三亞巖相潮間帶底棲海藻群落結構及其季節變化
2013-12-16 08:19:10陳自強廖一波高愛根曾江寧陳全震
生態學報 2013年11期
關鍵詞:物種
陳自強,壽 鹿,廖一波,高愛根,曾江寧,陳全震
(國家海洋局海洋生態系統與生物地球化學重點實驗室,國家海洋局第二海洋研究所,杭州 310012)
潮間帶是海洋生態系統的重要棲息地類型,位于海陸過渡地帶,能支撐很高的海洋生物多樣性[1]。大型底棲海藻作為潮間帶生態系統的重要組成部分,是巖相潮間帶生態系統重要的物質基礎,其種類組成直接影響著底棲生物群落的初級生產力,在近岸碳循環中起著重要作用[2-4]。國內外對于潮間帶底棲海藻的生態學研究早期主要集中于藻類群落的區系劃分、種類組成、分布特征等基礎研究方面[5-6]。近年來,潮間帶底棲藻類的研究向藻類生理生態、共生機制等更加微觀的方向發展,并開始關注藻類群落內部的營養結構、與其它共生群落的相互影響以及在整個潮間帶食物網中的作用[7-9]。
然而,由于調查取樣的困難以及分類學的局限,我國南海區的基礎資料陳舊,系統深入的研究僅見于中國科學院海洋所和南海所于20世紀60年代初和80年代所做的工作[10-11]。隨著近20年來海岸開發、旅游業的發展以及當地人口的快速增長,三亞潮間帶生態環境受到了人類活動的深刻影響[12-14],先前的資料已不能反映目前底棲海藻群落的實際情況。另外,水溫和光照等季節相關的因素是影響大型海藻分布的主要影響因子之一[2],三亞地處熱帶,常年水溫高、光照強,四季差異不大,所以相比溫帶海區,熱帶海區海藻群落的季節變化經常被忽略[15]。因此,本文通過對三亞巖相潮間帶大型底棲海藻的調查,試圖查明:(1)近年來三亞潮間帶大型海藻的分布特征;(2)海藻群落是否存在明顯的季節變化;(3)此區域大型海藻與我國其它海區在溫度性質上的差異。旨在為我國南海南區大型藻類的多樣性研究提供基礎資料,為三亞潮間帶藻類資源的保護以及合理利用提供依據。
1 材料與方法
1.1 研究區域概況
三亞地區地處大陸型島嶼海南島的最南端,位于北緯 18°09′34″—18°37′27″,東經 108°56′30″—109°48′28″,鄰近海域的地理氣候和環境條件具有典型的熱帶海洋特征。潮汐為非正規全日潮,年均潮差小于1 m[16],潮間帶以巖礁、珊瑚礁、石沼等巖相底質類型為主,底棲海藻資源豐富。
1.2 采樣方法
根據底質類型,在三亞沿海設置6條巖相潮間帶斷面(圖1)進行4季野外調查。其中,S1、S2斷面分別位于亞龍灣東西兩側,S3位于榆林港灣口內部,S4位于鹿回頭附近,S5、6分別位于天涯海角和南山寺附近開敞海區,6條斷面均避開游人活動區域,受人為破壞相對較小。另外,6條斷面中灣口和灣頂,遮蔽型和開敞型海岸均有涉及,以求更全面地反映出三亞潮間帶的綜合環境特征。采樣分別于2008年1月、4月、8月和11月的大潮日進行,使用GPS定位斷面位置,每條斷面分高、中、低潮帶進行采集。定量樣品在每個潮帶隨機選取3—4個25 cm×25 cm樣方。樣方內海藻使用鏟刀從附著根部刮下,清除附著沙石后裝入塑封袋避光保存并做好標號。同時在各斷面附近分潮帶廣泛采集定性樣品,一并帶回實驗室進行分析鑒定。樣品稱重、計算和資料整理均按《海洋調查規范》[17]進行。
1.3 數據處理
采用優勢度指數Y確定大型海藻群落生物量優勢種類[18]:
使用以下3個多樣性指數來表征大型海藻群落的多樣性[19]:

式中,ni為種i在各斷面的生物量總和;N為所有種的生物量總和;fi為種i在各斷面的出現頻率;S為采集生物的總種類數;Pi為第i種的生物的生物量占總生物量的比例。
采樣斷面圖使用ArcGIS 9.3繪制。在SPSS13.0軟件中采用Kolmogorov-Smirnov檢驗和Levene檢驗分別對數據進行正態性和方差齊性假設檢驗。如果數據同時滿足正態性和方差齊性假設,則對4季生物量以及多樣性指數值進行單因素方差分析(One-Way ANOVA),并進行Tukey′s post-hoc多重比較以檢驗季節間的生物量與多樣性差異,顯著性水平α=0.05。
聚類分析(CLUSTER)等多元統計分析方法比多樣性指數更能靈敏地反應群落結構的變化[20]。方法如下:首先為降低生物量上占優勢的個別物種對群落結構影響的權重,對原始生物量數據進行四次方根轉換,而后建立不同物種的個體生物量在不同斷面間的數據矩陣,并基于Bray-Curtis相似性指數構建各斷面間的相似性矩陣,最后采用組平均法構建群落聚類圖[21],群落的多元分析使用PRIMER 6.0軟件包。另外,為便于分析群落結構,本文規定若斷面間相似度>40%,可認為其較為相似。
種類的溫度性質劃分主要依據分類學文獻和現有資料[22-25],并參考項斯端和阮積惠[5]的劃分方法。
圖1 三亞巖相潮間帶大型海藻采樣斷面圖Fig.1 Sampling sections for intertidal benthic macroalgae in Sanya
2 結果
2.1 種類組成的分布特征和季節變化
綜合定量和定性數據,三亞巖相潮間帶共鑒定出大型底棲藻類130種,其中紅藻門41屬67種(占總物種數的51.54%),是三亞巖相潮間帶底棲海藻的主要構成成分,褐藻門16屬31種(占23.85%)、綠藻門15屬29種(占22.31%)、藍藻門3屬3種(占2.31%)。名錄詳見附錄1。
從水平分布來看,各斷面底棲海藻的四季平均物種數排序為S1>S6>S5>S3>S4>S2,物種數最多的S1斷面平均有(21±7)種分布,而物種數最少的S2斷面僅(7±2)種(圖2)。其中,S1斷面常見的種有日本仙菜Ceramium japonicum、海柏 Polyopes polyideoides、波利團扇藻 Padina boryana、礪菜 Ulva conglobata等,S2、S3 斷面主要有波狀軟凹藻Chondrophycus undulates和寬扁叉節藻Amphiroa dilatata等,S4斷面主要有喇叭藻Turbinaria ornata、石花菜 Gelidium amansii等,S5、S6斷面以褐藻門海藻居多,如冠葉馬尾藻 Sargassum cristaefolium、瓦氏馬尾藻Sargassum vachellianum和波利團扇藻等。不同季節物種數的水平分布有所不同:春季種類數的最高值出現在S3斷面(17種),最低值出現在S5斷面(2種);夏季S1斷面的物種數最高(24種),而S4斷面最低(6種);秋季S1斷面的物種數繼續增加至29種,仍為6條斷面中物種最豐富者,其它5條斷面差別不大;冬季S5、S6斷面的物種數達到峰值,分別為25種和30種,而S2、S3、S4斷面的物種數最少,分別為6種、8種和6種。
從垂直分布來看,在調查區域的大部分斷面中,低潮帶種類數最多,主要有石花菜、日本仙菜、寬扁叉節藻和多種馬尾藻屬藻類。中潮帶的物種相對較少,包括礪菜、礁膜 Monostroma nitidum、苔狀鴨毛藻Symphyocladia marchantioides、大團扇藻Padina crassa等,而高潮帶幾乎沒有藻類分布(僅在秋季S1斷面出現了少量苔垢菜Calothrix crustacea,圖2中省略)。中潮帶與低潮帶有共同物種分布,這種現象在夏季最為普遍,各條斷面均有出現。
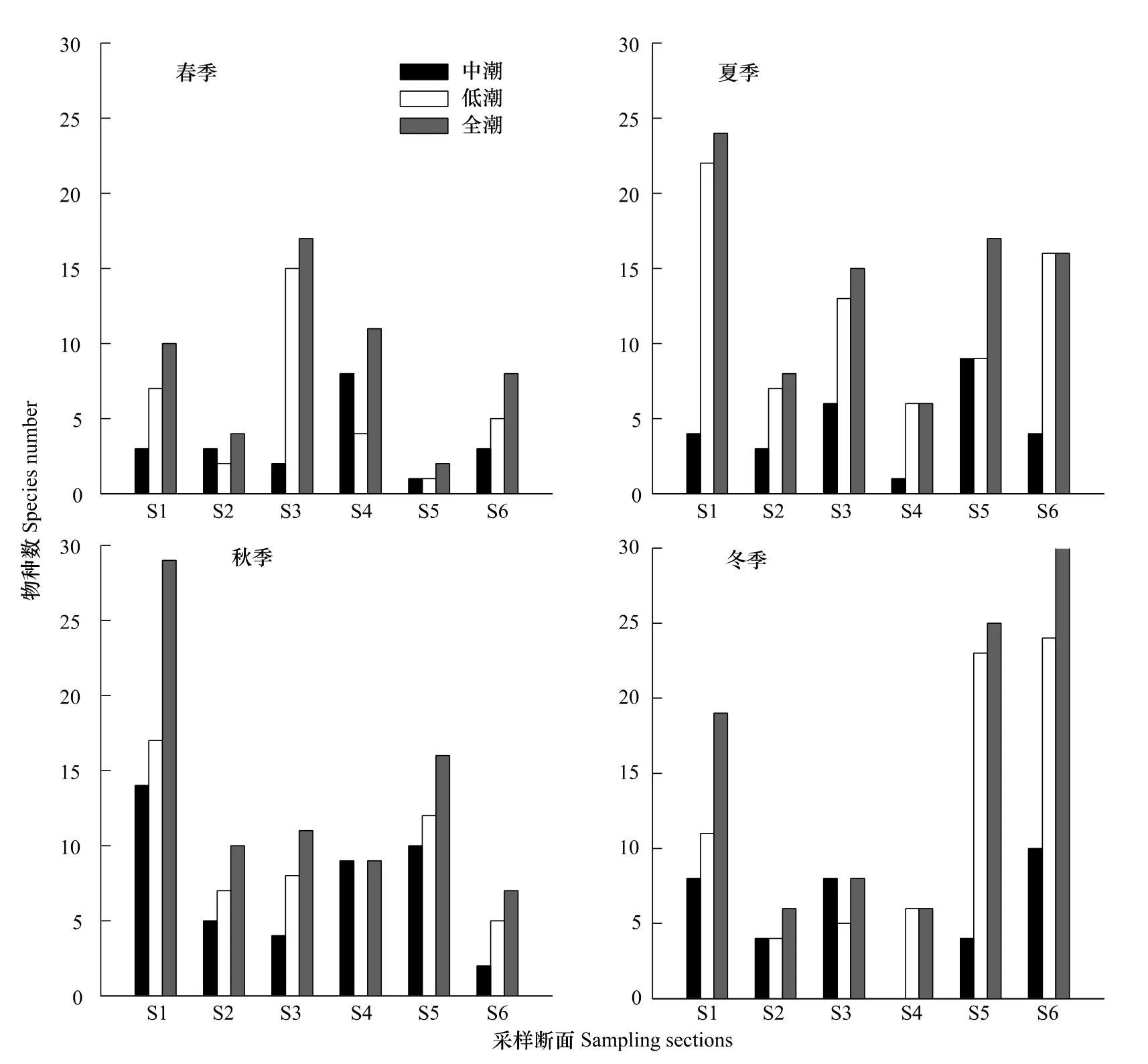
圖2 三亞巖相潮間帶底棲海藻物種數的四季水平分布Fig.2 The number of benthic algae species in different sampling sections of four seasons
底棲海藻物種數季節變化呈現夏、秋、冬3季多于春季的特征 (圖3),秋季(60種)>冬季(58種)>夏季(54種)>春季(37種),且秋、冬、夏3季差異不大。紅藻門種類數4季均占優勢,春季出現最少(15種);其次為褐藻門,春夏秋3季均占據次席。除紅藻門外,各門類種類數無明顯季節變化。
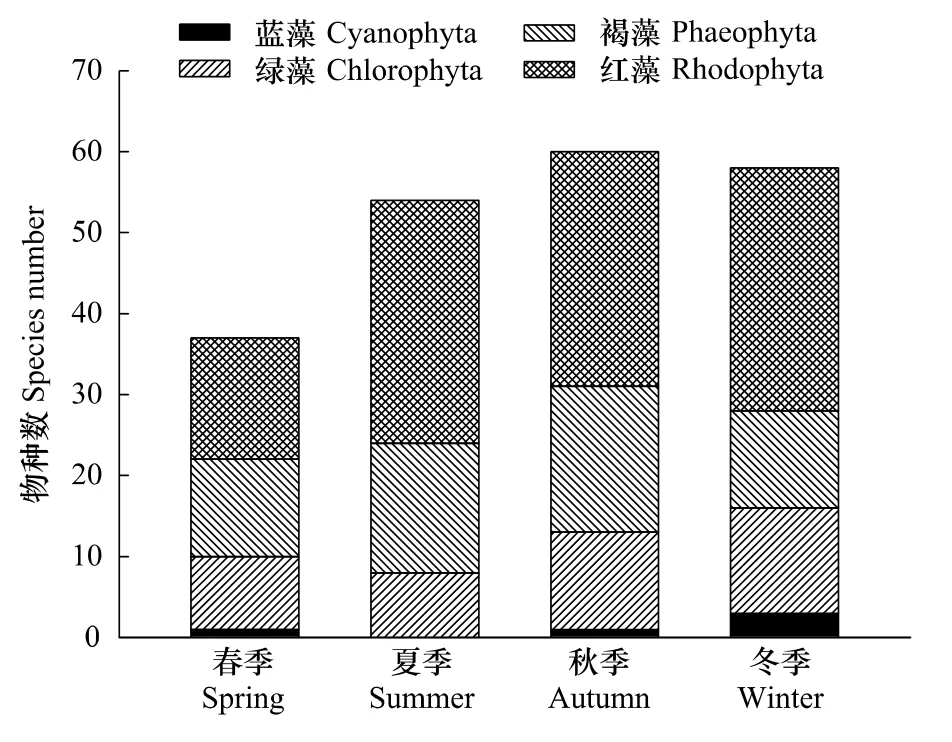
圖3 三亞巖相潮間帶不同類群底棲海藻的季節分布Fig.3 Seasonal distribution of benthic algae for different phyla in Sanya
2.2 種類的溫度性質
由表1可見,三亞巖相潮間帶大型底棲海藻區系全年以暖水性熱帶-亞熱帶種為主(占總種類的58.7%),其次為溫水性種類(占總種類的41.3%)。
分季節來看,暖水性種類在夏秋兩季分別占總物種數的60.4%和66.6%,溫水種類比例相對較低;相比之下,暖水性種類在春冬兩季出現的比例稍低為54.5%和49.1%。一些適溫較低的溫水種類,如刺松藻Codium fragile、節莢藻 Lomentaria hakodatensis、金膜藻Chrysymenia wrightii、扁江蘺Gracilaria textorii等僅在冬季出現。
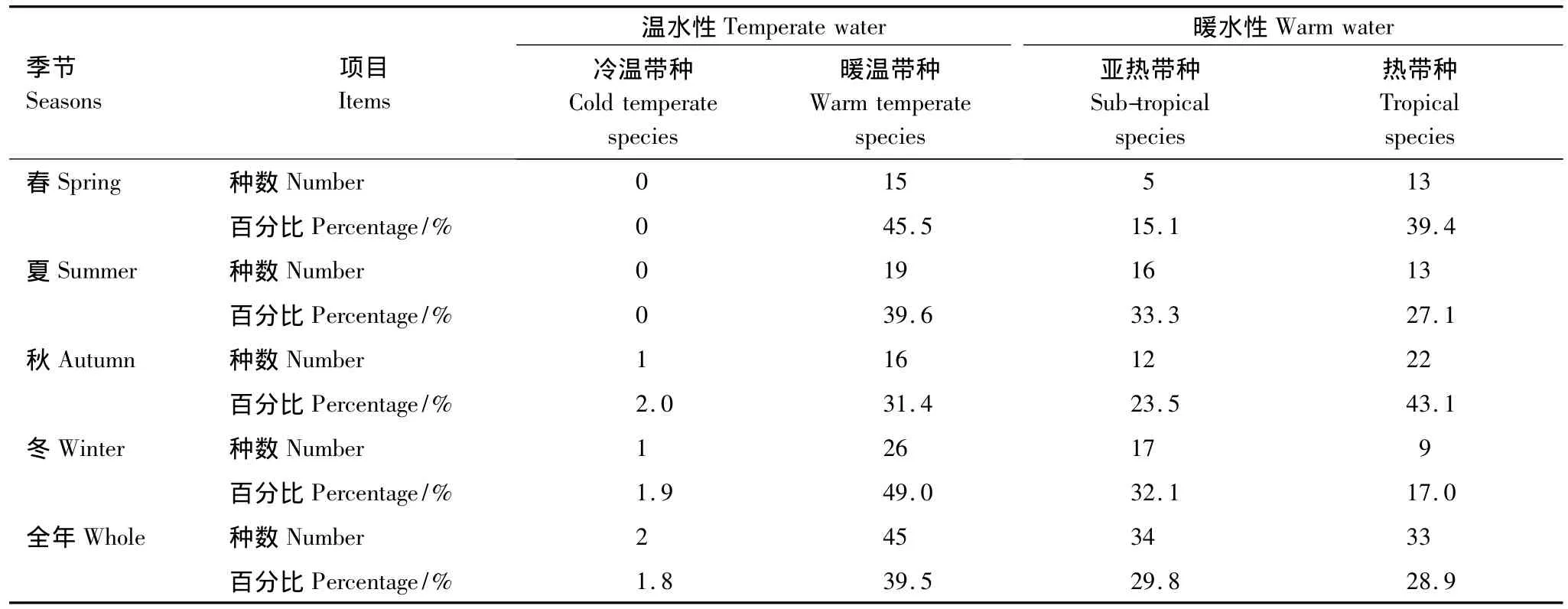
表1 三亞底棲海藻種類的溫度性質Table 1 Temperature features of benthic algae in Sanya
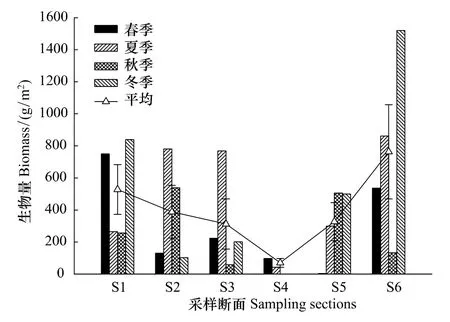
圖4 三亞巖相潮間帶底棲海藻生物量分布(mean±SD)Fig.4 Distribution of benthic algae biomass in Sanya
2.3 生物量的分布特征和季節變化
不同季節三亞巖相潮間帶大型底棲海藻生物量分布情況見圖4(S4斷面因海況不佳,缺秋冬季定量樣品)。從平面分布來看,各斷面大型海藻的4季平均生物量排序為:S6>S1>S2>S5>S3>S4,S6 斷面平均生物量達 (762.62±507.86)g/m2,而生物量最低的S4斷面,僅為(69.66±27.58)g/m2。從不同季節來看,4季的平均生物量有所不同,整個采樣區域4個季節的平均生物量排序為:冬季>夏季>秋季>春季。其中,春秋兩季平均生物量分別為(289.91±264.96)g/m2和(298.19±193.62)g/m2,而夏冬兩季分別為(502.58±312.56)g/m2和(632.78±513.03)g/m2,約為春秋兩季的兩倍,但統計結果顯示4季的生物量并無顯著差異(P=0.392,F=1.056)。
2.4 優勢種組成與季節變化
三亞巖相潮間帶大型海藻群落優勢種季節分布如表2所示。4季群落優勢種(Y≥0.02)由紅藻門和褐藻門物種組成。春、夏季各4種,分別占總生物量的73.10%和74.42%;秋季3種,占62.10%;冬季5種,占59.30%。其中波利團扇藻與日本仙菜是春冬兩季共有優勢種,波狀軟凹藻與冠葉馬尾藻是夏秋兩季共有優勢種,而春夏與秋冬并無共同優勢種。由4季群落優勢種的溫度性質表明,熱帶-亞熱帶性暖水種比例最大,優勢度較高,且4季均有分布。
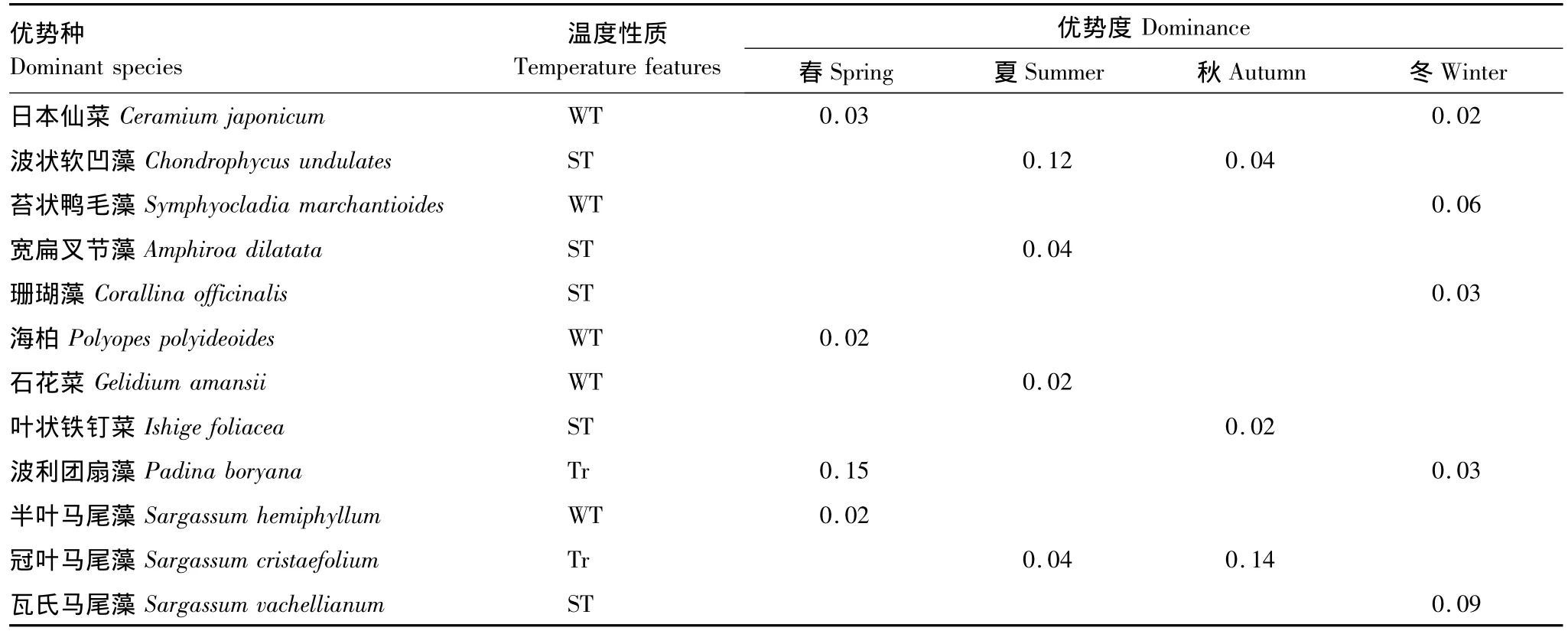
表2 三亞潮間帶底棲海藻優勢種四季分布及優勢度Table 2 Dominant species of benthic algae and its dominance in four seasons
2.5 物種多樣性指數
如表3所示,Shannon-Wiener物種多樣性指數均值的季節排序:冬季(2.02±0.38)>秋季(1.43±0.75)>春季(1.40±0.43)>夏季(1.13±0.65),但4個季節間無顯著差異 (P=0.166,F=1.901)。
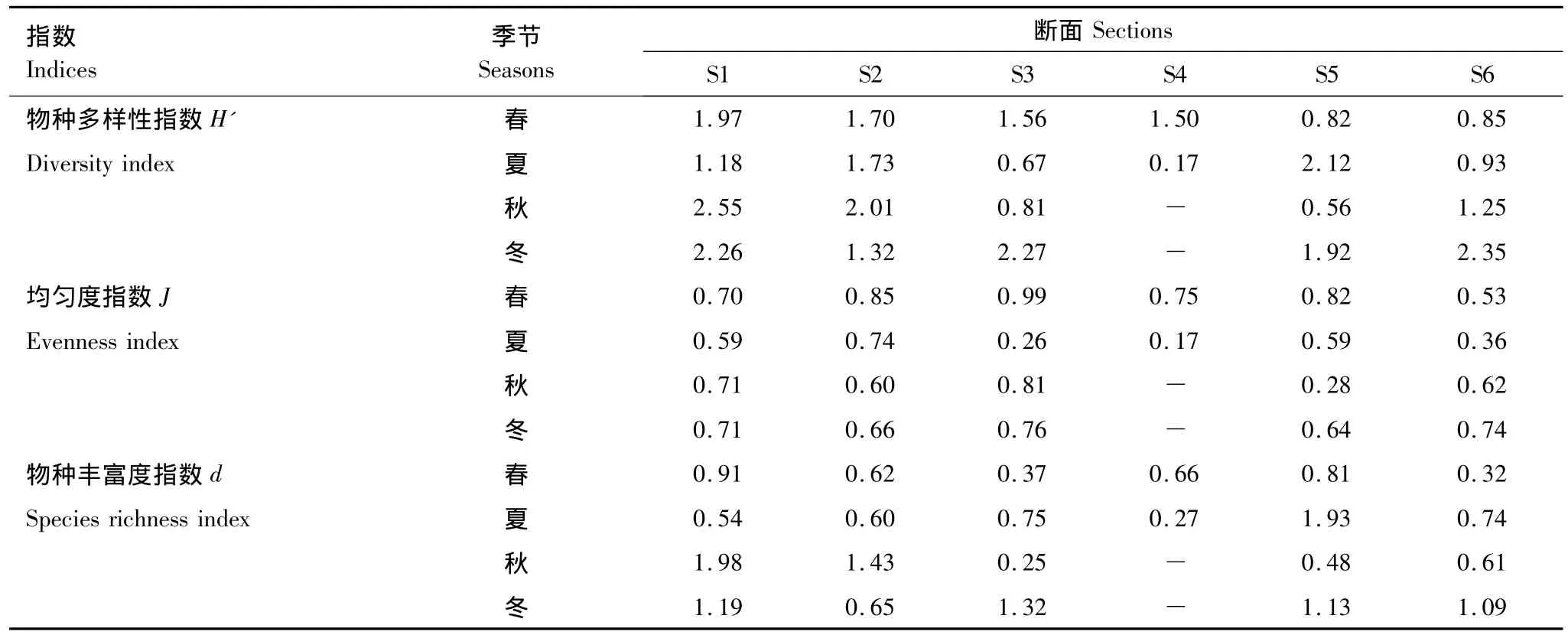
表3 三亞巖相潮間帶各斷面大型底棲藻類群落物種多樣性指數Table 3 Biodiversity indices of benthic algae among 6 sections in Sanya
Pielou均勻度指數均值的季節排序:春季(0.77±0.14)>冬季(0.70±0.05)>秋季(0.61±0.18)>夏季(0.46±0.20),4個季節間差異顯著,夏季明顯低于春冬兩季(P<0.05,F=3.812,Tukey′s test)。
Margalef物種豐富度指數的季節排序:冬季(1.07±0.23)>秋季(0.95±0.65)>夏季(0.80±0.53)>春季(0.61±0.21),但4個季節間無顯著差異(P=0.458,F=0.906)。
2.6 大型藻類群落結構和多元數據分析
聚類分析結果顯示,底棲海藻群落結構4個季差異較大:在相似度40%的水平上,春季,位于亞龍灣東西兩岸的斷面S1與S2相似性較高,均有較多波利團扇藻與礪菜出現;夏季,斷面S2與S3相似度較高,波狀軟凹藻在兩個斷面均有大量分布;秋季各斷面相似度較低,均小于40%;冬季相似度較高的斷面為S5與S6,對相似性貢獻較大的種有瓦氏馬尾藻和苔狀鴨毛藻等。
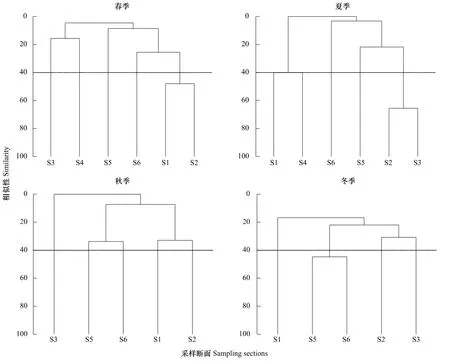
圖5 三亞巖相潮間帶底棲藻類聚類分析結果Fig.5 The results of CLUSTER in benthic algae community among the 6 stations in Sanya
3 討論
3.1 種類組成與區系的溫度性質
本次調查發現,三亞大型底棲海藻群落主要由紅藻類群構成,物種數最為豐富,其次是褐藻、綠藻和藍藻。這與之前的調查基本一致[11,15]。
從溫度性質來看,三亞巖相潮間帶大型底棲海藻區系與我國黃、渤海區以及浙江沿海有明顯差異,而與廣東沿海較為相近[5,24,26]。本次調查發現,盡管三亞位于海南島最南端,而三亞潮間帶海藻中熱帶種并沒有出現張水浸研究中的高比例[27],在58.7%的暖水種中,亞熱帶性和熱帶性的種類分布平均,本文與丁蘭平[24]、劉瑞玉[25]等的區系劃分稍有區別,更傾向于將三亞附近海區海藻歸為亞熱帶向熱帶過渡的區系。該海域是我國一些暖溫帶種的南限,又成為一些熱帶種的北限,形成了我國海藻南海南區獨特的物種分布現象。
從季節分布來看,大型底棲藻類物種數季節性變化不很明顯,但種類組成存在一定的季節差異。一些暖溫帶性種類僅在溫度相對較低的冬季出現,說明盡管三亞附近水溫季節變化不大,但冬季的水溫已經是一些溫帶種的適溫上限,這些種類難以在溫度稍高的其它3季生存。另外,季節性的水溫、光強變化可引起大型藻類在垂直分布上的遷移。尤其在夏季,由于水溫升高、光照強烈,部分常見于中低潮帶的海藻如角叉菜Choudrus ocellatus等喜陰的藻類會轉移到潮下帶生長。
從垂直分布來看,三亞地區氣溫高、光照強烈,長期暴露在空氣中的高潮帶幾乎沒有藻類生長,從中潮帶開始,一些典型的潮間帶海藻開始出現,低潮帶物種則更為豐富。另外,由于三亞附近海域潮差小,加之部分斷面坡度較緩,導致不同潮區的垂直界限相對模糊,一些延伸種從其占優的潮帶延伸到其他潮帶[2]。三亞巖相潮間帶的延伸種多出現在中潮帶下部和低潮帶上部,主要為波狀軟凹藻、冠葉馬尾藻、波利團扇藻等,這些藻類原本生長在低潮線附近[22-23],本次調查發現在中潮帶下部也有分布。
與該地區歷史數據比較,大型海藻種類數量并無明確增多或減少。1984年海南島全島沿海底棲海藻調查時,巖礁海岸記錄135種[11],與本次調查相差無多。而同期另一項針對三亞海藻的調查中[28],由于采樣區域小(調查區域位于大小東海、鹿回頭、天涯海角三亞港附近),僅記錄53種,遠少于本次調查發現的物種數。總的來說,現有對海南島大型底棲海藻的調查研究很少,針對三亞沿海的調查就更少,而且三亞位于海南島最南端,大型藻類種類分布最廣泛,豐富程度最高,分類學研究也最困難。基礎數據缺乏,很難對長時間尺度上大型海藻群落的變化做出明確的結論。
3.2 生物量分布以及群落結構特征
三亞潮間帶大型底棲海藻的生物量呈現出春秋兩季低、夏冬兩季高的趨勢。群落中的優勢種對生物量貢獻很大,其季節變化的特點為:四季差異較大,沒有四季共同優勢種出現,2個季節的共同優勢種有4種,但其優勢度在不同季節存在差異。如波狀軟凹藻是夏季第一優勢種,秋季其優勢地位則被冠葉馬尾藻取代,成為第二優勢種。波利團扇藻冬季優勢度并不高(0.03),春季卻成為第一優勢種。說明隨季節變化,它們在群落中的功能地位發生變化。造成這種變化的主要原因是:由于適溫性不同,一些熱帶和暖溫帶優勢種分別在不同季節進入生長盛期。例如:苔狀鴨毛藻是典型的暖溫帶性種類,其在山東沿海6—7月進入生長盛期[22],但在三亞冬季水溫較低時大量繁殖,生物量最高,成為冬季的優勢種。冠葉馬尾藻屬熱帶種類,每年3—4月開始長出幼芽,6—7月進入生長盛期,隨后大量繁殖[22],在夏秋兩季形成優勢種,而后逐漸衰退,群落中的優勢地位由其它藻類替代。瓦氏馬尾藻在浙江沿海4—6月進入生長盛期[22],但在三亞,1月時生物量已經很高,并能延續到春夏兩季。
從平面分布來看,海藻生物量和聚類分析結果與斷面所處經度并無明顯關系,而與斷面受風浪大小、巖相底質類型以及人為活動影響關系密切。因為這些因素往往決定了該斷面適宜生長的海藻種類,對大型海藻群落結構也會產生影響。例如在冬季,S5、S6斷面群落的相似度較高,很可能是因為兩斷面位于三亞灣西部,同屬于開敞型海區,受秋冬盛行的東北季風影響較大,適合個體粗大、喜風浪的馬尾藻屬海藻生長,如冠葉馬尾藻和瓦氏馬尾藻等。由于馬尾藻屬海藻通常在單位面積內具有較高的生物量,所以這也使S6斷面冬季生物量明顯高于其他斷面。另外,S6斷面潮間帶主要由大塊巖礁構成,且距岸較遠,人為活動少,也是其生物量高的原因之一。相反地,亞龍灣東岸內側的S2斷面以及位于榆林港內部的S3斷面受風浪影響相對最小,均適合一些喜靜水藻類生長,所以S2、S3斷面在西南季風盛行的夏季相似性較高。鹿回頭S4斷面平均生物量最小,這主要與人為活動有關。由于鹿回頭中低潮帶雜色蛤、團聚牡蠣等經濟貝類附著較多,人為采捕現象普遍,某種程度上破壞了藻類的生長環境,導致鹿回頭潮間帶大型海藻生物量偏低。
3.3 多樣性指數
從多樣性指數的平均值上看,冬季的Shannon物種多樣性和Margalef物種豐富度指數最高,這是因為,冬季三亞附近海區表層水溫介于亞熱帶種與熱帶種的最適溫區之間[29],是溫帶種的適溫上限,很多熱帶種和溫帶種共同出現。而Shannon多樣性和Pielou均勻度指數的最低值出現在夏季,這是因為,夏季水溫相對較高,不適宜一些溫帶種的生長,另外,夏季優勢度大于0.02的優勢種生物量占總生物量的74.42%,影響了其它藻類的生長。
與浙江附近海區巖相潮間帶相比[30],三亞潮間帶物種多樣性指數相對偏低,而總物種數卻遠遠高出。除了兩地的溫度差異外,也與兩地不同的底質環境有關:海南島巖相潮間帶大型海藻多數呈斑塊化、鑲嵌狀分布[11],而且熱帶地區大型海藻藻株分布面積大,通常一塊礁石上僅附著一種或幾種藻類,造成有限的樣方內種類相對單一,難以囊括該斷面所有大型海藻種類,導致三亞潮間帶多樣性指數相較其他海區偏低。而物種數中包含了斷面附近廣泛采集的定性樣本,物種數的豐富程度上遠高于浙江附近海區,這與現有資料一致[24]。
致謝:胡錫鋼、趙永強、江志兵等采集樣本,孫建璋、胡月妹等協助鑒定部分樣品,特此致謝。
Reference:
[1] Shou L,Gao A G,Zeng J N,Chen Q Z,Liao Y B,Xu X Q.The influence of the sediment environment on distribution of macrobenthos of the intertidal zone in Qushan Island.Chinese Journal of Zoology,2007,42(3):79-83.
[2] Zhang S Y,Liang J,Wang Z H,Wang K.Distribution characteristics of benthic algae in intertidal zone of Ma′an Archipelago of Zhejiang Province.Chinese Journal of Applied Ecology,2008,19(10):2299-2307.
[3] Zou D H,Gao K S.Effects of elevated CO2concentration on the photosynthesis and related physiological processes in marine macroalgae.Acta Ecologica Sinica,2002,22(10):1750-1757.
[4] Bruno F J,Boyer E K,Duffy J E,Lee C S,Kertesz S J.Effects of macroalgal species identity and richness on primary production in benthic Marine communities.Ecology Letters,2005,8(11):1165-1174.
[5] Xiang S D,Ruan J H.Checklist and the floral analysis of benthic seaweeds in Zhejiang Province.Journal of Zhejiang University(Science Edition),2002,29(5):548-557.
[6] Sun J Z,Ning X R,Le F F,Chen W D,Zhuang D G.Long term changes of biodiversity of benthic macroalgae in the intertidal zone of the Nanji Islands.Acta Ecologica Sinica,2010,30(2):106-112.
[7] Gaudêncio M J,Cabral H N.Trophic structure of macrobenthos in the Tagus estuary and adjacent coastal shelf.Hydrobiologia,2007,587(1):241-251.
[8] Diaz-Pulido G,Harii S,Mccook L J,Hoegh-Guldberg O.The impact of benthic algae on the settlement of a reef-building coral.Coral Reefs,2010,29(1):203-208.
[9] Soko?owskia A,Wo?owicza M,Asmusb H,Asmusb R,Carlierc A,Gasiunaitéd Z,Grémaree A,Hummelf H,Lesutienéd J,Razinkovasd A,Renaudg P E,Richardh P,K?drai M.Is benthic food web structure related to diversity of Marine macrobenthic communities?Estuarine,Coastal and Shelf Science,2012,108:76-86.
[10] Yang Z D.Primary studies on community ecology of benthic algae in Hainan Island.Studia Marina Sinica,1978(14):129-137.
[11] Jiang F K,Wu J F.Benthic algae of littoral zone in Hainan Island.Nanhai Studia Marina Sinica,1992(10):231-236.
[12] Mao L J,Zhang Y Z,Zhang Z K,Wei L,Ji X M,Zhu D K.Characteristics of sedimentary environments in Sanya bay of Hainan Island.Marine Geology& Quaternary Geology,2007,27(4):17-22.
[13] deBoer F W,Prins H T.Human exploitation and benthic community structure on a tropical intertidal flat.Journal of Sea Research,2002,48(16):225-240.
[14] Huang L M,Tan Y H,Song X Y,Huang X P,Wang H K,Si Z,Dong J D,Chen R Y.The status of the ecological environment and a proposed protection strategy in Sanya Bay,Hainan Island,China.Marine Pollution Bulletin,2003,47(1/6):180–186.
[15] Wang G Y,Wu H L,Yao X M,Feng Y Q,Wen Z,Xie G L,Peng H.Common bottom perch seaweed in Hainan Island.Journal of South China University of Tropical Agriculture,2010,1(2):175-182.
[16] Wang Y,Shen J W,Long J P.Ecological-sedimentary zonations and carbonate deposition,Xiaodonghai Reef Flat,Sanya,Hainan Island,China.Science China Earth Science,2011,41(3):362-374.
[17] Standardization Administration of the People′s Republic of China.GB/T 12763.6—2007.Specifications for oceanographic survey--Part 6:Marine biological survey.Beijing:China Standards Press.2007.
[18] Zhao Y J,Chen Q Z,Zeng J N,Shou L,Gao A G,Liao Y B.Distribution characteristics of intertidal zone macrozoobenthos in autumn and winter in Chemical Industrial Park and adjacent area,Jiaojiang Estuary.Acta Ecologica Sinica,2009,29(3):1578-1586.
[19] Sun J,Liu D Y.The application of diversity indices in Marine phytoplankton studies.Acta Oceanologica Sinica,2004,26(1):62-75.
[20] Zhou G,Zhang Z N.Rationale of the multivariate statistical software PRIMER and its application in benthic community ecology.Periodical of Ocean University of China,2003,33(1):58-64.
[21] Yang Q L,Lin G M.A multivariate analysis of variability of phytoplankton distribution in Xiamen seas during summer bloom period.Acta Ecologica Sinica,2007,27(2):465-476.
[22] Sun J Z.Anthology Research on Shellfish and Algae.Beijing:Marine Press,2006.
[23] Xia B M.Flora Algarum Marinarum Sinicarum,Tomus ii Rhodophyta.Beijing:Science Press,2004.
[24] Ding L P,Huang B X,Xie Y J.Advances and problems with the study of Marine macroalgae of China seas.Biodiversity Science,2011,19(6):798-804.
[25] Liu R Y.Checklist of Marine Biota of China Seas.Beijing:Science Press,2008.
[26] Zhao S F,Sun H J,Yuan Z J,Yang K.Resources of benthic algae from intertidal zone on Naozhou Island in spring.Journal of Shanghai Fisheries University,2006,15(3):322-327.
[27] Zhang S J.The species and distribution of seaweeds in the coast of China seas.Biodiversity Science,1996,4(3):139-144.
[28] Zhu H X,Li W X,Ding Z F.Resource investigation on macroalgae in Sanya,Hainan.Journal of Zhanjiang Ocean University,1989,9(1/2):116-120.
[29] Li W X,Zhu Z J,Liu F X.Phycology Introduction.Shanghai:Shanghai scientific& Technical Publishers,1982,232-235.
[30] Peng X,Xie Q L,Li S L,Chen S B,Qiu J B,Zhou Z M.Study on spatiotemporal distribution of intertidal benthic macro-algae and their diversity in southern Zhejiang Province.Journal of Tropical Oceanography,2010,29(3):135-140.
[1] 壽鹿,高愛根,曾江寧,陳全振,廖一波,徐曉群.底質環境對浙江衢山島潮間帶大型底棲動物分布的影響.動物學雜志,2007,42(3):79-83.
[2] 章守宇,梁君,汪振華,王凱.浙江馬鞍列島海域潮間帶底棲海藻分布特征.應用生態學報,2008,19(10):2299-2307.
[3] 鄒定輝,高坤山.高CO2濃度對大型海藻光合作用及有關過程的影響.生態學報,2002,22(10):1750-1757.
[5] 項斯端,阮積惠.浙江底棲海藻及其區系分析.浙江大學學報(理學版),2002,29(5):548-557.
[10] 楊宗岱.海南島潮間帶底棲海藻群落生態的初步研究.海洋科學集刊,1978(14):129-137.
[11] 蔣復康,吳進鋒.海南島沿海的底棲海藻.南海海洋科學集刊,1992(10):231-236.
[12] 毛龍江,張永戰,張振克,魏靈,季小梅,朱大奎.海南島三亞灣現代沉積環境.海洋地質與第四紀地質,2007,27(4):17-22.
[15] 王紅勇,吳洪流,姚雪梅,馮永勤,文珍,謝桂連,彭惠.海南島常見的大型底棲海藻.熱帶生物學報,2010,1(2):175-182.
[16] 王月,沈建偉,龍江平.海南島三亞小東海珊瑚礁坪生態沉積分帶和碳酸鹽沉積作用.中國科學:地球科學,2011,41(3):362-374.
[17] 中國國家標準化管理委員會.GB/T 12763.6—2007.海洋調查規范——第6部分:海洋生物調查.北京:中國標準出版社,2007.
[18] 趙永強,陳全震,曾江寧,壽鹿,高愛根,廖一波.椒江口化工園區及其鄰近區域潮間帶大型底棲動物秋冬季分布特征.生態學報,2009,29(3):1578-1586.
[19] 孫軍,劉東艷.多樣性指數在海洋浮游植物研究中的應用.海洋學報,2004,26(1):62-75.
[20] 周紅,張志南.大型多元統計軟件PRIMER的方法原理及其在底棲群落生態學中的應用.中國海洋大學學報,2003,33(1):58-64.
[21] 楊清良,林更銘.廈門海域浮游植物夏季赤潮期間分布變異的多元分析.生態學報,2007,27(2):465-476.
[22] 孫建璋.孫建璋貝藻類文選.北京:海洋出版社,2006.
[23] 夏邦美.中國海藻志第二卷第三冊紅藻門.北京:科學出版社,2004.
[24] 丁蘭平,黃冰心,謝艷齊.中國大型海藻的研究現狀及其存在的問題.生物多樣性,2011,19(6):798-804.
[25] 劉瑞玉.中國海洋生物名錄.北京:科學出版社,2008.
[26] 趙素芬,孫會強,袁振江,楊琨.硇洲島春季底棲海藻資源.上海水產大學學報,2006,15(3):322-327.
[27] 張水浸.中國沿海海藻的種類與分布.生物多樣性,1996,4(3):139-144.
[28] 朱華賢,李偉新,丁鎮芬.海南省三亞海藻資源調查.湛江海洋大學學報,1989,9(1/2):116-120.
[29] 李偉新,朱仲嘉,劉鳳賢.海藻學概論.上海:上海科學技術出版社,1982,232-235.
[30] 彭欣,謝起浪,李尚魯,陳少波,仇建標,周志明.浙南潮間帶大型底棲藻類時空分布及多樣性研究.熱帶海洋學報,2010,29(3):135-140.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
