芋頭軟腐病拮抗菌的篩選及生防效果研究
2014-01-18 09:18:52趙江濤趙延存黃開紅李鵬霞王毓寧胡花麗
食品科學 2014年7期
趙江濤,趙延存,黃開紅,李鵬霞*,王毓寧,胡花麗
(江蘇省農業科學院農產品加工研究所,江蘇 南京 210014)
芋頭軟腐病拮抗菌的篩選及生防效果研究
趙江濤,趙延存,黃開紅,李鵬霞*,王毓寧,胡花麗
(江蘇省農業科學院農產品加工研究所,江蘇 南京 210014)
分別從蔬菜田土壤和采后芋頭表面黏附土壤中分離純化到267和244株菌株,通過平板拮抗實驗,發現其中6個菌株對胡蘿卜軟腐歐文氏菌(Erwinia carotovora subsp. carotovora,Ecc)產生了清晰且明顯的拮抗圈(直徑>10 mm)。利用芋頭對它們的生物防控效果進行復篩,結果發現BGP14菌株發酵液處理芋頭的發病率僅為6.7%,顯著低于其他5個菌株(P<0. 05)。根據BGP14的形態學特征、芽孢產生及16S rRNA和gyrB基因的部分核酸序列,該菌株被鑒定為解淀粉芽孢桿菌BGP14(Bacillus amyloliquefaciens)。BGP14在芋頭傷口上具有較強的定殖能力,并將病原菌Ecc的群體數量壓制在一個較低水平。BGP14與病原菌Ecc聯合處理芋頭的苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)活性顯著高于其他處理。以上結果表明,解淀粉芽孢桿菌BGP14對病原菌E. carotovora具有強大的拮抗活性,能夠有效防治貯藏期芋頭軟腐病,具有潛在的應用開發前景。
胡蘿卜軟腐歐文氏菌;解淀粉芽孢桿菌;進化樹;群體動態;系統誘導抗病性
芋頭(Colocasia esculenta (L.) Schoot)屬于天南星科芋屬植物,其地下塊莖富含淀粉和各種微量礦物元素,具有藥食兩用的功效,在我國長江流域及華南地區廣泛種植,是人們所喜愛的一種健康食品[1-2]。每年10—11月份采收芋頭塊莖,進行貯藏,以便長期供應市場。在保藏過程中,由于芋頭呼吸旺盛、濕度高、傷口多,容易引起病原菌的侵染,從而腐爛變質。其中,由胡蘿卜軟腐歐文氏菌(Erwinia carotovora subsp. carotovora,Ecc)引起軟腐病是導致貯藏期芋頭腐爛變質的主要原因之一。
目前,貯藏期芋頭軟腐病的防治主要依靠化學殺菌劑和抗生素,例如次氯酸鹽、農用鏈霉素、代森銨等[3]。近年來,隨著人們食品安全意識的提高,化學殺菌劑和抗生素被嚴格限制或禁止在采后農產品上應用,這就促使研究者尋求更加安全的采后農產品病害防治替代技術。基于微生物的生物防控技術被認為是未來貯藏期農產品病害控制的主要發展方向之一[4-5],例如枯草芽孢桿菌、羅倫隱球酵母等[6-8]。
本實驗擬從容易發生軟腐病的環境中分離篩選對Ecc具有較高拮抗活性的生防菌株,評估其對貯藏期芋頭軟腐病的生防效果,初步分析其可能的作用模式。
1 材料與方法
1.1 樣品采集與菌株分離
樣品分別來源于江蘇省農業科學院蔬菜田表層土壤和新采收的芋頭表面黏附土壤,每種樣品隨機收集200 g,晾干、混勻。每種土壤樣品取10 g,分別加入到含有100 mL滅菌水的250 mL錐形瓶中,在200 r/min、30 ℃的條件下孵育4 h。利用蒸餾水梯度稀釋,涂布2YT(胰蛋白胨17 g/L、酵母提取物 10 g/L、NaCl 5 g/L,pH 7.0)瓊脂平板,將平板在30 ℃條件下培養16 h。從平板上挑取單菌落,在2YT平板上劃線純化培養2次,將純化后的單菌落劃線保存在2YT平板上待用。
1.2 生防菌的初步篩選
將純化的單菌落在2YT液體培養基中培養,培養條件為150 r/min、30 ℃、36 h,培養液備用。將病原菌Ecc(本研究室從貯藏期芋頭上分離鑒定并保存)在2YT液體培養基中培養,培養條件為150 r/min、30 ℃、12 h。將培養好的Ecc菌液按照1%的體積比加入到液態的2YT瓊脂培養基中(溫度低于40 ℃),迅速混勻、倒平板。凝固后利用滅菌打孔器在每個平板上均勻打6個孔(直徑為4 mm),在每個孔內滴加待驗證生防菌菌液40 μL。將處理好的平板放置在30 ℃培養24 h,然后調查每個菌株的拮抗圈直徑,以未加生防菌菌液的孔作為空白對照組。每個菌株接種3個孔,相同的實驗重復了3次。
1.3 生防效果的活體評價
根據初篩的結果,選擇拮抗圈直徑大于10 mm的菌株進行生防效果的活體評價。挑取具有較高拮抗活性的菌株在2YT液體培養基中培養,培養條件為30 ℃、150 r/min、36 h。取上述培養液40 mL、10 000 r/min條件下離心30 min,將上清液經0.22 μm細菌過濾器過濾后備用。另外,挑取病原菌Ecc單菌落于2YT液體培養基中,在30 ℃、150 r/min條件下培養12 h,利用滅菌蒸餾水稀釋50倍備用。實驗用芋頭是從江蘇省靖江市購買的香沙芋,選擇新鮮、大小基本一致、無損傷的健康香沙芋,利用次氯酸鈉溶液(體積比0.1%的有效氯)浸泡1 min,然后用自來水沖洗、晾干。將芋頭的柄掰斷1~2 cm,產生新的創傷斷面。將芋頭的創傷斷面分別浸沒在拮抗菌發酵液或上清中10 s,對照處理樣品浸泡在清水中,在室溫條件下風干2 h。然后在每個斷面接種Ecc菌液5 μL。將處理好的芋頭放置在塑料保鮮箱內,22 ℃條件下孵育,每天觀察芋頭的發病情況,于第10天調查芋頭的發病率和腐爛情況。病斑面積大于0.2 cm2時,定義為發病。每個處理設置3個重復,每個重復包含10個芋頭。
1.4 拮抗菌株BGP14的鑒定
1.4.1 菌株的形態觀察
將菌株在2YT平板上劃線培養,30 ℃條件下培養1~3 d,連續觀察菌株在2YT平板上的菌落形態。挑取菌體,按照改良的Schaeffer和Fulton氏染色法觀察菌體形態和芽孢的形成[9]。
1.4.2 根據16S rRNA和gyrB基因序列進行分子鑒定
基于生防菌BGP14菌株的16S rRNA和gyrB基因部分序列,對其進行分子鑒定。利用基因組DNA提取試劑盒(天根生化科技(北京)有限公司)提取BGP14的基因組DNA。利用16S rRNA通用引物fd2(5’-AGAGTTTGATCATGGCTCAG-3’)和rp1(5’-ACGGTTACCTTGTTACGACTT-3’)擴增其部分序列[10],將PCR產物測序。利用gyrB的通用引物UP-1(5’-GAAGTCATCATGACCGTTCTGCAYGC NGGNGGNAARTTYGA-3’)和UP-2r(5’-AGCAGG GTACGGATGTGCGAGCCRTCNACRTCNGCRTCN GTCAT-3’)擴增其部分序列,利用測序引物UP-1S(5’-GAAGTCATCATGACCGTTCTGCA-3’)和UP-2Sr(5’-AGCAGGGTACGGATGTGCGAGCC-3’)對PCR產物進行測序[11]。基于獲得的16S rRNA、gyrB及GenBank下載的參考序列,利用ClustalX 1.83軟件分別進行多序列分析[12]。然后,利用MEGA 5.05軟件分別構建基于16S rRAN和gyrB基因的遺傳進化樹[13],選擇Pseudomonas fl uorescens SBW25(NC_012660.1)作為遺傳進化樹的外圍參照菌株。
1.5 拮抗菌BGP14和病原菌Ecc在芋頭傷口上的群體動態
按照1.3節的描述接種BGP14和Ecc,將接種好的芋頭按照1.3節的條件進行孵育。分別于接種后0、1、3、6、10 d,調查BGP14和Ecc在芋頭傷口上的群體動態[14]。在每個調查時間點,每個處理隨機取4個芋頭,利用滅菌手術刀將芋頭傷口及腐爛部分切下來,放置到滅菌坩堝內,添加2~3 mL滅菌水研磨勻漿,最后利用滅菌水定容至15 mL。將研磨好的樣品梯度稀釋,分別涂布2YT平板(調查BGP14菌落數)和含有100 μg/mL利福平的2YT平板上。30 ℃條件下孵育16 h,測定Ecc菌落數量。每個處理設置3個重復,每個重復接種25個芋頭。
1.6 苯丙氨酸解氨酶和過氧化物酶活性的動態測定
按照1.3節的描述接種BGP14和Ecc,將接種好的芋頭按照1.3節的條件進行孵育。分別于接種后0、1、3、6、10 d取樣。在每個調查時間點,每個處理隨機取5個芋頭。切取與接種傷口及腐爛病斑鄰近的組織,切成約5 mm×5 mm的小塊,用液氮速凍,保存在-70 ℃備用。實驗開始時,每個處理取冷藏的樣品5 g,加入到10 mL 200 mmol/L磷酸鈉緩沖液(pH 6.4)中,再添加0.5 g聚乙烯吡咯烷酮。利用勻漿機勻漿,然后在4 ℃、10 000 r/min離心40 min,上清被用來測定酶活力。參照文獻Zhao Yancun等[14]的描述方法測定芋頭的苯丙氨酸解氨酶(L-phenylalanin ammonia-lyase,PAL)活性。
參照湯章城[15]的描述方法測定過氧化物酶(peroxidase,POD)活性,根據芋頭的特點進行了一些改動。反應混合液包括0.5 mL的粗提酶液和2 mL愈創木酚(100 mmol/L磷酸鈉,pH 6.4,10~50 mmol/L愈創木酚,現配現用),在30 ℃條件下孵育5 min。然后加入1 mL雙氧水(H2O2,24 mmol/L),測定470 nm波長處的吸光度變化。POD酶活力單位定義:每微克蛋白中每分鐘吸光值上升0.01個單位定義為1個酶活力單位(1 U)。
1.7 統計分析
所有實驗數據是3個重復實驗結果的平均值。利用Excel 2003和SPSS 13.0軟件對實驗結果進行統計分析,采用Tukey’s測驗進行差異顯著性分析(P<0.05)。
2 結果與分析
2.1 拮抗菌的分離和初步篩選
分別從蔬菜田土壤和芋頭表面黏附土壤分離純化了267和244株菌,對其進行平板拮抗實驗,結果發現3株(BGP14、BGP67、BGP109)來源于蔬菜田土壤和3株(YT38、YT103、YT231)來源于芋頭表面土壤的細菌在含有病原菌Ecc的2YT平板上產生了清晰且明顯的拮抗圈(直徑>10 mm)(圖1)。其中,BGP14對Ecc的拮抗圈直徑最大,為12.2 mm,但是該菌株與其他5個菌株之間差異不顯著(P>0.05)。

圖1 生防菌對病原菌Ecc的拮抗活性Fig.1 Inhibition zones of antagonistic bacteria against the pathogen Ecc
2.2 拮抗菌生防效果的活體評價
為了進一步評估上述6個拮抗菌株的實際生防效果,分別測定了它們的發酵菌液和無菌上清對貯藏期芋頭軟腐病發病率的影響。如圖2所示,發酵菌液處理芋頭的發病率在6.7%~40.0%,均顯著低于對照(90.0%)(P<0.05),其中BGP14處理的發病率顯著低于YT38、YT103、BGP67和BGP109處理,僅為6.7%;無菌上清液處理芋頭的發病率在36.7%~60.0%之間,對貯藏期芋頭軟腐病也具有一定防治效果,但是明顯高于相應發酵菌液處理芋頭的發病率。以上實驗結果表明,拮抗菌BGP14的發酵菌液對貯藏期芋頭軟腐病具有良好生防效果。
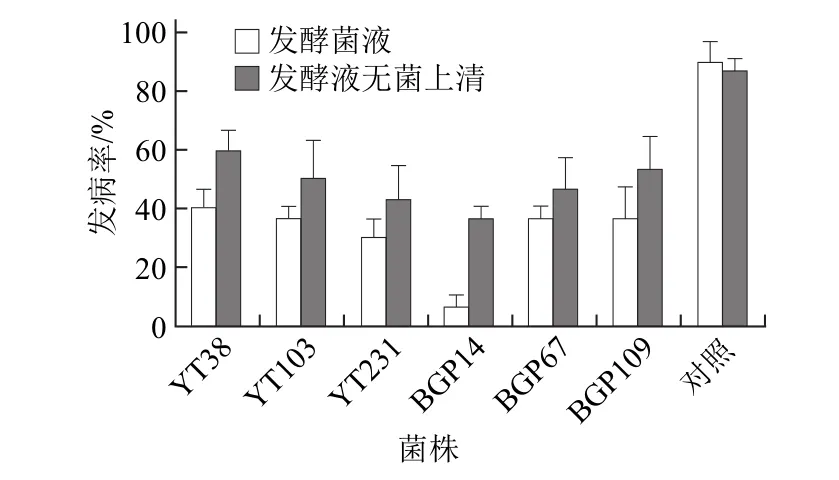
圖2 拮抗菌對貯藏期芋頭軟腐病生防效果的活體評價Fig.2 in vivo screening of potential antagonists against the postharvest soft rot of taros
2.3 BGP14的分類鑒定
拮抗菌BGP14在2YT培養基平板上的菌落形態為乳白色,表面皺縮、菌體為桿狀、形成芽孢。根據以上特征,初步判定為芽孢桿菌。為了進一步確定BGP14的分類地位,利用16S rRNA和gyrB基因序列對其進行了分子鑒定。
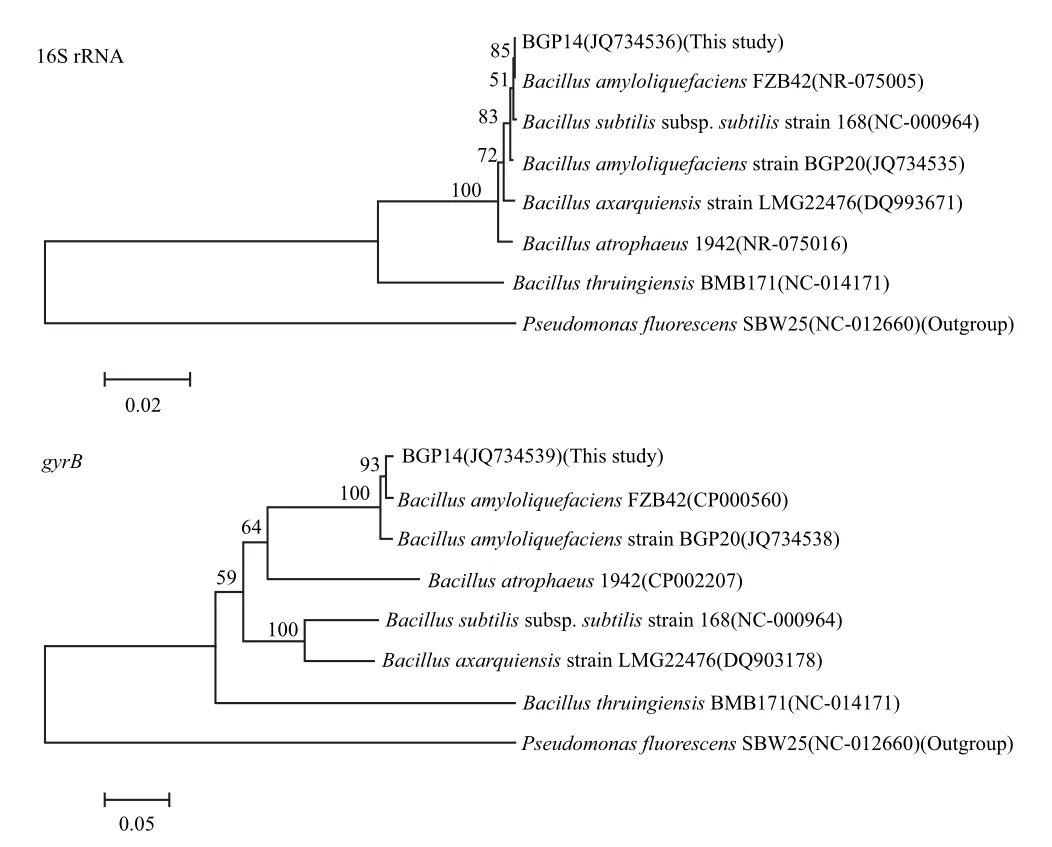
圖3 基于16S rRNA和gyrB基因部分序列構建BGP14的兩個進化樹Fig.3 Two phylogenetic trees of the antagonist BGP14 based on the partial nucleotide sequences of 16S rRNA and gyrB gene
利用通用引物,我們成功獲得了1 6 S r R NA(1432 bp)和gyrB(1179 bp)的部分序列,并提交到GenBank數據庫,序列登陸號分別為JQ734536(16S rRNA)和JQ734539(gyrB)。利用鄰接法(Neighbour-Joining)分別構建了基于16S rRNA和gyrB序列的進化樹如圖3所示。在這兩個進化樹中,拮抗菌株BGP14都與Bacillus amyloliquefaciens FZB42聚類在一起。對這兩個菌株的16S rRNA和gyrB基因序列分別進行BLASTn分析,結果發現它們的核酸序列同源性分別為99.9%和99.1%。以上結果表明,BGP14菌株屬于Bacillus amyloliquefaciens。
2.4 BGP14和Ecc在芋頭傷口上的群體動態
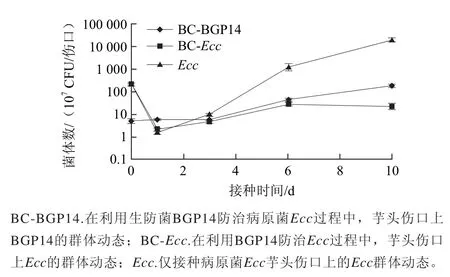
圖4 拮抗菌BGP14和病原菌EccEcc在芋頭傷口上的群體動態Fig.4 Population dynamics of the antagonist BGP14 and the pathogen Ecc in postharvest taro wounds
由圖4可知,在接種后24 h,在僅接種病原Ecc的對照處理和聯合接種生防菌BGP14及Ecc的生防處理芋頭傷口上,Ecc的群體數量分別下降99.1%和99.3%。隨后,對照處理芋頭傷口上Ecc的群體數量迅速上升,接種后第10天的菌體密度達到2.0×1011CFU/傷口;但是,在生防處理的芋頭傷口上,病原菌Ecc增殖緩慢,接種后第10天的菌體密度為2.3×108CFU/傷口,僅為對照處理的0.1%。上述結果表明,生防菌BGP14顯著抑制了Ecc的增殖,從而達到對芋頭軟腐病的防控。另外,在生防菌與病原菌互作的初期階段,BGP14的群體密度維持在一個相對恒定的水平,此后緩慢增加。在接種后第10天,BGP14的群體密度達到1.8×109CFU/傷口,是初始接種量的35.6倍。
2.5 PAL和POD的活性
PAL和POD是與誘導抗病性緊密相關的酶。圖5A表明,在創傷接種處理后,各處理組和對照組芋頭的PAL活性均下降,第3天下降到最低點,隨后上升。在整個實驗過程中,利用生防菌BGP14和病原菌E. carotovora處理芋頭的PAL活性始終高于僅接種E. carotovora的芋頭,大部分時間內也高于清水對照處理。以上結果表明,在生物防治過程中,BGP14誘導芋頭的PAL活性顯著上升。
圖5B表明,病原菌E. carotovora能夠誘導芋頭POD活性大幅上升,接種后第6天達到最高點,是初始水平的1.93倍,然后迅速下降;在先后利用生防菌BGP14和病原菌E. carotovora處理的芋頭上,POD活性迅速上升,在接種后第3天達到最高值,是初始水平的1.73倍,然后迅速下降。滅菌水對照處理芋頭的POD活性變化幅度較小。
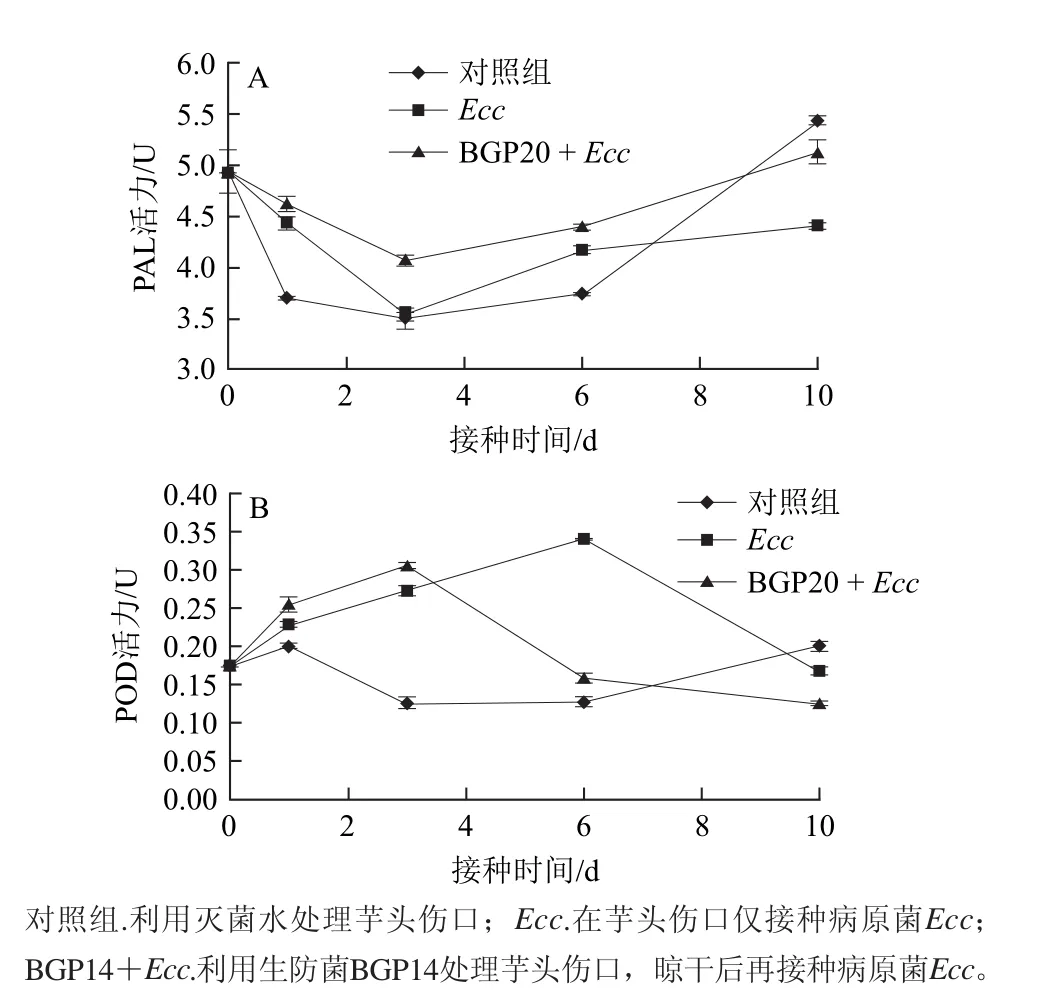
圖5 芋頭PAL和POD活性的誘導表達Fig.5 Induction of phenylalanine ammonia-lyase (PAL) and peroxidase (POD) activities in postharvest taros
3 討論與結論
胡蘿卜軟腐歐文氏菌(Ecc)能夠引起多種天南星科植物軟腐病,例如魔芋、芋頭等,大量相關研究主要集中在田間病害的生物防治[16-17]。截止目前,通過文獻檢索還沒有發現針對貯藏期芋頭軟腐病生物防治的研究報道。
病害的生物防治是一個生防菌、病原菌與寄主互作的復雜過程,并且該過程受到環境因素的影響。通過實驗室內的平板拮抗實驗,本研究從蔬菜田土壤和芋頭表面黏附土壤中共獲得6株對病原菌Ecc具有較強拮抗活性的生防菌,但是,這并不表示它們對實際病害具有良好生防效果[18]。de Costa等[19]通過平板拮抗實驗從香蕉種植園分離獲得32株對香蕉炭疽病菌(Colletotrichum musae)具有顯著拮抗活性的細菌,但是僅有4株拮抗菌在生產上對香蕉炭疽病表現出良好生防效果。通過活體評價方式,本實驗發現僅BGP14菌株的發酵菌液對芋頭軟腐病具有較好生防效果,處理芋頭的發病率僅為6.7%,其他5個菌株處理的發病率為30%~40%。
大量研究表明,生防菌主要通過拮抗次生代謝產物的產生、對寄主傷口營養和空間的競爭或誘導寄主抗病性等方式抑制或殺死病原菌,從而阻止或延緩病害的發生及發展[4,20]。本實驗發現,BGP14發酵液無菌上清對芋頭軟腐病的防效為63.3%,但是其發酵菌液的生防效果高達93.3%,這表明BGP14菌體細胞和其產生的抗菌次生代謝產物在防治芋頭軟腐病過程中都發揮了重要作用。通過對生防菌BGP14和病原菌Ecc在芋頭傷口上的群體競爭動態分析,也發現BGP14在芋頭傷口上具有較強的適應能力和增殖能力,并且將Ecc的群體數量壓制在一個較低水平,從而阻止和延緩病害的發生及發展。Demoz等[21]研究也發現在利用枯草芽孢桿菌(Bacillus subtilis)防治鱷梨果莖腐病的過程中,該生防菌在鱷梨的花朵上具有較強定殖能力。
誘導系統抗病性(induced systemic resistance,ISR)是一些芽孢桿菌(Bacillus spp.)的重要生防機制之一,它能夠誘導寄主病程相關蛋白(pathogenesis related proteins)的表達,例如PAL和POD[14,22]。本實驗發現,相對于病原菌Ecc,BGP14能夠顯著提高芋頭的PAL活性,較高的PAL活性可能對Ecc的增殖具有一定的抑制作用。另外,Ecc處理誘導芋頭POD活性大幅上升,并且在中后期顯著高于BGP14和Ecc聯合處理的芋頭。在處理的第3~10天,芋頭的POD活性與BGP14的群體數量呈負相關。根據病原菌及生防菌在芋頭傷口上的群體動態及其誘導的PAL和POD活性變化,本研究表明,生防菌BGP14對芋頭的PAL和POD具有一定的耐受性,但是病原菌Ecc對芋頭的誘導性防衛反應比較敏感。
本實驗從芋頭表面黏附土壤中分離獲得一株解淀粉芽孢桿菌(Bacillus amyloliquefaciens)BGP14,該菌株的發酵液能夠有效防治貯藏期芋頭軟腐病,具有潛在的應用開發前景。為了推動生防菌BGP14在生產上的應用,還需對該菌株的生物學特性及生防機制進行深入研究。
[1] 許曉春, 林朝朋, 朱定和. 不同包裝處理對切分芋頭貨架期的影響[J].食品研究與開發, 2008, 29(1): 137-140.
[2] 王毓寧, 胡花麗, 李鵬霞, 等. 不同殺菌劑對紅香芋貯藏品質及抗氧化能力的影響[J]. 食品與發酵工業, 2012, 38(11): 207-210.
[3] 陳利鋒, 徐敬友. 農業植物病理學[M]. 北京: 中國農業出版社, 2001: 300-303.
[4] DROBY S, WISNIEWSKI M, MACARISIN D, et al. Twenty years of postharvest biocontrol research: is it time for a new paradigm[J]. Postharvest Biology and Technology, 2009, 52: 137-145.
[5] SHARMA R R, SINGH D, SINGH R. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: a review[J]. Biological Control, 2009, 50: 205-221.
[6] ARREBOLA E, SIVAKUMAR D, BACIGALUPO R, et al. Combined application of antagonist Bacillus amyloliquefaciens and essential oils for the control of peach postharvest diseases[J]. Crop Protection, 2010, 29: 369-377.
[7] 劉海波, 田世平, 秦國政, 等. 羅倫隱球酵母對葡萄采后病害的拮抗效果[J]. 中國農業科學, 2002, 35(7): 831-835.
[8] 張旭, 尚楠, 張寶, 等. 抗瓜果腐霉芽孢桿菌優良菌株的篩選及生物學特性[J]. 食品科學, 2012, 33(5): 138-143.
[9] 趙斌, 何紹江. 微生物學實驗[M]. 北京: 科學出版社, 2002: 42-43.
[10] WEISBURG W G, BARNS S M, PELLETIER D A, et al. 16S ribosomal DNA amplification for phylogenetic study[J]. Journal of Bacteriology, 1991, 173: 697-703.
[11] YAMAMOTO S, HARAYAMA S. PCR amplification and direct sequencing of gyrB genes with universal primers and their application to the detection and taxonomic analysis of Pseudomonas putida strains[J]. Applied and Environmental Microbiology, 1995, 61: 1104-1109.
[12] THOMPSON J D, GIBSON T J, PLEWNIAK F, et al. The Clustal_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools[J]. Nucleic Acids Research, 1997, 24: 4876-4882.
[13] TAMURA K, PETERSON D, PETERSON N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Molecular Biology and Evolution, 2011, 28: 2731-2739.
[14] ZHAOh ao Yancun, LI Pengxia, HUANG Kaihong, et al. Control of postharvest soft rot caused by Erwinia carotovora of vegetables by a strain of Bacillus amyloliquefaciens and its potential modes of action[J]. World Journal of Microbiology and Biotechnology, 2013, 29: 411-420.
[15] 湯章城. 現代植物生理學實驗指南[M]. 北京: 科學出版社, 1999: 317-318.
[16] 姬廣海, 吳亞鵬, 白學慧, 等. 抗生素溶桿菌對魔芋軟腐病和根際微生物多樣性的影響[J]. 江西農業大學學報, 2009, 31(3): 499-503; 544.
[17] 張麗輝, 王永吉, 廖林, 等. 生防菌06-4對魔芋軟腐病的防治及機理的初步研究[J]. 湖南農業大學學報, 2011, 37(3): 286-289.
[18] NALLY M C, PESCE V M, MATURANO Y P, et al. Biocontrol of Botrytis cinerea in table grapes by non-pathogenic indigenous Saccharomyces cerevisiae yeasts isolated from viticultural environments in Argentina[J]. Postharvest Biology and Technology, 2012, 64: 40-48.
[19] de COSTA D M, ZAHRA A R F, KALPAGE M D, et al. Effectiveness and molecular characterization of Burkholderia spinosa, a prospective biocontrol agent for controlling postharvest diseases of banana[J]. Biological Control, 2008, 47: 257-267.
[20] 馬桂珍, 付泓潤, 王淑芳, 等. 海洋多黏類芽孢桿菌L1-9菌株生產抑菌物質發酵條件及其抑菌譜的研究[J]. 食品科學, 2012, 33(19): 231-235.
[21] DEMOZ B T, KORSTEN L. Bacillus subtilis attachment, colonization, and survival on avocado flowers and its mode of action on stem-end rot pathogens[J]. Biological Control, 2006, 37: 68-74.
[22] IPPOLITO A, GHAOUTH A E, WILSON C L, et al. Control of postharvest decay of apple fruit by Aureobasidium pullulans and induction of defense responses[J]. Postharvest Biology and Technology, 2000, 19: 265-272.
Screening of Antagonistic Strain and Its Biocontrol Ability against Postharvest Soft Rot of Taro Caused by Erwinia carotovora subsp. carotovora
ZHAO Jiang-tao, ZHAO Yan-cun, HUANG Kai-hong, LI Peng-xia*, WANG Yu-ning, HU Hua-li
(Institute of Agro-Product Processing, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China)
In this study, 267 and 244 strains were isolated from the vegetable farm soil and the adherent soil on postharvest taros, respectively. Six strains generated clear and obvious antagonistic zones (diameter > 10 mm) on 2YT agar plates with Erwinia carotovora subsp. carotovora (Ecc). in vivo assays showed that the BGP14 cell culture was the most effective in controlling the postharvest soft rot of taros among six antagonistic strains tested. The incidence of soft rot in taros treated by the BGP14 cell culture was only 6.7%, which was significantly lower than that of those treated by five other strains (P < 0.05). Based on morphology, sporulation, and partial nucleotide sequences of 16S rRNA and gyrB gene, the isolate BGP14 was identified as Bacillus amyloliquefaciens. The antagonist BGP14 had a stronger colonizing ability in taro woundas compared to the pathogen Ecc in the process of biological control, and the viable count of Ecc was suppressed to a very low level. In addition, the treatment with both BGP14 and Ecc induced a higher activity of phenylalanine ammonia-lyase (PAL) than with others in taros. In conclusion, the present study demonstrated that the isolate BGP14 showed highly antagonistic activity against Ecc, and could effectively control the postharvest soft rot of taros, thus possessing potential application prospect.
Erwinia carotovora subsp. carotovora; Bacillu s amyloliquefaciens; phylogenetic tree; population dynamics; induced systemic resistance
S476;Q939
A
1002-6630(2014)07-0155-05
10.7506/spkx1002-6630-201407031
2013-04-12
江蘇省農業科技自主創新資金項目(CX(11)2063);江蘇省博士后基金項目(2011年第2批)
趙江濤(1973—),男,副研究員,碩士,研究方向為食品科學。E-mail:zhjt@jaas.ac.cn
*通信作者:李鵬霞(1976—),女,副研究員,博士,研究方向為果蔬采后保鮮與加工。E-mail:lpx213@126.com
