低溫對蝶蛹金小蜂卵成熟及其數量動態的影響
2013-12-19 08:41:00夏詩洋李保平
生態學報 2013年4期
夏詩洋,孟 玲,李保平
(南京農業大學植物保護學院,農作物生物災害綜合治理教育部重點實驗室,南京 210095)
寄生蜂產卵于寄主體內或體外,子代幼蟲依賴于寄主營養完成發育并最終致使寄主死亡。雌蜂終生實現的產卵量主要取決于3個因素的互作:遇到的適宜寄主數量、終生成熟的卵數量和產卵速率的行為調節[1]。由于環境因素的不確定性,每當遇到適宜的寄主時總有足夠的成熟卵可產的完全吻合情況是難以實現的。但有證據表明,自然選擇會調整卵發育特征以吻合遇到適宜寄主的預期[2-3]。
寄生蜂中不同類群間的卵成熟特征存在很大差異。由于卵成熟特征與寄生蜂的生殖策略密切相關,因此,探究卵成熟動態就成為理解寄生蜂產卵策略以及生活史策略的重要途徑[1,4-8]。1950年Flanders首先提出了卵預熟和卵漸熟等兩類卵成熟模式,前者指成蟲羽化時所有卵均已成熟,后者指成蟲羽化后卵陸續成熟。這一劃分也成為后來建立寄生蜂生活史策略模型[7,9]和寄主-寄生蜂種群動態模型的基本屬性[10]。但這一分類方法的依據是定性特征,難以涵蓋復雜多樣的卵成熟方式。對此,2001年Jervis等提出定量指標—卵熟指數(成蟲羽化時成熟卵占一生總產卵量的比例)[1],用于認識寄生蜂的卵成熟規律,并發現該特征與其他生活史策略存在必然的聯系。后來該指標也被用于認識其他植食性昆蟲的卵成熟規律[8]。
對于卵漸熟類寄生蜂而言,根據進化原理預測,自然選擇理應偏好那些能夠協調好卵成熟進程以滿足遭遇適宜寄主時的產卵之需的寄生蜂,從而演化出具有最優抱卵量的雌蜂[11]。但根據生態學原理預測,自身生理因素和外界生態因素的復雜多變一定會影響寄生蜂的能量分配(生存抑或生殖),從而演化出動態的產卵決策行為。如何綜合考慮進化和生態學原理解釋寄生蜂的產卵策略,就成為寄生蜂進化生態學研究的主要挑戰之一。最優搜尋理論綜合運用進化和生態學原理來解釋生物對資源的獲取規律[12],不但是迄今最常用的解釋寄生蜂產卵策略(即最優產卵策略)的原理,而且成為寄生蜂行為生態學研究的基本原理[13],并用于指導生物防治實踐[14]。但用經典的最優搜尋原理解釋寄生蜂的產卵策略時,通常不考慮生理或其他生態因素的影響,實際上生理和生態因子可能通過影響雌蜂的卵成熟動態而影響雌蜂的產卵決策。
本研究以蝶蛹金小蜂(Pteromalus puparum)-菜粉蝶(Pieris rapae)為模式生物,該寄生蜂營群集性寄生方式,雌蜂在1頭寄主蛹(蝶類)內產若干卵,并發育成熟若干子代蜂[15-16]。其卵成熟方式屬于漸熟類,在遭遇缺乏適宜寄主時可臨時吸回卵營養[17]。蝶蛹金小蜂以老熟幼蟲或蛹在寄主菜粉蝶蛹內越冬,其生長和發育經歷冬季低溫[15-16],但迄今不清楚冬季低溫是否影響早春羽化雌蜂的成熟卵數量動態。由于蝶蛹金小蜂成蟲在寄主體內羽化后再爬出寄主,而且雄蜂先于雌蜂爬出,故需要掌握雌蜂的準確羽化時間才可及時解剖寄主,以獲得剛羽化的雌蜂進行解剖計數成熟卵數量,因此本研究首先詳細觀察了子代蜂幼期形態變化及其寄主蛹的形態;為探究冬季低溫對越冬雌蜂來年早春羽化后卵巢發育和成熟動態的影響,本研究模擬冬季低溫(5℃)處理了含有寄生蜂老熟幼蟲的寄主蛹,于雌蜂羽化后解剖觀察卵巢形態,并連續觀察了成熟卵數量的時間動態變化。該研究可為深入闡釋群集性寄生蜂的產卵策略提供生理依據。
1 材料與方法
1.1 材料
蝶蛹金小蜂和菜粉蝶均采自南京市東郊甘藍菜地。將采回的被寄生蛹放入智能人工氣候箱((25±1)℃,RH(60±10)%,L∶D=16∶8)待成蟲羽化;為成蟲提供新鮮蝶蛹供其寄生并繁育多代后供試。采回的菜粉蝶幼蟲用“京豐1號”甘藍葉片飼喂直至化蛹,取化蛹后24—48 h的蝶蛹供試。
1.2 幼期發育的形態觀察
給1頭雌蜂提供1頭寄主蛹寄生,然后每隔24 h解剖被寄生蛹,并在體視顯微鏡(Olympus SZ-CTV;麥克奧迪SMZ 171)下測量和拍照(寄主和子代蜂),測量蜂卵和幼蟲的體長和寬(最寬處),測量蛹體長;自第9天開始,解剖并取出蜂蛹保留,待羽化后確定性別。測量觀察個體數為20—30頭。
1.3 低溫處理試驗
設2個溫度處理:(25±1)℃(適宜溫度,對照)和(5±1)℃(低溫),該低溫為越冬期間持續時間最長的溫度。在低溫處理中,取寄生后發育到第9天(蜂處于老熟幼蟲或蛹階段)的寄主蛹放入人工氣候箱中(L∶D=16∶8)中低溫處理30 d,然后取出放入適宜溫度下,于4 d后逐日8:00檢查出蜂情況;從開始出蜂起連續7 d解剖雌蜂,觀察卵巢管、計數成熟卵量,并測量雌蜂后足脛節長度。對照則一直在適宜溫度下,于第13天開始逐日檢查出蜂情況,出蜂后按照上述方法解剖并測量雌蜂。觀察個體數量大于30頭。
1.4 數據分析
為揭示寄生蜂形態特征隨時間的變化規律,采用非參數平滑方法擬合變化趨勢。用基于quasipoissson分布型的對數線性模型擬合溫度處理對卵成熟數量的影響,把雌蜂體型大小(后足脛節長度)和日齡作為協變量放入模型。分析用R統計分析軟件[18-19]。
2 結果與分析
2.1 蝶蛹金小蜂胚后發育的形態及其寄主形態變化
蝶蛹金小蜂雌蜂一般喜好產卵于寄主菜粉蝶蛹的翅外側(圖1A)與內側(圖1B)之間,侵染寄主的一般順序為翅外側—翅內側—空腔—頭部與尾部(圖1)。子代蜂各發育期的體色及其寄主的體色存在差異(圖2,表1)。經歷2—3 d的暴食期后,幼蟲體長在孵化后第4天達到最大值,但體寬直到第7天達到最大;蛹體長變化不大,雌性明顯長于雄性(圖3)。

圖1 健康菜粉蝶蛹外部形態Fig.1 Exomorphology of healthy P.rapae pupae
2.2 低溫對蝶蛹金小蜂成熟卵數量動態的影響
適宜溫度條件下,羽化當天卵室形成并開始有卵黃沉積(圖4:0a);1天后在卵巢管底部出現成熟卵(圖4:1a);第2—3天,成熟卵大量形成,并逐漸往卵巢管端部堆積,接近底部的卵巢管略有增粗,頂部出現大量未成熟卵(圖4:2a—3a);第4天后在卵巢管底部出現明顯的卵吸收現象,卵巢管進一步增粗,卵量接近飽和(圖4:4a);第5—6天,成熟卵數量增加緩慢,甚至略有下降,并與卵吸收速度達到動態平衡,卵量飽和,卵巢管最粗(圖4:5a—6a)。而在低溫處理下,卵巢管較常溫下略細,成熟卵量較少(圖4:0b—6b)。

圖2 寄主菜粉蝶蛹外部形態隨蝶蛹金小蜂發育的日變化Fig.2 Daily exomorphology of parasitized P.rapae pupae along with P.puparum progeny

圖3 蝶蛹金小蜂幼蟲和蛹形態測量值的日變化Fig.3 Variation in morphological measures of P.puparum larva and pupa with time after parasitism
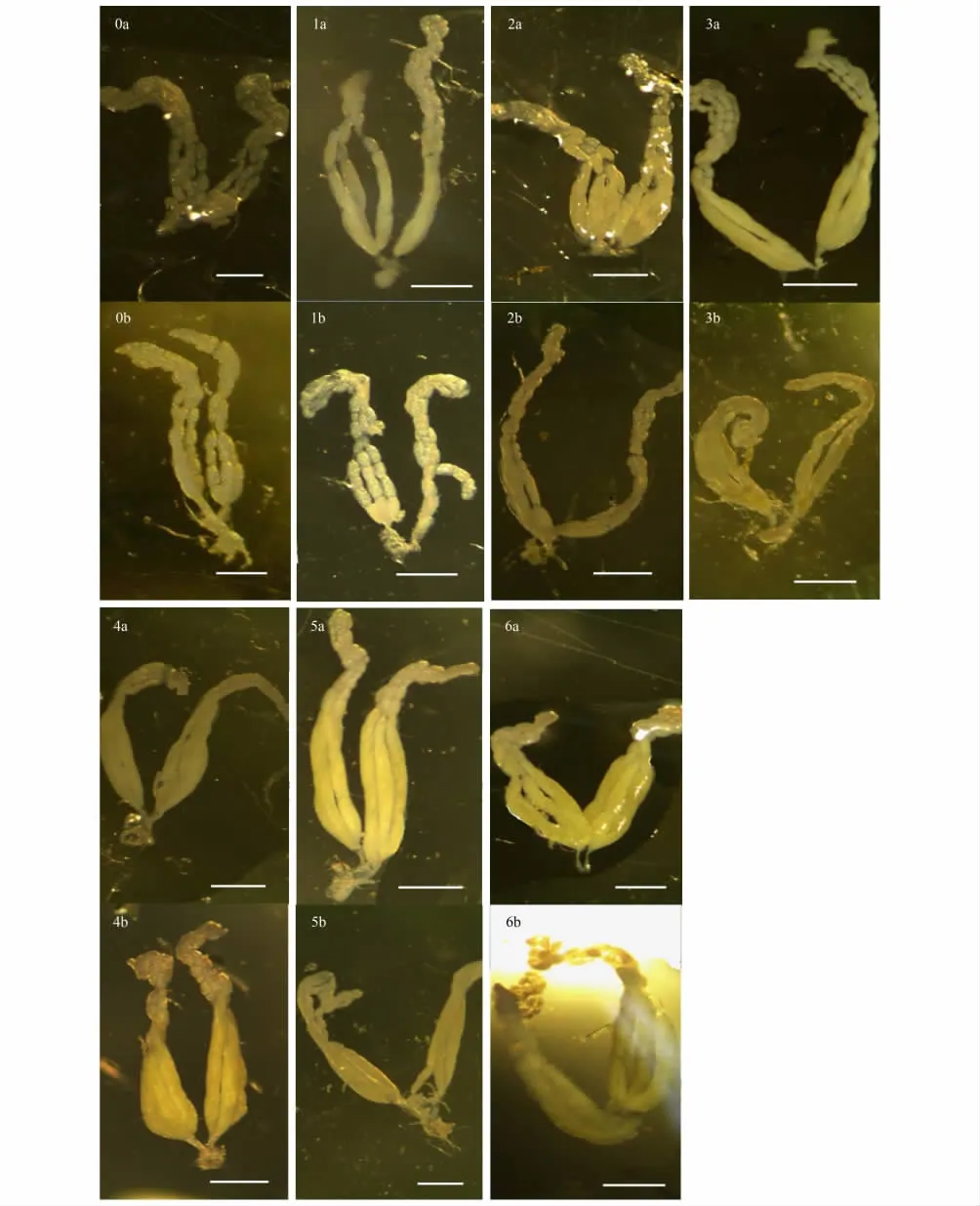
圖4 適溫與低溫處理下蝶蛹金小蜂成蟲卵巢管發育的變化Fig.4 Morphology of P.puparum ovary during adult stage under optimal and low temperature treatments
對數線性回歸模型分析表明,溫度和母蜂日齡不僅單獨、而且互作影響成熟卵的數量;母蜂體型大小顯著影響成熟卵的數量(表2)。與適溫處理相比,低溫處理明顯減緩卵成熟速率,各日齡期成熟卵數量均明顯減少;適溫與低溫處理下成熟卵數量的變化格局存在差異,適溫下前2d成熟卵量直線增加,此后2d放緩,然后(第4天起)不再增加;低溫下成熟卵量以較緩慢的速度直線增加至第4天,然后略放緩(圖5)。成熟卵量在不同溫度處理下均隨雌蜂體型增大而顯著增加,但在適溫下增加的幅度略小于低溫處理下(圖6)。

表1 蝶蛹金小蜂幼期發育各階段及其寄主菜粉蝶蛹的外部形態變化Table 1 Exomorphological changes of larval stages of P.puparum and its host P.rapae pupae
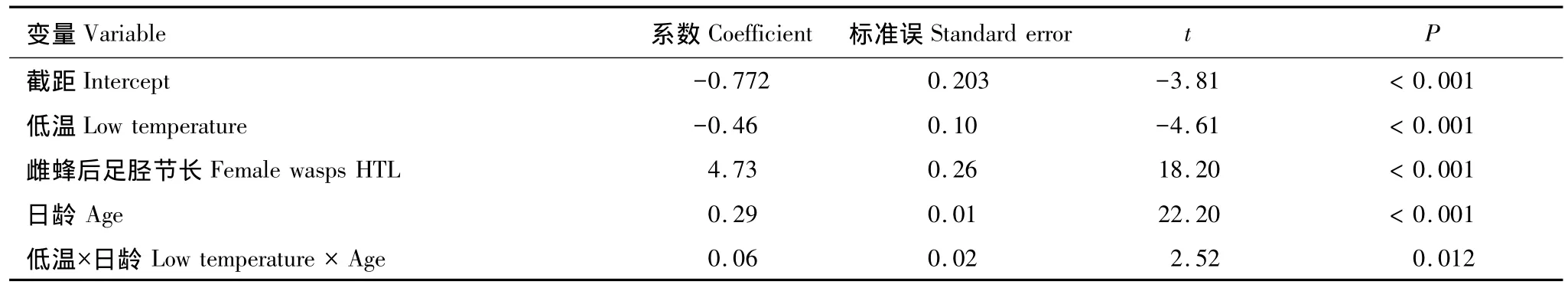
表2 溫度處理和其他協變量影響蝶蛹金小蜂成熟卵數量的對數線性模型參數估計值Table 2 Coefficient estimates of log-linear model for effects of temperature and other covariates on number of mature eggs in P.puparium

圖5 蝶蛹金小蜂成熟卵數量隨成蟲羽化后天數的變化Fig.5 Variation in number of mature eggs with time after adult emergence in P.puparum under optimal and low temperature treatments
3 討論
從寄主外部形態變化推測其體內寄生蜂幼期發育進程,不僅對于野外調查寄生蜂種群數量動態具有參考價值,而且可為及時解剖在寄主體內羽化的雌蜂、檢查卵成熟動態提供依據。龍心林和陳惠英[20]曾對蝶蛹金小蜂蛹的體色變化進行過簡單的文字描述,但在參考中由于沒有圖片而難以把握。對此,本研究對幼期發育的各階段及其寄主菜粉蝶蛹的形態進行了更為詳細的文字描述和拍照;逐日解剖觀察發現,蝶蛹金小蜂從卵孵化開始歷時約2周化蛹,在寄生后第2—4天可將寄主取食殆盡。這一特性的適應意義可能在于,被寄生的寄主在早期易于受到病原微生物的侵染,加速取食可以降低寄主染病的機會。這一特性與蒼白寬口姬小蜂(Hyssopus pallidus)相似[21]。本研究對被寄生的寄主蛹外部形態(主要是體色)隨體內蝶蛹金小蜂發育而發生的變化進行了描述,該結果在田間調查菜粉蝶與蝶蛹金小蜂種群動態中為推測寄生進程提供參考依據。

圖6 不同溫度處理蝶蛹金小蜂成熟卵量隨體型大小的變化Fig.6 Variation in number of mature eggs with body size under optimal and low temperature treatments
許多研究表明,生殖器官易于受到低溫脅迫的不良[22]。本研究發現,對被寄生的寄主蛹(蝶蛹金小蜂成蟲羽化前)進行低溫處理(模擬越冬溫度)后,卵巢管萎縮,卵成熟速度減緩,抱卵量降低。同樣的現象在其他寄生蜂中也有報道,例如,對卡氏小蜂(Euchalcidia caryobori(Hanna))的研究表明,低溫能延遲卵成熟,甚至引起雌、雄蜂生殖器官畸形[23];低溫處理達到一定強度和持續時間后,還可能降低寄生蜂的生殖力[24-26]。但也有報道未發現低溫對寄生蜂生殖力造成不良影響[27],可能是低溫處理尚未達到脅迫程度的原因。
卵的成熟和再吸收在卵漸成熟型寄生蜂中維持著動態平衡,是寄生蜂協調產卵策略的重要表現[13]。本研究表明,成熟卵數量的時間變化格局不僅受低溫處理、而且受雌蜂體型大小的影響,低溫處理明顯減緩卵成熟速率,推遲達到最大成熟卵數量的時間和卵吸收時間。該結果說明,越冬會明顯延緩蝶蛹金小蜂的卵成熟動態,進而可能影響春季羽化雌蜂的產卵決策行為。
致謝:感謝南京農業大學植物保護學院劉潔在試驗中給予的幫助。
[1] Jervis M A,Heimpel G E,Ferns P N,Harvey J A,Kidd N A C.Life-history strategies in parasitoid wasps:a comparative analysis of‘ovigeny’.Journal of Animal Ecology,2001,70(3):442-458.
[2] Rosenheim J A.An evolutionary argument for egg limitation.Evolution,1996,50(5):2089-2094.
[3] Heimpel G E,Rosenheim J A.Egg limitation in parasitoids:a review of the evidence and a case study.Biological Control,1998,11(2):160-168.
[4] Flanders S E.Regulation of ovulation and egg disposal in the parasitic hymenoptera.The Canadian Entomologist,1950,82(6):134-140.
[5] Price P W.Niche breadth and dominance of parasitic insects sharing the same host species.Ecology,1971,52(4):587-596.
[6] Jervis M A,Kidd N A C.Host-feeding strategies in hymenopteran parasitoids.Biological Reviews,1986,61(4):395-434.
[7] Ellers J,Sevenster J G,Driessen G.Egg load evolution in parasitoids.American Naturalist,2000,156(6):650-665.
[8] Jervis M A,Ferns P N.The timing of egg maturation in insects:ovigeny index and initial egg load as measures of fitness and of resource allocation.Oikos,2004,107(3):449-461.
[9] Rosenheim J A,Heimpel G E,Mangel M.Egg maturation,egg resorption,and the costliness of transient egg limitation in insects.Proceedings of the Royal Society B:Biological Sciences,2000,267(1452):1565-1573.
[10] Shea K,Nisbet R M,Murdoch W W & Yoo H J S.The effect of egg limitation on stability in insect host-parasitoid population models.Journal of Animal Ecology,1996,65(6):743-755.
[11]Jervis M A,Vilhelmsen L B.Mouthpart evolution in adults of the basal,‘symphytan’,hymenopteran lineages.Biological Journal of Linnean Society,2000,70(1):121-146.
[12] Stephens D W,Krebs J R.Foraging Theory.Princeton:Princeton University Press,1986.
[13] Godfray H C J.Parasitoids:Behavioural and Evolutionary Ecology.Princeton:Princeton University Press,1994:99-125.
[14] Mills N J,Wajnberg é.Optimal foraging behavior and efficient biological control methods//Wajnberg é,Bernstein C,van Alphen J,eds.Behavioral Ecology of Insect Parasitoids:from Theoretical Approaches to Field Applications.Oxford:Blackwell,2008:3-30.
[15] Hu C.Life history and occurrence of Pteromalus puparum L.in Hangzhou.Acta Entomologica Sinica,1984,27(3):302-307.
[16] Ho C L.A study on the hymneopterous parasites of cabbage butterfly(Pieris rapae L.)in Nanking.Acta Entomologica Sinica,1962,11(1):45-58.
[17] Guo J Y,Dong S Z,Ye G Y,Li K,Zhu J Y,Fang Q,Hu C.O?sorption in the endoparasitoid,Pteromalus puparum.Journal of Insect Science,2011,11:90.
[18] R Development Core Team.R:A language and environment for statistical computing.R Foundation for Statistical Computing,Vienna,Austria,2007.ISBN 3-900051-07-0,URL.[2009-5-31].http://www.R-project.org.
[19] Crawley M J.The R Book.London:John Wiley& Sons Ltd,2007.
[20] Long X L,Chen H Y.Summer and autumn generations and pupal colour changes of Pteromalus puparum.Natural Enemies of Insects,1988,10(2):122-122.
[21] Tschudi-Rein K,Dorn S.Reproduction and immature development of Hyssopus pallidus(Hymenoptera:Eulophidae),an ectoparasitoid of the codling moth.European Journal of Entomology,2001,98:41-45.
[22] Denlinger D L,Lee R E Jr.Physiology of cold sensitivity//Hallman G J,Denlinger D L,eds.Temperature Sensitivity in Insects and Application in Integrated Pest Management.Boulder:Westview Press,1998:55-96.
[23] Hanna A D.Fertility and toleration of low temperature in Euchalcidia carybori Hanna(Hymenoptera:Chalcidinae).Bulletin of Entomological Research,1935,26(3):315-322.
[24] Foerster L A,Nakama P A.Efeito da estocagem em baixa temperatura na capacidade reprodutiva e longevidade de Trissolcus basalis(Wollaston)e Telenomus podisi Ashmead(Hymenoptera:Scelionidae).Neotropical Entomology,2002,31(1):115-120.
[25] ?zder N.Effect of different cold storage periods on parasitization performance of Trichogramma cacoeciae(Hymenoptera,Trichogrammatidae)on eggs of Ephestia kuehniella(Lepidoptera,Pyralidae).Biocontrol Science and Technology,2004,14(5):441-447.
[26] Al-Tememi N K,Ashfaq M.Effect of low temperature storage on the fecundity and parasitizing efficacy of Bracon hebetor(Say).Journal of Agricultural Research,2005,43:155-160.
[27] Ismail M,Vernon P,Hance T,van Baaren J.Physiological costs of cold exposure on the parasitoid Aphidius ervi,without selection pressure and under constant or fluctuating temperatures.BioControl,2010,55(6):729-740.
參考文獻:
[15] 胡翠.蝶蛹金小蜂在杭州的年生活史.昆蟲學報,1984,27(3):302-307.
[16] 何繼龍.南京地區菜粉蝶Pieris rapae Linné寄生蜂考查.昆蟲學報,1962,11(1):45-57.
[20] 龍心林,陳惠英.蝶蛹金小蜂夏秋世代及蛹色變化.昆蟲天敵,1988,10(2):122-122.
