應(yīng)與選擇效應(yīng))來分析其效應(yīng)(sequential fitter order)(基于type Ⅲ平方和基礎(chǔ)上;SPSS 16.0, SPSS Inc, Chicago, IL, USA)。同時(shí),差異顯著性用Tukey檢驗(yàn),統(tǒng)計(jì)顯著性α = 0.05,且所有數(shù)據(jù)以均值±標(biāo)準(zhǔn)誤(S.E.)表示。
2 結(jié)果與分析
2.1 植物多樣性與多樣性效應(yīng)的關(guān)系
2007年與2008年中物種組成對(duì)互補(bǔ)效應(yīng)有顯著影響見表2)。2008年物種豐富度與互補(bǔ)效應(yīng)呈顯著地線形負(fù)相關(guān)關(guān)系,而2007年呈單峰格局,其關(guān)系式為:y =-0.389x2+6.974x-10.707(R=0.247)(圖1)。2007年和2008年分別有67.0%與58.5%混種群落中互補(bǔ)作用對(duì)生產(chǎn)力有正效應(yīng),總體上都顯著大于0,且2008年的互補(bǔ)效應(yīng)平均值顯著比2007年的高。另外,2007年與2008年的互補(bǔ)效應(yīng)與生產(chǎn)力都呈顯著正相關(guān)關(guān)系,這表明互補(bǔ)效應(yīng)對(duì)生產(chǎn)力的提高有重要作用(圖1)。
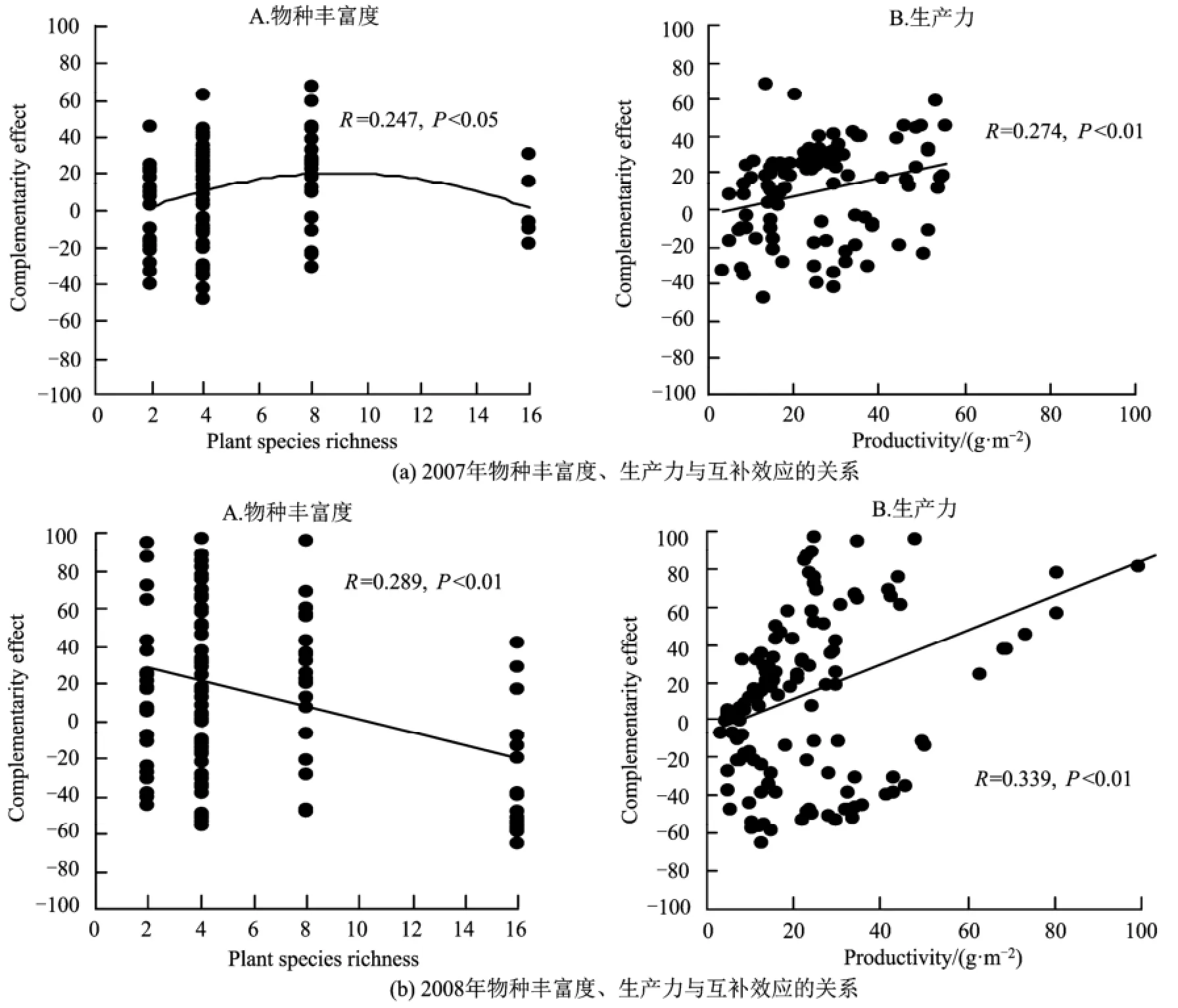
圖1 2007年與2008年中物種豐富度、生產(chǎn)力與互補(bǔ)效應(yīng)的關(guān)系Fig.1 Relationships of plant species richness, productivity and complementaity effects in 2007 and 2008

表2 舟山人工濕地多樣性實(shí)驗(yàn)中多樣性效應(yīng)在2007年與2008年的分析Table2 Analysis biodiversity effects for mixtures of 2007 and 2008 in the biodiversity experiment of the constructed wetland in Zhoushan
2007年與2008年中不同物種組成對(duì)選擇效應(yīng)都有顯著影響(表2),但2007年與2008年選擇效應(yīng)與物種豐富度之間都沒有顯著的相關(guān)性(圖2)。2007年與2008年選擇效應(yīng)小于0的值分別占62.0%與57.7%,總體上都顯著小于0,且2008年的負(fù)選擇效應(yīng)顯著比2007年大(表2)。另外,2007年與2008年的選擇效應(yīng)與群落生產(chǎn)力的相關(guān)性都不顯著,表明選擇效應(yīng)對(duì)生產(chǎn)力的提高作用不明顯(圖2)。總之,2008年負(fù)選擇效應(yīng)顯著比2007年大,而2008年的互補(bǔ)效應(yīng)和凈多樣性效應(yīng)都比2007年顯著小,且2007年凈選擇效應(yīng)總體上顯著大于0,2008年總體上顯著小于0,主要是由于2007年的正互補(bǔ)效應(yīng)的值顯著高于負(fù)選擇效應(yīng)的值,同樣2008年負(fù)選擇效應(yīng)的值也顯著高于正互補(bǔ)效應(yīng)的值(圖1、2,表2)。

圖2 2007年與2008年中物種豐富度、生產(chǎn)力與選擇效應(yīng)的關(guān)系Fig.2 Relationships of plant species richness, productivity and selection effects in 2007 and 2008
2.2 植物豐富度與生產(chǎn)力的關(guān)系
2007年物種豐富度與群落生產(chǎn)力(地上生物量的單位為g·m-2,且地上生物量進(jìn)行平方根轉(zhuǎn)換)關(guān)系呈線形正相關(guān)關(guān)系,而2008年顯著的單峰格局,即生產(chǎn)力隨物種豐富度的增加而增加,當(dāng)物種數(shù)達(dá)到一定水平(8種)后,生產(chǎn)力不再隨物種豐富度的增加而增加,反而呈下降趨勢(shì),其關(guān)系式為:y =-0.213x2+3.455x +15.192(R=0.215)(圖3)。2007年與2008年各多樣性水平內(nèi)群落生產(chǎn)力變化幅度都較大,表明在同一多樣性的水平內(nèi),不同的物種組合對(duì)生產(chǎn)力有顯著影響(表2)。這說明除物種豐富度外,物種的成分對(duì)生態(tài)系統(tǒng)生產(chǎn)力也有重要影響。另外,2007年所有區(qū)塊生物量的平均值為(838±432)g·m-2,2008年則為(922±927)g·m-2,2007年到2008年的生物量有近似顯著的增加趨勢(shì)(表2)。
2007年與2008年中不同物種組成對(duì)多樣性凈效應(yīng)(生物多樣性效應(yīng)的值進(jìn)行平方根轉(zhuǎn)換,但原有的正負(fù)號(hào)保持不變,下同)都有顯著的影響(表2)。2008年物種豐富度與多樣性凈效應(yīng)呈極顯著地線形負(fù)相關(guān)關(guān)系,而2007年物種豐富度與多樣性凈效應(yīng)呈顯著單峰格局,其關(guān)系式為:y=-0.329x2+5.968x-12.659 (R=0.234)(圖4)。另外,2007年與2008年的多樣性凈效應(yīng)與生產(chǎn)力都呈顯著正相關(guān)關(guān)系,表明混種群落的生產(chǎn)力變化趨勢(shì)與多樣性凈效應(yīng)的變化趨勢(shì)是同步的(圖4)。
2.3 抽樣效應(yīng)的驗(yàn)證
本實(shí)驗(yàn)2年中單種最高產(chǎn)的物種(蘆竹)在混種群落中生物量比例都是隨著物種豐富度的增加而顯著降低的(圖5)。因而,本文結(jié)果與抽樣效應(yīng)假預(yù)測(cè)的不同,因?yàn)槌闃有?yīng)假設(shè)認(rèn)為單種最高產(chǎn)物種應(yīng)該在混種時(shí)占優(yōu)勢(shì)。由此,本研究中表現(xiàn)為在大多數(shù)混種群落中沒有超產(chǎn)效應(yīng)。
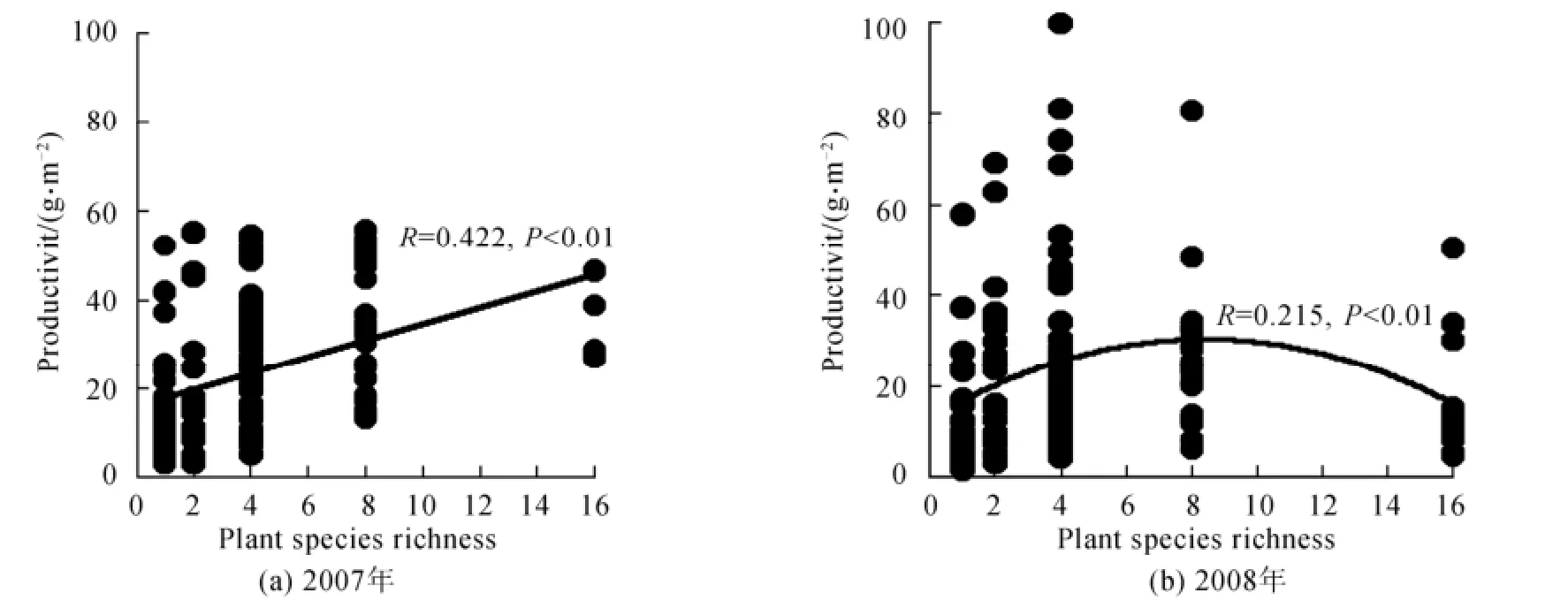
圖3 2007年與2008年中物種豐富度與生產(chǎn)力的關(guān)系Fig.3 Relationships of plant species richness and productivity in 2007 and 2008
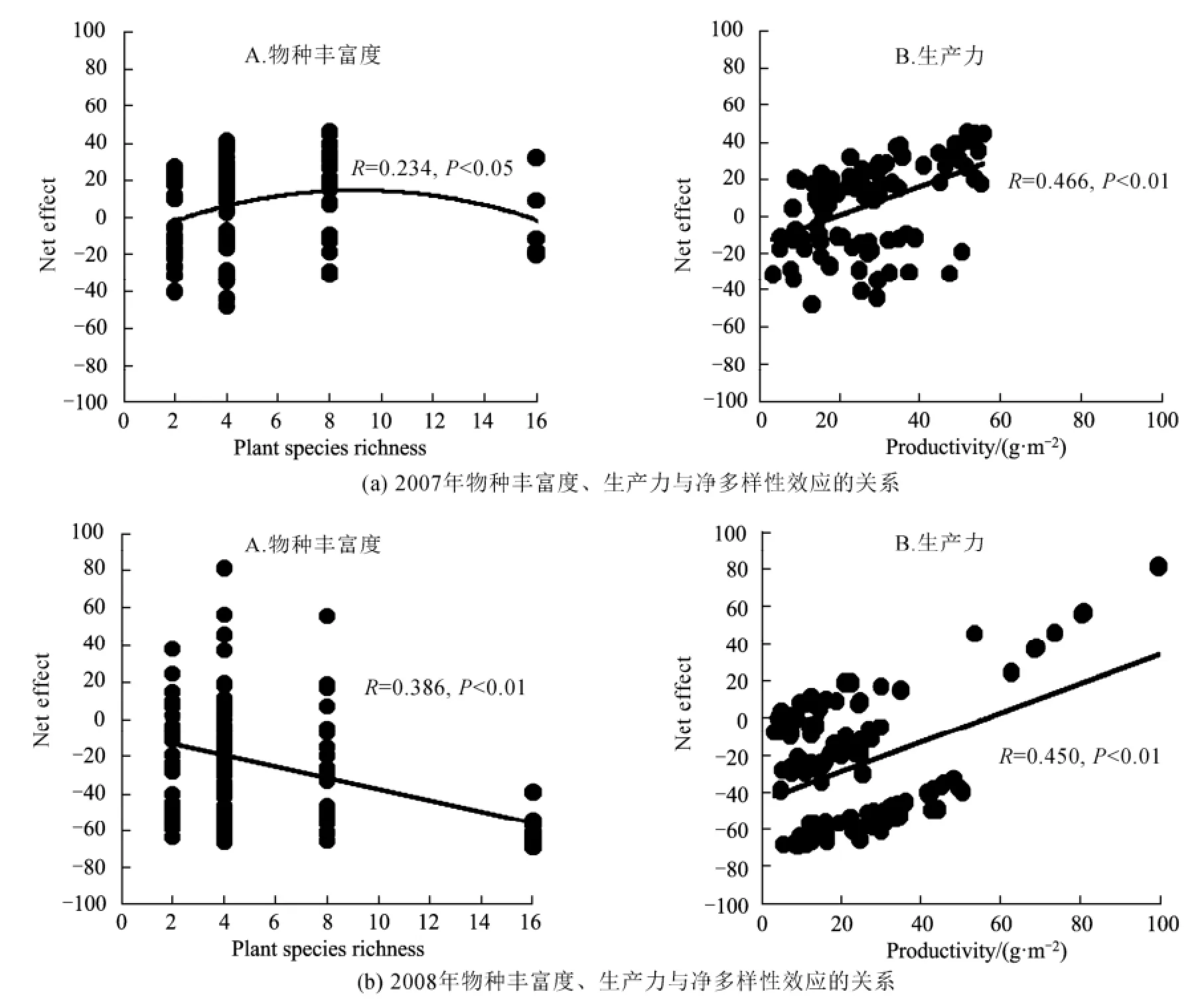
圖4 2007年與2008年中物種豐富度、生產(chǎn)力與凈多樣性效應(yīng)的關(guān)系Fig.4 Relationships of plant species richness, productivity and net diversity effects in 2007 and 2008
3 討論
3.1 物種豐富度對(duì)多樣性效應(yīng)的影響
隨著多樣性的增加,群落的互補(bǔ)效應(yīng)亦應(yīng)線性增加,群落生產(chǎn)力水平應(yīng)不斷提高(Tilman等,1997)。然而,2008年物種豐富度與互補(bǔ)效應(yīng)呈顯著的線形負(fù)相關(guān)。2007年植物物種豐富度與互補(bǔ)效應(yīng)呈顯著的單峰格局,這可能是由于在混種群落中,不僅存在互補(bǔ)效應(yīng),還可能存在其他作用機(jī)理(如競(jìng)爭(zhēng)作用、生化相克作用等)(Hooper,1998)。另外,2007年與2008年的選擇效應(yīng)與生產(chǎn)力的相關(guān)性都不顯著,這可能是由于特殊特種(高生產(chǎn)力的物種)在混種群落中優(yōu)勢(shì)地位因不同的植物組成而表現(xiàn)不同,不能表現(xiàn)出相對(duì)一致的趨勢(shì)(Huston等,2000)。

圖5 2007年與2008年中物種豐富度與最高產(chǎn)物種(蘆竹)在混種群落中生物量比例的關(guān)系Fig.5 Relationships of plant species richness and faction of Ad (Arundo donax) biomass in 2007 and 2008
2008年負(fù)選擇效應(yīng)顯著地比2007年要高,而2008年的互補(bǔ)效應(yīng)與凈多樣性效應(yīng)都比2007年要小(圖1~4,表2),正如Huston等(2000)、Pacala S和Tilman(2002)理論分析的一樣,即關(guān)于從“選擇向互補(bǔ)轉(zhuǎn)型”的理論認(rèn)為群落建立初期,多樣性的作用機(jī)制主要是選擇效應(yīng),但是生態(tài)位互補(bǔ)的作用會(huì)隨時(shí)間的推移而加強(qiáng),并逐步成為主要的多樣性作用機(jī)制。分離加性法結(jié)果也表明,在生長(zhǎng)穩(wěn)定期,互補(bǔ)效應(yīng)為主要的多樣性作用機(jī)制,但關(guān)于這種作用機(jī)制的原因及其作用尚待進(jìn)一步探討(Loreau和Hertor,2001;江小雷等,2004)。
本研究發(fā)現(xiàn),物種豐富度與凈多樣性效應(yīng)的關(guān)系在2007年呈顯著單峰格局,這與Jiang等(2008)的研究結(jié)果一致,而2008年呈顯著地線形負(fù)相關(guān)性,這種變化可能是由于生物多樣性-生態(tài)系統(tǒng)功能關(guān)系的影響因素在2年中有所變化,因?yàn)樯锒鄻有?生態(tài)系統(tǒng)功能關(guān)系的形成取決于物種的競(jìng)爭(zhēng)力和它們對(duì)環(huán)境的影響(物種生態(tài)位)等因素(Jiang等,2008)。
3.2 物種豐富度對(duì)生產(chǎn)力、多樣性-生產(chǎn)力關(guān)系的影響
本文研究發(fā)現(xiàn),2007年物種豐富度與群落的生產(chǎn)力關(guān)系呈顯著的線形正相關(guān)關(guān)系(P<0.01),而2008年呈顯著的單峰格局,這種變化趨勢(shì)與以前有關(guān)生物多樣性研究結(jié)果相一致。Tilman等(2001)對(duì)陸地草本植物群落的研究結(jié)果表明,當(dāng)物種數(shù)為16時(shí),群落的生產(chǎn)力仍然隨物種豐富度升高而上升;然而,歐洲草地實(shí)驗(yàn)結(jié)果表明,當(dāng)物種數(shù)超過8后,群落的生產(chǎn)力反而下降(Tilman,1999;Grace等,2007)。這種“飽和上升型”增長(zhǎng)模式可能意味著到達(dá)了某一較高的物種豐富度水平后,進(jìn)一步增加物種豐富度植物群落生物量將不再增大(Tilman等,1996,1997;Hooper和Vitousek,1997)。因此,本文結(jié)果與在其他大部分實(shí)驗(yàn)結(jié)果是一致的,即多樣性-生產(chǎn)力正相關(guān),但有些實(shí)驗(yàn)的結(jié)果是負(fù)相關(guān)或單峰關(guān)系,從而表明了環(huán)境條件與競(jìng)爭(zhēng)的相互作用對(duì)以上關(guān)系的形成有重要作用(Hector等,2007)。
在本研究中,雖然僅對(duì)多樣性-生產(chǎn)力關(guān)系中的選擇效應(yīng)和互補(bǔ)效應(yīng)做了較為細(xì)致的探討,但并不能據(jù)此而完全否認(rèn)其他多樣性效應(yīng)(如物種共存機(jī)制、種間其他相互作用關(guān)系、冗余種效應(yīng)及競(jìng)爭(zhēng)效應(yīng)等)存在的可能性(江小雷和張衛(wèi)國(guó),2006)。多樣性效應(yīng)的各種作用機(jī)理間存在一定的內(nèi)在聯(lián)系,各種不同的機(jī)理有可能相互重疊,相互影響,生態(tài)系統(tǒng)功能對(duì)物種豐富度的反應(yīng)有可能是多種機(jī)制共同作用的結(jié)果(Roy,2001)。本文在實(shí)驗(yàn)第2年的負(fù)取樣效應(yīng)非常顯著,可能是由于研究時(shí)間相對(duì)較短,正如大多數(shù)有關(guān)多樣性-生產(chǎn)力的研究一樣,實(shí)驗(yàn)時(shí)間是相對(duì)短期(小于3年),往往低估了互補(bǔ)或互促的效應(yīng)。另外,有研究證明施肥有利于高大植物在短期實(shí)驗(yàn)中的競(jìng)爭(zhēng)(Hooper,2004),正如本實(shí)驗(yàn)中蘆竹第2年的株高達(dá)到3~5 m。
3.3 物種組成對(duì)生產(chǎn)力與多樣性效應(yīng)的影響
2007年與2008年中不同物種組成對(duì)多樣性凈效應(yīng)、互補(bǔ)效應(yīng)與選擇效應(yīng)都有顯著影響(表2),這表明,除物種豐富度外,物種(或功能群)的成分對(duì)生產(chǎn)力、養(yǎng)分循環(huán)等過程的影響更大,說明物種的特性是多樣性研究中不可忽視的重要因素(Tilman 等,1997;Jiang 等,2008;Tilman,1998;尤海舟等,2013)。另外,大量有關(guān)生物多樣性-生態(tài)系統(tǒng)功能關(guān)系的研究文獻(xiàn)表明,選擇效應(yīng)比互補(bǔ)效應(yīng)更為普遍,從而說明物種組成要比物種間的相互作用更能影響生態(tài)系統(tǒng)功能(Hooper和Vitousek,1997;岳靜等,2011)。
與抽樣效應(yīng)假說相反的是:本實(shí)驗(yàn)中單種最高產(chǎn)的物種(蘆竹)在混種群落中生物量比例是隨著物種豐富度的增加而顯著降低的(圖5),表明單種高產(chǎn)的物種(蘆竹)在混種時(shí)并不高產(chǎn),其原因可能是由于:生長(zhǎng)的分配與競(jìng)爭(zhēng)、對(duì)不同資源的競(jìng)爭(zhēng)能力、在特定環(huán)境中資源的變化等(Cardinale等,2006;Hector等,2002)。
4 結(jié)論
2年的人工濕地植物多樣性實(shí)驗(yàn)表明:2007年物種豐富度與生產(chǎn)力關(guān)系呈線形正相關(guān),而2008年顯著單峰格局。物種組成對(duì)群落生產(chǎn)力與多樣性效應(yīng)(包括互補(bǔ)效應(yīng)、選擇效應(yīng)、凈多樣性效應(yīng))均有顯著影響;2008年物種豐富度與互補(bǔ)效應(yīng)呈顯著線形負(fù)相關(guān),而2007年呈單峰格局,且2年的互補(bǔ)效應(yīng)與生產(chǎn)力均呈顯著正相關(guān),表明互補(bǔ)效應(yīng)對(duì)生產(chǎn)力的提高有重要作用。然而,2年的選擇效應(yīng)與物種豐富度、生產(chǎn)力都沒有顯著差異,表明選擇效應(yīng)對(duì)生產(chǎn)力的提高作用不顯著。同時(shí),2008年物種豐富度與凈多樣性效應(yīng)呈極顯著線形負(fù)相關(guān),而2007年呈顯著單峰格局。與抽樣效應(yīng)假說不同的是:本實(shí)驗(yàn)中單種最高產(chǎn)的物種(蘆竹)在混種群落中生物量比例與物種豐富度呈顯著負(fù)相關(guān)。研究結(jié)果為植物多樣性與生態(tài)系統(tǒng)功能關(guān)系在人工濕地實(shí)驗(yàn)系統(tǒng)中的應(yīng)用研究提供了數(shù)據(jù)支持,也為人工濕地的科學(xué)管理提供理論依據(jù)。
致謝:本文實(shí)驗(yàn)在濕地植物種植、收割與生產(chǎn)力測(cè)定中,得到葛韓亮、王猛、劉冬、儲(chǔ)子彥、曹海芹等同學(xué)的幫助,在此一并表示感謝。同時(shí),非常感謝澳大利亞昆士蘭大學(xué)李凌教授對(duì)本文中英文摘要的審閱與修改!
CARDINALE B J, SRIVASTAVA D S, DUFFY J E, et al.2006. Effects of biodiversity on the functioning of trophic groups and ecosystems[J]. Nature, 443: 989-992.
FRIDLEY J D.2001. The influence of species diversity on ecosystem productivity: how, where, and why[J] ? Oikos,93: 514-526.
GILLER, P S, Hillebrand H, Berninger U G, et al.2004. Biodiversity effects on ecosystem functioning: emerging issues and their experimental test in aquatic environments[J]. Oikos,104: 423-436.
GRACE J B, ANDERSON T M, SMITH M D, et al.2007.Does species diversity limit productivity in natural grassland communities?[J] Ecology Letters, 10: 680-689.
HECTOR A, BAZELEY-WHITE E, LOREAU M, et al.2002. Overyielding in grassland communities: testing the sampling effect hypothesis with replicated biodiversity experiments[J]. Ecology Letters, 5: 502-511.
HECTOR A, HOOPER R.2002. Darwin and the first ecological experiment[J]. Science, 295:639-640.
HECTOR A, JOSHI J, SCHERER-LORENZEN M, et al.2007. Biodiversity and ecosystem functioning: reconciling the results of experimental and observational studies[J]. Functional Ecology, 21:998-1002.
HECTOR A. 1998.The effect of diversity on productivity: detecting the role of species complementarity[J]. Oikos, 82: 597-599.
HEDLUND K, SANTA REGINA I, VAN DER PUTTEN W H, et al.2003. Plant species diversity, plant biomass and responses of the soil community on abandoned land across Europe: idiosyncracy or above-belowground time lags[J]. Oikos, 103: 45-58.
HOOPER D U, DUKES J S. 2004.Overyielding among plant functional groups in a long-term experiment[J]. Ecology Letters, 7: 95-105.
HOOPER D U, VITOUSEK P M. 1997.The effects of plant composition and diversity on ecosystem processes[J]. Science, 277: 1302-1305.
HOOPER D U.1998. The role of complementarity and competition in ecosystem responses to variation in plant diversity[J]. Ecology, 79: 704-719.
HUSTON M A, AARSSEN K W, AUSTIN M P, et al.2000. No consistent effect of plant diversity on productivity[J]. Science, 289: 1255-1259.
JIANG L, PU Z C, NEMERGUT D R. 2008.On the importance of the negative selection effect for the relationship between biodiversity and ecosystem functioning[J]. Oikos, 117: 488-493.
LIU D, GE Y, CHANG J, PENG C H, et al.2009. Constructed wetlands in China: recent developments and future challenges[J]. Frontier Ecological Environmental, 7: 261-268.
LOREAU M, HERTOR A. 2001.Partitioning selection and complementarity in biodiversity experiments[J]. Nature, 412: 72-76.
LOREAU M. 1998.Separating sampling and other effects in biodiversity experiments[J]. Oikos, 82:600-602.
MAESTRE FT, REYNOLDS J F. 2006.Spatial heterogeneity in soil nutrient supply modulates nutrient and biomass responses to multiple global change drivers in model grassland communities[J]. Global Change Biology, 12: 2431-2441.
PACALA S, TILMAN D. 2002.The transition from sampling to complementarity // KINZIG A, PACALA S, TILMAN D. The functional consequences of biodiversity: empirical progress and theoretical extensions[M]. Princeton: Princeton University Press.
ROY J.2001. How does biodiversity control primary productivity?[M]//ROY J, SAUGIER B, MOONEY H A. Global terrestrial productivity. New York:Academic Press: 169-186.
TILMAN D, KNOPS J, WEDIN D, et al. 1997.The influence of functional diversity and composition on ecosystem processes[J]. Science, 277: 1300-1302.
TILMAN D, LEHMAN C L, BRISTOW C E.1997. Plant diversity and ecosystem productivity: theoretical considerations[J]. Proceedings of the National Academy of Sciences of the United States of America, 94: 1857-1861.
TILMAN D, REICH P B, KNOPS J, et al. 2001.Diversity and productivity in a long-term grassland experiment[J]. Science, 294: 843-845.
TILMAN D, WEDIN D, KNOPS J. 1996.Productivity and sustain ability influenced by biodiversity in grassland ecosystems[J]. Nature, 379: 718-720.
TILMAN D.1998. Plant strategies and the dynamics and structure of plant communities [M]. Princeton: Princeton University Press.
TILMAN D.1999. The ecological consequences of changes in biodiversity: a search for general principles[J]. Ecology, 80: 1455-1474.
WANG H, CHEN Z X, ZHANG X U, et al.2013. Plant species richness increased belowground plant biomass and substrate nitrogen removal in a constructed wetland[J]. Clean-Soil, Air, Water, 41(7): 657-664.
ZHANG C B, WANG J, LIU W L, et al. 2010a.Effects of plant diversity on microbial biomass and community metabolic profiles in a full-scale constructed wetland[J]. Ecological Engineering, 36: 62-68.
ZHANG C B, WANG J, LIU W L, et al.2010b. Effects of plant diversity on nutrient retention and enzyme activities in a full-scale constructed wetland[J]. Bioresource Technology, 101: 1686-1692.
ZHU S X, CHANG J, GE Y, et al.2012a. Positive effects of plant species diversity on substrate nitrogen mineralization in a full-scale constructed wetland[J]. Fresenius Environmental Bulletin, 21(6b): 1689-1694.
ZHU S X, GE H L, GE Y, et al.2010. Effects of plant diversity on productivity and substrate nitrogen in a subsurface vertical flow constructed wetland[J]. Ecological Engineering, 36(10): 1307-1313.
ZHU S X, ZHANG P L, WANG H, et al.2012b. Plant species richness affected nitrogen retention and ecosystem productivity in a full-scale onstructed wetland[J]. Clean-Soil, Air, Water, 40(4): 341-347.
馮劍豐,李宇,朱琳. 2009.生態(tài)系統(tǒng)功能與生態(tài)系統(tǒng)服務(wù)的概念辨析[J].生態(tài)環(huán)境學(xué)報(bào), 18(4): 1599-1603.
賈力, 趙娜. 2012.生物多樣性和生態(tài)系統(tǒng)功能的研究歷史與現(xiàn)狀[J]. 內(nèi)蒙古科技與經(jīng)濟(jì), 10: 46-50.
江小雷, 張衛(wèi)國(guó), 嚴(yán)林, 等. 2004.植物群落物種多樣性對(duì)生態(tài)系統(tǒng)生產(chǎn)力的影響[J]. 草業(yè)學(xué)報(bào), 13(6): 8-13.
江小雷, 張衛(wèi)國(guó). 2006.一年生植物群落內(nèi)取樣效應(yīng)和互補(bǔ)效應(yīng)的分離[J].生態(tài)學(xué)報(bào), 26(6): 1896-1902.
馬姜明, 吳蒙, 占婷婷, 等. 2013.漓江流域巖溶區(qū)檵木群落不同恢復(fù)階段物種組成及多樣性變化[J]. 生態(tài)環(huán)境學(xué)報(bào), 22(1): 66-71.
馬克平. 2013.生物多樣性與生態(tài)系統(tǒng)功能的實(shí)驗(yàn)研究[J]. 生物多樣性, 21 (3): 247-248.
尤海舟, 葛春, 王超, 等. 2013.冀東鐵礦2 種恢復(fù)模式植被演替過程中群落結(jié)構(gòu)及生物多樣性比較研究[J]. 生態(tài)環(huán)境學(xué)報(bào), 22(9): 1482-1487.
岳靜, 郝敏, 張靈菲, 等. 2011.物種多樣性-生態(tài)系統(tǒng)功能關(guān)系的影響因素[J]. 草業(yè)科學(xué), 28(4): 532-535.
鄭克舉,唐旭利,張靜, 等. 2013.季風(fēng)常綠闊葉林演替系列菌根資源及其與群落多樣性的關(guān)系[J]. 生態(tài)環(huán)境學(xué)報(bào),22(5): 729-738.
Effects of plant diversity on productivity and diversity effect in a full-scale constructed wetland
ZHU Sixi1,2*, CHANG Jie2, GE Ying2, WANG Hai2,3, WANG Fengyou1, ZHANG Jianmin4, SHU Chunhua1
1. College of Chemistry and Environmental Science, Guizhou Minzu University, Guiyang 550025, China; 2. College of Life Science, Zhejiang University, Hangzhou 310058, China; 3. College of Life Science, Shaoxing University, Shaoxing 312000, China; 4. Hydrochina Guiyang Engineering Corporation, Guiyang 550081, China
An experiment was conducted in a full-scale constructed wetland in Southeastern China between 2007 and 2008 to study the effects of plant diversity (including plant species richness and plant compositions) on the community productivity (such as complementary effects, selection effects and net diversity effects). The results show a strong, positive correlation between the community productivity and plant species richness in 2007. With a unimodal pattern, the community productivity correlated poorly with the species richness in 2008 (R=0.215 for the fitted relationship of y=-0.213x2+3.455x+15.192). This indicates that the interaction of environmental conditions and competition played an important role in the formation of the above relationships. The plant species richness was found to correlate negatively with complementary effects in 2008, but a unimodal pattern was evident in 2007 (R=0.247 for the fitted relationship of y=-0.389 x2+6.974 x-10.707). Meanwhile, a significant, positive relationship between complementary effects and productivity show that complementary effects played an important role in the improvement of community productivity in both 2007 and 2008. In contrast, no significant relationship found among selection effects and plant species richness and productivity, suggesting that selection effects had no important role in the improvement of community productivity in 2007 and 2008. Plant composition had significant effects on the productivity as well as complementary effects, selection effects and net diversity effects in 2007 and 2008. This indicates that the plant configuration in the full-scale constructed wetland was particularly important for maintaining the functionality of the ecosystem. In addition, the plant species richness was found to correlate negatively with net diversity effects in 2008, but such a correlation was absent in 2007 and instead a unimodal pattern was evident (R=0.234 for the fitted relationship y=-0.329 x2+5.968 x-12.659). The different correlation patterns were due to variations of various factors that influenced the biodiversity-system functioning in the two years, such as competitiveness of different species and ecological niche. No consistent relationship between selection effects and productivity was found, suggesting that the variation trends of net diversity effects and productivity were synchronous. Inconsistent with the sampling effect hypothesis, the species (Arundo donax) with the highest monoculture yield did not produce higher yield in the mixtures, probably due to growing allocation, resources competition and environmental changes. The results from this study provide data support to the research on and application of the relationship between plant diversity and ecosystem functioning in constructed wetland experiments, and also offer a theoretical base for scientific management of constructed wetland systems.
plant diversity; constructed wetlands; diversity effect; productivity; plant c omposition; unimodal pattern
Q948.12
A
1674-5906(2014)01-0035-08
朱四喜, 常杰, 葛瀅, 王海, 王鳳友, 張建民, 蘇春花. 全尺度人工濕地中植物多樣性對(duì)生產(chǎn)力與多樣性效應(yīng)的影響[J]. 生態(tài)環(huán)境學(xué)報(bào), 2014, 23(1): 35-42.
ZHU Sixi, CHANG Jie, GE Ying, WANG Hai, WANG Fengyou, ZHANG Jianmin, SHU Chunhua. Effects of plant diversity on productivity and diversity effect in a full-scale constructed wetland [J]. Ecology and Environmental Sciences, 2014, 23(1): 35-42.
教育部新世紀(jì)人才支持計(jì)劃項(xiàng)目(NCET-12-0659);國(guó)家自然科學(xué)基金(31270377);貴州省優(yōu)秀科技教育人才省長(zhǎng)資金項(xiàng)目(黔省專合字[2012]71號(hào));貴陽(yáng)市低碳科技計(jì)劃項(xiàng)目(筑科合同[2012205]號(hào));貴州省科技廳項(xiàng)目(黔科合J字[2011]2363)
朱四喜(1972年生),男,副教授,博士,碩士生導(dǎo)師,主要從事濕地生態(tài)學(xué)研究。E-mail: zhusixi2011@163.com
2013-08-24
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:18:18
核科學(xué)與工程(2021年4期)2022-01-12 06:30:26
今日農(nóng)業(yè)(2020年19期)2020-12-14 14:16:52
科學(xué)大眾(中學(xué))(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
中學(xué)物理·高中(2016年12期)2017-04-22 11:53:03
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學(xué)周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學(xué)周刊·少年版(2015年1期)2015-07-07 17:15:12
