北京西山地區大山雀與其它鳥類種群種間聯結分析
2013-12-21 05:00:02董大穎范宗驥李扎西姐陳卓琳王敏增王奇峰關文彬
生態學報 2013年20期
關鍵詞:物種
董大穎,范宗驥,李扎西姐,陳卓琳,王敏增,王奇峰,李 凱,關文彬,*
(1.北京林業大學,北京 100083;2.北京市西山試驗林場,北京 100093)
種間聯結是指不同物種在空間分布上的相互關聯性,通常是由于群落生境的差異影響了物種的分布為引起的[1],是用來解釋群落結構的重要數量特征之一[2]。引起種間聯結有4種原因:(1)相似與不相似的環境需求;(2)一個種為另一個種創造了定居條件或者前者對后者施加了壓力;(3)兩者在競爭資源中相互排斥造成負聯結;(4)兩者在根系中通過物理的或化學的因素相互影響[3]。研究群落的種間聯結性,能夠有效地反映各物種在群落中的分布情況,各物種對環境因子的適應程度以及物種在特定環境因子作用下的種間相互關系,有助于進一步認清群落的結構、類型、及群落的演替趨勢[4]。
在植物生態研究上已經在森林[5-6]、草原[7]、沼澤[8]、灌叢[9]、荒漠[10]等多種類型的植物群落中廣泛的應用;微生物群落方面,對鳥類巢穴內的菌群進行了種間聯結分析,同時,通過菌落的實例細致評價了不同種間聯結測度的應用[11];在昆蟲方面,種間聯結運用于西雙版納聚果榕榕果小蜂的種間關系研究,發現其在昆蟲生態學中同樣可以清楚的反映群落內物種間的相互關系[12]。可見,種間聯結應用范圍很廣,同時,在種群的動態、群落結構、演替方面具有重要意義,但種間聯結在鳥類群落的研究卻沒有。
鳥類群落的物種組成并不是隨機的,有許多因素影響著群落的結構組成[13]。但是,鳥類群落組成機制的研究一直受到鳥類流動、遷徙等鳥類空間不固定特征的困擾。如,鳥類群落物種是否存在種間聯結?種間聯結的尺度如何界定?存在種間聯結的話,如何檢驗與測度?導致的原因是什么?因此,本研究以北京西山側柏古樹林及其周邊針闊混交林、落葉闊葉林為對象,試圖通過改進鳥類調查方法,引入表聯結的檢驗與聯結度測度的公式,以森林鳥類廣布種大山雀為目標種,探討鳥類群落種間聯結的研究方法,結合鳥類的營巢、取食行為集團[14-16]、生態位[17]等研究,分析鳥類群落的種間聯結的關系。以豐富鳥類群落學研究的科學方法,進而為鳥類及其棲息地的保護提供科學依據。
1 研究方法
1.1 研究地概況
北京靜福寺位于海淀區四季青鄉許木蘭陀山麓,西山試驗林場的中部,北緯40°00',東經116°11',海拔232 m,年平均溫度為11.8℃,年平均降水量為680 mm,土壤以砂質土為主,多石礫,山體坡度在15°—35°之間。該地區植被類型屬于暖溫帶落葉闊葉林,植被主要包括人工針葉林和次生落葉闊葉林,喬木以側柏(Platycladus orientalis)、刺槐(Robinia pseudoacacia)、構樹(Broussonetia papyrifera)等為主,灌木則以荊條(Vitex negundo)、扁擔桿(Grewia biloba)、酸棗(Ziziphus jujuba)等為主。
1.2 鳥類調查方法
調查方法采用固定樣帶法月連續全天定位,植物群落樣帶(面積300 m×10 m)始于北緯40°00'01.4″東經116°11'10.3″終到北緯 40°00'13″東經 116°10'48.9″。鳥類觀測范圍是以 25 m 為半徑的圓。調查時間從 2012年3月到2013年1月,每月進行2次調查,每日調查時間冬季從7:00—16:00,其余季節從6:00—18:00。記錄用8×42倍雙筒望遠鏡或裸眼見到和耳聽到的鳥類實體種類、數量、棲息環境等,主要依據《中國鳥類野外手冊》辨別種類。將遇見率≥1只/h作為優勢種,0.1—1只/h作為常見種,小于0.1只/h作為稀有種。
1.3 群落種間聯結的總體檢驗
總體相關性檢驗采用方差比率法[18]:按照方差比率法來檢驗多物種間的關聯可說明在某地出現的多物種間是否存在顯著的關聯性,先假設群落物種間無顯著關聯,按下列公式計算檢驗統計量:

式中,S為總的物種數,N為總樣方數,Ti為樣方j內出現的研究物種總數,ni為物種i出現的樣方數,t為樣方中的平均數。在獨立性假設條件下VR期望值為1。VR=1表示復合種間關聯性是獨立的;VR>1表示物種間表現出正的關聯;VR<1表示物種間存在負關聯。采用統計量W=N×VR來檢驗VR值偏離1的顯著程度,若種間無關聯,則W落入卡平方分布給出的界限概率為
種間聯結度多采用2×2列聯表給出的定性數據來測度[1]:

表1 2×2列聯表Table 1 2×2 contingency table
1.4 種間聯結的檢驗
χ2檢驗 用連續性校正[19]公式計算,這個校正公式考慮了邊緣頻率的變化[20-21]:

式中,n為取樣總數,當χ2<3.841時,種間關聯性不強;當χ2≥6.635時,種間有顯著的生態聯結性。同時,用ad-bc的正負作為種間聯結性的判斷標準,值大于零為正聯結,反之則為負聯結。
1.5 種間聯結度的計算
(1)點相關系數PCC指數[22]
采用變化幅度由+1通過0而達到-1的有中心指數,表示種間的聯結程度,PCC為負值時表示為負聯結,其絕對值越大負聯結度越高;PCC為正值時表現正聯結,其值越大正聯結度越高。

(2)成對物種間匹配系數
選用以下2個無負值變化的于0與1幅度變化的無中心指數(noncent redindices)來表示種間關聯程度和相伴出現的機率,在“無關聯”時等于0,在“最大關聯”時為1。
共同出現百分率[23],也稱作物種的群落系數[24]:


2 結果與分析
北京西山地區鳥類群落調查共65個鳥類物種,總體相關性檢驗,鳥類總體種間聯結8月份為顯著負聯結,其余各月為顯著正聯結(表2)。大山雀的生態位寬度值較大,屬廣性分布[26]森林鳥類,而且是調查中出現的頻率最高的,因而,本文以大山雀作為“目標種”,分析其與留鳥、夏候鳥、冬候鳥、旅鳥等不同居留型鳥類的聯結關系。
2.1 種間聯結檢驗與聯結度測度指數的選擇
聯結度的測定及其統計意義的檢驗常常是混亂的[1],每個指數都有其自己的性質,同時它們能得出很不同的結果[27],因而,選取物種數量較多、大山雀聯結種對較多的2012年4月26日全天調查數據作為日尺度的研究對象,通過對χ2統計量、點相關系數PCC、共同出現百分率PC、OI系數4個指數進行對比分析,篩選適合用于鳥類種間聯結分析的聯結度檢驗與測度指數。
由χ2檢驗可知,在18個物種的153個種對中,正負聯結聯結比為9∶8,其中6個種對呈現極顯著正聯結(P<0.01),大山雀與大嘴烏鴉(Corvus macrorhynchus)、黃腰柳鶯(Phylloscopus proregulus)、喜鵲(Pica pica);大嘴烏鴉與黃腰柳鶯、喜鵲;黃腰柳鶯與喜鵲。顯著正聯結(0.01<p<0.05)有8個種對:大山雀與紅嘴藍鵲(Urocissa erythrorhyncha)、沼澤山雀(Parus palustris);大嘴烏鴉與紅嘴藍鵲、沼澤山雀;黃腰柳鶯與紅嘴藍鵲、沼澤山雀;喜鵲與紅嘴藍鵲、沼澤山雀。大山雀、大嘴烏鴉、黃腰柳鶯、喜鵲4個物種分別與北紅尾鴝(Phoenicurus auroreus)、黃眉柳鶯(Phylloscopus inornatus)、白眉鹀(Emberiza tristrami)、大斑啄木鳥(Dendrocopos major)、環頸雉(Phasianus colchicus)、黃喉鹀(Emberiza elegans)、灰頭綠啄木鳥(Picus canus)、山斑鳩(Streptopelia orientalis)、山噪鹛(Garrulax davidi)、星頭啄木鳥(Dendrocopos canicapillus)、銀喉長尾山雀(Aegithalos caudatus)、棕頭鴉雀(Paradoxornis webbianus)12個物種,組成的48個種對呈顯著負聯結(0.01<P<0.05)(圖 1)。
點相關系數 PCC 指數值可劃分 0≤PCC<0.3,0.3≤PCC<0.5,0.5≤PCC<0.7,PCC≥0.7,-0.3≤PCC<0,

圖1 全天數據的χ2指數半矩陣Fig.1 Semi-matrix graph ofχ2 value for all-day data
Ochiai系數[25]:-0.5≤PCC<-0.3,-0.7≤PCC<-0.5,-1≤PCC<-0.7 等 8 個區間來表現種間聯結程度[28]。PCC 為正值時表示正聯結,數值越大聯結越強,PCC≥0.7有42個種對,包含χ2檢驗中呈極顯著正聯結的6個物種對;0.5≤PCC<0.7 有 11 個物種對,0.3≤PCC<0.5 有 10 個物種對,均為不顯著正聯結;0≤PCC<0.3 有 18 個種對,其中有8個種對在χ2檢驗中表現出顯著正聯結。PCC為負值時表示負聯結,指標絕對值越大負聯結性越強,-0.3≤PCC<0有58個物種對,其中48個物種對在χ2檢驗中呈顯著負聯結關系;-0.5≤PCC<-0.3有11個種對;-0.7≤PCC<-0.5有3個物種對;這兩個區間均為不顯著負聯結。可見,PCC指標在描述顯著性種對時絕對值偏小,χ2檢驗的顯著聯結種對多數在PCC指標絕對值0.3以下,但PCC能準確的反映物種間的正負聯結性,與χ2檢驗的結論基本一致(圖2)。
共同出現百分率 PC 指數值可劃分為 PC<0.3,0.3≤PC<0.5,0.5≤PC<0.7,PC≥0.7,4 個區間來表現種間聯結程度[5]。PC指數越大正聯結越強,PC≥0.7有48個種對,均為正聯結關系,其中有6對χ2檢驗為極顯著正聯結,4對為顯著正聯結;0.5≤PC<0.7有15個種對,均為正聯結種對,其中4個種對χ2檢驗為顯著正聯結;χ2檢驗為顯著負聯結的種對PC指標均在0.4以下(圖3)。
OI指數值可劃分為 OI<0.3,0.3≤OI<0.5,0.5≤OI<0.7,OI≥0.7,4 個區間來表現種間聯結程度[29]。OI≥0.7有63個物種對,均為正聯結,比同區間下的PC指標多了15個種對,其中6對為極顯著正聯結,有8個物種對呈顯著正聯結,0.5≤OI<0.7 有 27 個種對,其中 8 個物種對呈顯著負聯結,0.3≤OI<0.5 有 42 個種對,多數為負聯結。本研究中OI指標表現種間聯結時較PC指標偏大,不能很好的揭示聯結關系(圖4)。

圖2 全天數據的PCC指數半矩陣Fig.2 Semi-matrix graph of the point correlation coefficients(PCC)for all-day data

圖3 全天數據的PC指數半矩陣Fig.3 Semi-matrix graph of percentage co-occurrence(PC)for all-day data
綜上所述,PCC、OI、PC三種關聯度在反映種間聯結情況上,OI、PC實質上是等效的,其值越高物種對同時出現的機率越大,但OI指標在本研究中數值過大,不能準確的反映聯結關系,同時,PCC與PC的結果與整體常是無偏的[30-31],即使是在樣方數較少的情況下,也與總體一致。因此,選擇χ2檢驗、PCC、PC指數相結合能充分反映鳥類群落的種間聯結。這一結論與菌物群落的種間聯結研究是一致的[11]。

圖4 全天數據的OI指數半矩陣Fig.4 Semi-matrix graph of Ochiai index(OI)for all-day data
2.2 測度時間尺度確定
植物的種間聯結性跟樣方大小有較大關系,種間相互作用有一定的空間范圍,超過一定的界限就不會有相互作用[6]。由于鳥類是活動的,本研究調查采用林中視能及的以25 m為半徑的圓形樣方組成的樣帶進行全天候定位觀測[32],這里暫且不考慮空間問題,而是針對鳥類的活動性,探討在時間尺度上的種間聯結的差異。
2.2.1 日尺度數據分析
將4月份觀測的數據劃分為全天、上午、下午3個尺度,分別進行χ2檢驗、PCC、PC指數進行聯結分析,繪出星座圖(圖5),探討在時間維度上鳥類聯結性的變化情況。
χ2檢驗,全天、上、下午共出現18、17、12個種,全天數據有62個顯著聯結種對,上、下午的顯著聯結種對別為33、30個,與大山雀有顯著聯結的種對依次是17、16、11個,分別占各時間尺度總體數量的27%、48%、37%,可見與大山雀有關的種對在總體數量中占有很大比例(圖1;圖5a,b)。
PCC指數,在-0.3≤PCC<0區間內全天、上、下午分別有58、42、24個種對,全天種對數將近下午的2倍,與大山雀組成的種對依次分別是12、11、7對,占各時間尺度的21%、26%、29%。當PCC≥0.7時,在全天、上、下午的種對數分別為42、31、13個,全天種對數將近下午的3倍,與大山雀組成的種對依次分別是3、2、2對,占各時間尺度的7%、6%、15%(圖2;圖5c,d)。
PC指數,在區間PC≥0.7上全天、上、下午的種對數分別是47、36、13個,全天種對數將近下午的3倍,與大山雀有關的種對數依次分別是4、3、2對,占各時間尺度的9%、8%、11%。0≤PC<0.3區間內全天、上、下午的種對數分別是73、59、34個,全天種對數將近下午的2倍,與大山雀有關的種對依次是11、10、6個,占各時間尺度的 15%、17%、18%(圖 3;圖 5e,f)。

圖5 不同時間尺度的種間聯結的星座圖Fig.5 The constellation diagrams of temporal scales
2.2.2月尺度的數據分析
由于鳥類的居留、繁殖、遷移等季節動態性特征,僅以日尺度上分析種間聯結存有不確定性。因而,用月連續全天調查數據,計算大山雀與4種居留型的PCC月聯結頻率。
PCC月聯結頻率由正、負聯結月份數分別除以11(調查取樣為11個月,2月份數據由于春節原因沒有調查)來計算,這里假設留鳥全年12個月均棲息于研究地區。然而,對于夏候鳥、冬候鳥、旅鳥而言,其在研究地區的全年活動時間并不是12月,北京地區夏候鳥、冬候鳥駐留的時間一般分別在3—10月份和10—3月份,旅鳥春、秋季遷徙時停留在北京的時間分別在3—5月份和8—10月份。因此,為了保證PCC月聯結頻率可比性,對于夏候鳥,采用駐留期PCC月聯結頻率,由正、負聯結月份數分別除以除以8(北京地區夏候鳥最長駐留月份數)來計算;對于冬候鳥,采用駐留期PCC月聯結頻率,由正、負聯結月份數分別除以除以6(北京地區冬候鳥最長駐留月份數)來計算;對于旅鳥,采用停留期PCC月聯結頻率,由正、負聯結月份數分別除以除以6(北京地區旅鳥最長停留月份數)來計算。
2.3 大山雀與其他物種聯結性分析
χ2檢驗顯著性分析可知,大山雀與留鳥的顯著正關聯種對共有61個,占所有顯著種對的31%,聯結關系非常密切。進一步結合鳥類的營巢和取食習性集團[15-16]的劃分(附錄2)、生態位等來分析大山雀與四種居留型鳥類的聯結關系:
2.3.1 大山雀與留鳥物種聯結性分析
根據χ2檢驗、PCC、PC與PCC的月聯結頻率(表2),將大山雀與其他留鳥的聯結可劃分成3個聯結組5類,正聯結組:①大嘴烏鴉、喜鵲、灰喜鵲、紅嘴藍鵲;正負均等組:②沼澤山雀(Parus palustris);負聯結較強的組:③銀喉長尾山雀(Aegithalos caudatus)、褐頭山雀,④環頸雉、山噪鹛(Garrulax davidi)、大斑啄木鳥(Dendrocopos majo r),⑤灰頭綠啄木鳥(Picus canus)、小嘴烏鴉(Corvus corone)。
(1)大山雀與大嘴烏鴉、喜鵲、灰喜鵲、紅嘴藍鵲的PCC月聯結頻率正聯結較高,達到73%以上(表2),同時,極顯著以及顯著正聯結月份的共同出現比率PC、PCC多大于0.5,說明大山雀與這四種的正聯結強度很大。大嘴烏鴉、喜鵲、灰喜鵲、紅嘴藍鵲均屬于鴉科鳥類,表明大山雀與鴉科鳥類的聯結性較高,這四種鳥類屬于林冠樹枝營巢集團,大山雀是林冠取食集團[16],大山雀與以上物種在棲息空間上有重疊,同時大山雀、大嘴烏鴉、喜鵲、灰喜鵲、紅嘴藍鵲均為西山林場的優勢鳥類物種[32],共同遇見率高(附錄1)。
(2)大山雀與沼澤山雀的PCC正負聯結頻率比是1∶1,聯結頻率相同,χ2檢驗顯著正聯結為4、5、6月份(繁殖期),PC指標均大于0.5;顯著負聯結為3、7、10、11月份。兩物種的棲息生境與取食習性相近,生態位重疊度很大[33],雖然均為洞巢與林冠食蟲集團[34],但兩物種在取食高度,樹冠層取食位置(水平和垂直),取食樹種、取食食物大小和數量等方面存在差異,使競爭減少到最低程度[35],說明是繁殖行為導致了正聯結。
(3)大山雀與褐頭山雀、銀喉長尾山雀同科的3個種均為常見種,χ2檢驗顯示種對間整體負聯結強于正聯結。三者均為林冠食蟲集團,食性和取食行為相似(附錄1,附錄2),但其PCC月聯結頻率負聯結30%—50%之間,其正聯結都是9%,種間負聯結強于正聯結(表2),這緣于有相似習性的山雀類與大山雀在共同分布地區內少見[36]。
(4)大山雀與環頸雉、山噪鹛、大斑啄木鳥的PCC月聯結頻率負聯結均大于50%,正聯結小于30%。僅環頸雉在3月份與大山雀表現出顯著正聯結,而其負聯結月份均具有顯著性,同時,聯結指標顯示負聯結強度較大。環頸雉屬于灌草叢營巢、地面取食集團,與大山雀生態位重疊度較低[37];山噪鹛屬于灌草叢營巢、灌叢取食集團,大斑啄木鳥洞巢營巢、樹干取食集團[16],兩物種不僅在營巢和取食行為上與大山雀具有差異性,而且僅為林區落葉闊葉林的常見種[32],因而,導致環頸雉,大斑啄木鳥,山噪鹛與大山雀的遇見率低,從而導致它們負聯結。
(5)大山雀與小嘴烏鴉、灰頭綠啄木鳥的PCC月聯結頻率正負聯結頻率都低于30%,但負聯結強于正聯結,同時兩物種遇見率較低(附錄1)。這是由于大山雀常利用啄木鳥廢巢來筑巢[38-39];灰頭綠啄木鳥屬于洞巢營巢、樹干取食集團[40],小嘴烏鴉屬于林冠枝干營巢、地面雜食取食集團[16]。
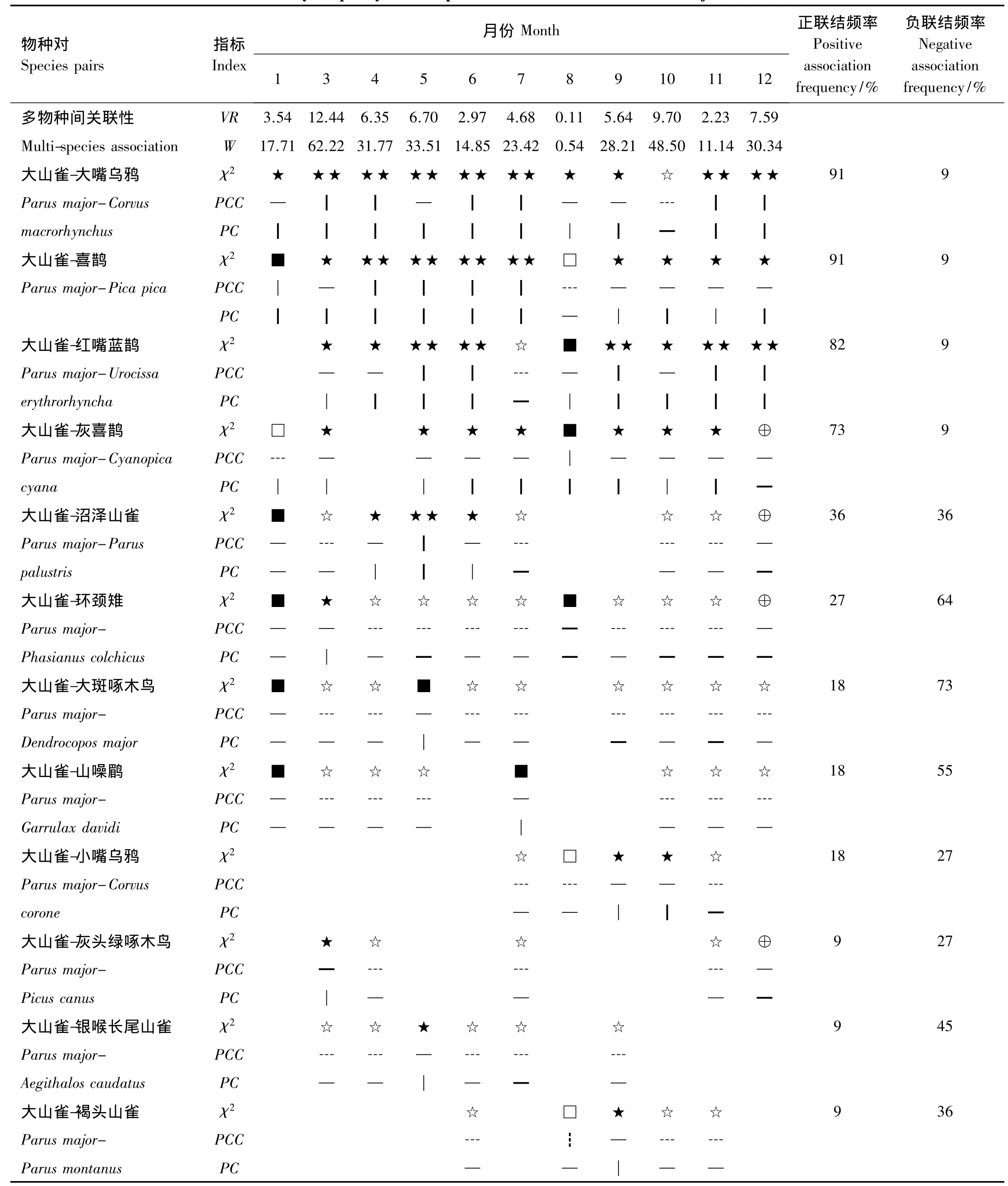
表2 大山雀與其他留鳥種間月聯結頻率Table 2 The monthly frequency of interspecific associations between Parus major and other residents
2.3.2 大山雀與夏侯鳥物種聯結性分析
根據χ2檢驗、PCC、PC與PCC的月聯結頻率(表3),將大山雀與夏候鳥的聯結劃分成3個聯結組,正聯結組:①四聲杜鵑(Cuculus micropterus);②正負均較弱的組為白眉姬鹟(Ficedula zanthopygia)、黃腹山雀(Parus venustulus);③負聯結較強的組為黃腰柳鶯、北紅尾鴝(Phoenicurus auroreus)。

表3 大山雀與夏侯鳥種間月聯結頻率Table 3 The monthly frequency of interspecific associations between Parus major and summer migrants
(1)大山雀與四聲杜鵑在5—7月份均呈顯著正聯結,共同出現百分率PC較高在0.5以上,駐留期PCC月聯結頻率正聯結38%,由于四聲杜鵑營巢屬于巢寄生性,灰喜鵲的巢穴是四聲杜鵑的常見寄居的巢穴[41],兩物種生態位重疊值較高[26],從而導致兩物種的正聯結。
(2)白眉姬鹟、黃腹山雀調查中僅5—7月份才有發現,導致PCC月聯結頻率正負聯結均小于30%,駐留期PCC月聯結頻率負聯結25%,但整體負聯結較強,這是由于白眉姬鹟屬于灌叢取食集團(附錄2),其在取食高度上白眉姬鹟在0—5 m的范圍,大山雀大于15 m的范圍[16];大山雀與黃腹山雀是同科物種,在筑巢和取食行為上均有相似性,但兩物種存在垂直代替現象[36],呈現負聯結。
(3)大山雀與黃腰柳鶯、北紅尾鴝PCC月聯結頻率負聯結在30%—50%之間,正聯結小于30%,負聯結強于正聯結,黃腰柳鶯1、4、5、7—11月份均有出現,1月份為負聯結,雖不具顯著性,但其PCC絕對值較高,說明大山雀與黃腰柳鶯的負聯結較強,其在1、11月份出現可能是由于氣候變暖,城市熱島效應使得一部分黃腰柳鶯居留型發生變化,對其的探討還有待研究,黃腰柳鶯也為林冠層取食集團(附錄2),駐留期PCC月聯結頻率負聯結達到63%,與大山雀存在垂直代替現象[36],表明兩物種競爭關系明顯。大山雀與北紅尾鴝多數月份呈負聯結,其駐留期PCC月聯結頻率負聯結較高,大山雀與北紅尾鴝食性有部分重疊(附錄1),雖然為林冠取食、洞巢集團(附錄2),但駐留期PCC月聯結頻率負聯結大于正聯結,是由于北紅尾鴝營巢喜于在人類的活動場所[42],西山地區靠近北京市區,為北紅尾鴝選擇理想巢址提供便利。
2.3.3 大山雀與冬侯鳥物種聯結性分析
根據χ2檢驗、PCC、PC與PCC的月聯結頻率(表4),大山雀與冬侯鳥劃分成2個聯結組:①正負聯結均較弱組為大山雀與棕眉山巖鷚(Prunella montanella);②負聯結組為斑鶇(Turdus naumanni)、田鹀(Emberiza rustica)、小鹀(Emberiza pusilla)、黃眉鹀(Emberiza chrysophrys)。
(1)大山雀與巖鷚科棕眉山巖鷚的正負聯結均較弱,駐留期PCC月聯結頻率正負聯結分別為17%、33%,PC為0.24—0.25;其在1月份與大山雀表現為不顯著正聯結,且PC指標較小;3、12月份為顯著負聯結,棕眉山巖鷚為地面屬于取食集團[43],與大山雀在習性上有較大差異。
(2)大山雀與鶇科的斑鶇、鹀科多種鳥類呈明顯的負聯結,駐留期PCC月聯結頻率負聯結為17%—50%,正聯結為0,PC為0.19—0.24;斑鶇屬地面取食集團(附錄2),鹀類屬于地面取食集團[43],且與大山雀在習性上有較大差異。

表4 大山雀與冬侯鳥種間月聯結頻率Table 4 The monthly frequency of interspecific associations between Parus major and winter migrants
2.3.4 大山雀與旅鳥物種聯結性分析
與大山雀有顯著正聯結關系的旅鳥2種,根據χ2檢驗、PCC、PC指數、停留期PCC月聯結頻率(表5),可劃分成2個聯結組,①正負聯結均較弱組為大山雀與褐柳鶯;②負聯結較強組為大山雀與黃眉柳鶯。

表5 大山雀與旅鳥種間月聯結頻率Table 5 The monthly frequency of interspecific associations between Parus major and migratory birds
(1)大山雀與褐柳鶯正負聯結均較弱,停留期PCC月聯結頻率正負聯結相同,5月、11月為顯著負聯結,8月為不顯著正聯結、10月為顯著正聯結。雖然同為林冠食蟲集團,由于其體重、嘴峰、跗跖都有明顯差異,使得在植物群落停息的水平位置和取食數量、大小上也有差別,說明對資源的再次分割導致了這種聯結關系。
(2)大山雀與黃眉柳鶯負聯結較強,4—5、8、10—11月PC指標均小于0.5,點相關系數PCC在8月份中為-0.61,停留期PCC月聯結頻率負聯結83.3%,雖然2種同屬林冠取食集團(附錄2),但二者存在垂直代替現象[36]。
3 結論
3.1 鳥類群落種間聯結度的測度
用固定樣帶法月連續全天定位調查,引入2×2列聯表以日尺度的定性數據檢驗與測度聯結度,與月尺度的聯結頻率結合起來,分析鳥類群落的種間聯結是可行的,因而,鳥類群落的種間聯結具有時間與空間尺度的雙重屬性。
3.2 以大山雀為“目標種”的月尺度種間聯結
大山雀與留鳥、夏候鳥、冬候鳥、旅鳥等不同居留型的聯結關系,隨著種數的減少,顯著聯結種對、聯結度、聯結復雜程度呈遞減趨勢;留鳥與夏候鳥之間通過PCC月聯結頻率正負均較弱的山雀科聯系著,夏候鳥與冬候鳥之間通過PCC月聯結頻率呈負聯結的鶇科而聯系著,夏候鳥與旅鳥之間通過PCC月聯結頻率呈負聯結的鶯科而聯系著。
3.2.1 大山雀與留鳥的種間聯結
大山雀與留鳥的聯結關系可劃分為正聯結組、正負聯結均等且較弱組、負聯結組。大山雀與生態位有部分重疊的鴉科鳥類呈明顯的正聯結,如大嘴烏鴉、灰喜鵲、喜鵲、紅嘴藍鵲等,PCC月聯結頻率正聯結為70%—90%,PC為0.25—0.98;大山雀與同科鳥類的由正負均等趨向于負聯結,如沼澤山雀、銀喉長尾山雀、褐頭山雀等,PCC月聯結頻率正聯結為9%—36%、負聯結為36%—45%,PC為0.2—0.98;大山雀與體型較大環頸雉、啄木鳥、山噪鹛、小嘴烏鴉等鳥類呈負聯結,PCC月聯結頻率正聯結為9%—27%、負聯結為27%—73%,PC 為 0.2—0.78。
3.2.2 大山雀與夏候鳥的種間聯結
大山雀與夏候鳥的聯結關系可劃分為正聯結組、正負聯結均較弱組、負聯結組。大山雀與杜鵑科四聲杜鵑呈正聯結,駐留期PCC月聯結頻率正聯結為38%,PC為 0.59—0.78;大山雀與同科的黃腹山雀、鹟科的白眉姬鹟等的正負聯結均較弱,駐留期PCC月聯結頻率正聯結均為13%,負聯結均為25%,PC為0.2—0.78;大山雀與鶯科的黃腰柳鶯、鶇科的北紅尾鴝等呈負聯結,駐留期PCC月聯結頻率正聯結為9—27%,負聯結頻率較高均為 63%,PC 為 0.2—0.98。
3.2.3 大山雀與冬候鳥的種間聯結
大山雀與巖鷚科棕眉山巖鷚的正負聯結均較弱,駐留期PCC月聯結頻率正負聯結分別為17%、33%,PC為0.24—0.25;大山雀與鶇科的斑鶇、鹀科多種鳥類呈明顯的負聯結,駐留期PCC月聯結頻率負聯結為17—50%,正聯結為 0,PC 為 0.19—0.24。
3.2.4 大山雀與旅鳥的種間聯結
大山雀與柳鶯科鳥類的月尺度聯結由正負聯結均較弱到明顯的負聯結,停留期PCC月聯結頻率,褐柳鶯正負聯結均為33%,黃眉柳鶯負聯結為83%;PC為0.2—0.59。
3.3 鳥類群落種間聯結度的生態機制
植物群落的種間聯結度的測度是空間結構與格局的表征,而鳥類群落的種間聯結度不僅是鳥類群落時空結構與格局的表征,而且反映了鳥類群落的動態性。鳥類群落的種間聯結對棲息的植物群落具有依賴性與關聯性,只有將植物、鳥類、昆蟲等組成的生物群落統一起來,才能更好地揭示其種間聯結的生態機制,這有待深入研究。
[1]Whittaker R H.Ordination of Plant Communities.The Hague:W.Junk,1978.
[2]Zhang J T.Quantitative Ecology.Beijing:Science Press,2004:107-111.
[3]Kershaw K A,Looney JH H.Quantitative and Dynamic Plant Ecology.London:Edward Arnold Limited.1985:78-94.
[4]Shen Y Y,Li Y,Lu N.Competitive ability and interspecific relationship among four forage species.Acta Pratacultural Science,2002,11(3):8-11.
[5]Wang B S,Peng S L.Studies on the measuring techniques of interspecific association of lower-subtropical evergreen-broadleaved forests-I.the exploration and the revision on the measuring formulas of interspecific association.Acta Phytoecologica et Geobotanica Sinica,1985,9(4):274-285.
[6]Guo Z H,Zhuo Z D,Chen J,Wu M F.Interspecific association of trees in mixed evergreen and deciduous broadleaved forest in Lushan mountain.Acta Phytoecologica Sinica,1997,21(5):424-432.
[7]Fang F,Hu Y K,Zhang W,Gong Y M,Liu Y Y,Yang X J.Numerical analysis of inter-specific relationships in Alpine steppe community in Bayanbulak.Acta Ecologica Sinica,2012,32(6):1898-1907.
[8]Lou Y J,Zhao K Y.Analysis of interspecific associations of Carex lasiocarpa community in recent 30-year succession in Sanjiang Plain.Chinese Journal of Ecology,2008,27(4):509-513.
[9]Kang B,Liu SR,Cai D X,Wen Y G,Shi Z M,Guo W F,Zhu H G,Zhang G J,Liu L.Species composition and correlation of understorey woody plants in chinese fir plantation in the lower subtropical area.Acta Ecologica Sinica,2005,25(9):2173-2179.
[10]Chu G M,Pan C D,Wan M,Jin Y,Meng M,He JC.An analysis on interspecific association of dominant desert species in outer space of Karamay agricultural comprehensive developmental region.Journal of Xinjiang Agricultural University,2004,27(4):1-6.
[11]Hubalek Z.Coefficients of association and similarity,based on binary(presence-absence)data:an evaluation.Biological Reviews,1982,57(4):669-689.
[12]Zhang GM,Yang D R,Xu L,Peng Y Q,Lu Y.Species associations of fig wasps associated with Ficus racemosa in Xishuangbanna.Chinese Journal of Ecology,2003,22(4):20-26.
[13]Cui P,Deng W H.Review on the status and development of bird community research.Chinese Journal of Zoology,2007,42(4):149-158.
[14]De Greef R M,Tilghman N G,Anderson SH.Foraging guild of North American birds.Environmental Management,1985,9(6):493-536.
[15]Bi Z L,Zhang Z W,Song J.Preliminary study on the relationship between the bird community and vegetation structure at Dongling mountain of Beijing.Journal of Beijing Nornal University,2003,39(5):663-668.
[16]Fan X S,Hu D F,Chen H Z,Wang F,Su X.Study on blocstructure and econiche characteristics of bird communities in the farming areas in North China plain.Arid Zone Research,2008,25(4):544-549.
[17]Gao Y,Qian G Z.The niche analysis of bird community in evergreen broad-leaf forest in Tiantong Mountain,Zhejiang Province.Acta Ecologica Sinica,1987,7(1):73-82.
[18]Schluter D.A variance test for detecting species associations,with some example applications.Ecology,1984,65(3):998-1005.
[19]Yates F.Contingency tables involving small numbers and the χ2test.Supplement to the Journal of the Royal Statistical Society,1934,1(2):217-235.
[20]Pielou E C.Association tests versus homogeneity tests:their use in subdividing quadrats into groups.Vegetatio,1969,18(1/6):4-18.
[21]Pielou E C.An Introduction to Mathematical Ecology.New York:Wiley Interscience,1969:286-286.
[22]Yule G U.On the methods of measuring association between two attributes.Journal of the Royal Statistical Society,1912,75(6):579-642.
[23]Whittaker R H,Fairbanks C W.A study of plankton copepod communities in the Columbia Basin,southeastern Washington..Ecology,1958,39(1):46-65.
[24]Walter H,Ellenberg H.Einführung in die Phytologie.Eugen Ulmer,1956,4:136-136.
[25]Driver H E,Kroeber A L.Quantitative Expression of Cultural Relationships.California:The University of California Publications in American Archaeology and Ethnology,1932,31:211-256.
[26]Chen Y.Ecological Distribution,the Relationship of Plants and the Feeding Habits of Birds in An Evergreen Broadleaved Forest in Tiantong's 20hm2plot,Zhejiang[D].Shanghai:East China Normal University,2012.
[27]Morisita M.Measuring of interspecific association and similarity between communities.Memoirs of the Faculty of Science,Kyushu University,Series E(Biology),1959,3:65-80.
[28]Shangguan T L,Zhang F.Research on the pattern and associations between dominants of the vegetation in mian mountain,Shanxi province.Journal of Wuhan Botanical Research,1988,6(4):354-364.
[29]Guo X Y,Li Y H,Lin H P.Studies on the Inter-specific Association among main species of cyclocarya paliurus community in Niumulin natural reserve.Journal of Fujian College of Forestry,2011,21(2):181-185.
[30]Goodall D W.Sample similarity and species correlation//Wittaker P H,ed.Ordination and Classification of Communities.The Hagues:W.Junk,1972:105-154.
[31]Liu P P,Cheng J M.Study on plant interspecific association.Research of Soil and Water Conservation,2000,7(2):179-184.
[32]Fan Z J,Dong D Y,Zheng R,Wang M Z,Wang Q F,Guan W B.Avian community diversity in Platycladus orientalis ancient trees at the Jingfu Temple in Beiing.Journal of Beijing Forestry University,2013,35(5):46-55.
[33]Chen H P,Gao Z X,Li X M.Structure of forest bird communities of Zhanhe forest region in summer.Jourual of Northeast Forestry University,1992,20(6):49-55.
[34]Hou JH,Wu M L,Hu Y F,Zhang X Z,Hu Z T.A school study on the vertical space patterns of bird community in Mulan wei chang forest park.Journal of Agricultural University of Hebei,2000,23(1):78-81.
[35]Gao W,Xiang G Q,Feng H L,Deng Q X,Zhao H.On changes of foraging behavior of the tits of two species in secondary deciduous wood.Chinese Journal of Ecology,1996,15(5):1-5.
[36]Zheng G M.Ecological distribution of birds in Beijing and its vicinity during summer.Zoological Research,1984,5(1):29-40.
[37]Hou JH,Guo SB,Li J L.Structure of bird communities of Xitai,Xiaowutai mountains in summer.Journal of Hebei Forestry College,1994,9(4):332-338.
[38]Cai Q K.The Avifauna of Beijing.Beijing:Beijing Press,1988.
[39]Zhou T.Structure Processes and Determinants of Bird Community in the Secondary Broadleaf Forest[D].Changchun:Northeast Normal University,2011.
[40]Gao W,Xiang G Q,Shang JW,Feng H L.On the guild structure and its relationship of secondary forest bird community.Journal of Northeast Forestry University,1990,18(S3):114-125.
[41]Zheng Z X,Xian Y H,Guan G X.Fauna Sinica Aves,Vol.6.Beijing:Science Press,1991.
[42]Zhao Z J.Ecology of Daurian redstart in Changbai area and the importance of plant protection.Acta Zoologica Sinica,1975,21(3):272-287.
[43]Zheng G M.Ecological distribution of birds during the winter in peking & vicinity.Acta Zoologica Sinica,1962,14(3):321-336.
參考文獻:
[2]張金屯.數量生態學.北京:科學出版社,2004:107-111.
[4]沈禹穎,李昀,陸妮.4種牧草種間競爭力和種間關系的研究.草業學報,2002,11(3):8-13.
[5]王伯蓀,彭少麟.南亞熱帶常綠闊葉林種間聯結測定技術研究——Ⅰ.種間聯結測式的探討與修正.植物生態學與地植物學叢刊,1985,9(4):274-285.
[6]郭志華,卓正大,陳潔,吳梅鳳.廬山常綠闊葉,落葉闊葉、混交林喬木種群種間聯結性研究.植物生態學報,1997,21(5):424-432.
[7]房飛,胡玉昆,張偉,公延明,柳妍妍,楊秀娟.高寒草原植物群落種間關系的數量分析.生態學報,2012,32(6):1898-1907.
[8]婁彥景,趙魁義.三江平原毛苔草群落近30年演替過程中的種間聯結性分析.生態學雜志,2008,27(4):509-513.
[9]康冰,劉世榮,蔡道雄,溫遠光,史作民,郭文福,朱宏光,張廣軍,劉磊.南亞熱帶人工杉木林灌木層物種組成及主要木本種間聯結性.生態學報,2005,25(9):2173-2179.
[10]楚光明,潘存德,萬猛,晉瑜,蒙敏,何江成.克拉瑪依農業綜合開發區外圍主要荒漠植物種間聯結測定分析.新疆農業大學學報,2004,27(4):1-6.
[12]張光明,楊大榮,徐磊,彭艷瓊,盧耀.西雙版納聚果榕榕果小蜂種間聯結性研究.生態學雜志,2003,22(4):20-26.
[13] 崔鵬,鄧文洪.鳥類群落研究進展.動物學雜志,2007,42(4):149-158.
[15]畢中霖,張正旺,宋杰.北京東靈山地區鳥類群落與棲息環境的關系研究.北京師范大學學報(自然科學版),2003,39(5):663-668.
[16]范喜順,胡德夫,陳合志,王峰,蘇鑫.華北平原耕作區鳥類群落的集團結構及生態位.干旱區研究,2008,25(4):544-549.
[17]高穎,錢國楨.天童常綠闊葉林中鳥類群落結構的空間生態位分析.生態學報,1987,7(1):73-82.
[26]陳宇.天童常綠闊葉林樣地鳥類生態分布及鳥類與植物關系[D].上海:華東師范大學,2012.
[28]上官鐵梁,張峰.山西綿山植被優勢種群的分布格局與種間聯結的研究.武漢植物學研究,1988,6(4):357-364.
[29]郭相億,李裕紅,林慧萍.牛姆林區青錢柳群落的主要種群種間關聯特征.福建林學院學報,2001,21(2):181-185.
[31]劉萍萍,程積民.植物種間聯結關系的研究.水土保持研究,2000,7(2):179-184.
[32]范宗驥,董大穎,鄭然,王敏增,王奇峰,關文彬.北京靜福寺側柏古樹林鳥類群落多樣性研究.北京林業大學學報,2013,35(5):46-55.
[33]陳化鵬,高中信,李先敏.沾河林區夏季森林鳥類群落結構研究.東北林業大學學報,1992,20(6):49-55.
[34]侯建華,武明錄,胡永富,張向忠,胡振天.木蘭圍場國家森林公園鳥類群落垂直空間格局研究.河北農業大學學報,2000,23(1):78-81.
[35]高瑋,相桂權,馮賀林,鄧秋香,趙虹.次生闊葉林中大山雀和沼澤山雀取食行為的研究.生態學雜志,1996,15(5):1-5.
[35]鄭光美.北京及其附近地區夏季鳥類的生態分布.動物學研究,1984,5(1):29-40.
[36]鄭光美.北京及其附近地區夏季鳥類的生態分布.動物學研究,1984,5(1):29-40.
[37]侯建華,郭書彬.小五臺山西臺夏季鳥類群落結構研究.河北林學院學報,1994,9(4):332-338.
[38]蔡其侃.北京鳥類志.北京:北京出版社,1988
[39]周彤.山地次生闊葉林鳥類群落的確定和結構過程[D].長春:東北師范大學,2011.
[40]高瑋,相桂權,尚金城,馮賀林.山地次生林鳥類集團結構及關系的研究.東北林業大學學報,1990,18(S3):114-125.
[41]鄭作新,冼耀華,關貫勛.中國動物志·鳥綱:第六卷,鴿形目·鸚形目·鵑形目·鸮形目.北京:科學出版社,1991.
[42]趙正階.北紅尾鴝在長白山地區的生態學及其在植物保護中的意義之研究.動物學報,1975,21(3):272-287.
[43]鄭光美.北京及其附近地區冬季鳥類的生態分布.動物學報,1962,14(3):321-336.

附錄1 北京靜福寺鳥類調查名錄Appendix1 Bird survey list at Jingfu Temple,Beijing

續表

續表

附錄2 鳥類營巢及取食集團劃分Appendix 2 Divisions of the avian nesting and foraging guilds

續表

續表
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
