人工高效經營雷竹林CO2通量估算及季節變化特征
2013-12-25 06:36:39陳云飛周國模
生態學報 2013年11期
陳云飛,江 洪,2,*,周國模,楊 爽,陳 健
(1.浙江農林大學浙江省森林生態系統碳循環與固碳減排重點實驗室,浙江 311300;2.南京大學國際地球系統科學研究所,南京 210093)
溫室氣體排放使得地球生物化學循環碳收支失衡,森林生態系統在吸收CO2維持生物圈碳氧平衡過程中作用巨大,可計量的森林碳匯日益成為國際研究的熱點。研究生物圈的碳循環機理和全球陸地生態系統碳收支及其對環境變化的響應已成為當今一系列大型國際計劃共同關注的核心問題,科學界急切的要揭示“碳失匯”問題[1],這部分未知的碳匯有待研究。CO2和H2O通過植被的光合作用和呼吸作用在土壤-植被-大氣圈空間層次上時刻進行著交換,以渦度相關技術為主體對土壤-植被-大氣間的CO2/H2O和能量通量以及生態系統碳水循環的關鍵過程進行長期連續的觀測,所獲取的觀測數據將被用來量化和對比分析研究區域內的生態系統碳收支與平衡特征及其對環境變化的響應[2]。目前渦度相關技術已在全球范圍內廣泛的應用于陸地生態系統的碳通量吸收與排放測定中,并取得了很好的成效,該方法已成為國際通量觀測網(FLUXNET)的標準方法。中國陸地生態通量觀測網絡(ChinaFLUX)也已經利用該技術開展了廣泛的觀測[3-4]。
亞熱帶季風氣候分布在北緯25°—35°亞熱帶大陸東岸地區,分布地區包括日本、韓國,美國、巴西的部分地區,中國的秦嶺-淮河以南的區域,并以中國面積最大。在亞熱帶通量觀測的系統還很有限,只有江西千煙州人工針葉林觀測站站以及哀牢山地常綠闊葉林觀測系統,兩站點南北跨度大僅對兩大植被類型進行了觀測,而亞熱帶植被類型以常綠闊葉林為主,混雜有落葉林、針葉林、混交林和竹類,亞熱帶季風區域植被類型豐富,同時多丘陵,森林覆蓋率高,有巨大的碳匯潛力和重要的碳源匯角色。竹林的通量觀測正是對亞熱帶通量觀測的補充和深入研究。
雷竹(Phyllostachysviolascens)作為經濟竹類在長江以南均有種植,且種植面積日益擴大。竹林采伐期短,更新代謝快,高效經營,收獲竹筍經濟效益顯著,同時應對氣候變化,增加森林碳匯有著積極的意義。獨特的管理方式和生理生態過程使其兼具森林、農田兩大系統的特點,雷竹林生態系統較一般森林,作為常綠樹種全年光合能力強,采伐期短更新代謝快,經濟林型種植面積廣大。很多研究表明,青幼年森林的碳吸收能力要強于衰老森林。北美黑云杉的采伐對碳源匯的影響[5],EOBS站點為采伐前林分年齡105a微弱碳匯(6±4)Cg·m-2·a-1, HBS75站點青幼年林型林分年齡33a為強的碳匯(143±35)Cg·m-2·a-1。因此,從生物學角度雷竹始終為青幼年的林齡,應具有較強碳匯潛力,但竹林的通量過程與固碳能力國內外文獻報道很少,有待于深入研究。在亞熱帶地區建立長期連續觀測的通量塔,對揭示竹林碳源匯過程以及影響碳平衡機制有深刻意義,對區域尺度下特色生態系統的碳平衡研究有啟發指導作用。
本文以浙江省臨安太湖源雷竹林生態系統觀測塔的一年通量數據為依據,結合雷竹高效經營覆蓋耕作科學的管理措施[6]。分析雷竹林系統CO2通量變化過程和碳收支量,及其人為干擾和環境因子對CO2通量變化的影響。
1 材料與方法
1.1 觀測站點
試驗區位于浙江省臨安市太湖源鎮,地理位置N 30°18′169″ E119°34′104″屬亞熱帶季風氣候,溫暖濕潤,雨熱同期,氣候特征春季多雨,夏濕熱有梅雨期,秋氣爽,冬干冷。全年降水量1600mm,年平均氣溫16℃,全年日照時數1900h。試驗地土壤以紅壤為主,海拔185m,坡向北偏東35°,坡度2—3°,雷竹林建園歷史10 a。觀測塔下雷竹林群落平均高度4.5m,平均胸徑4cm,以2、3年生竹為主,總蓋度80%,立竹密度每畝1500株,林下灌木草本很少,有竹葉糠皮覆蓋,為人工經營的筍用經濟林。雷竹是禾本科竹亞科剛竹屬竹種,由于早春打雷即出筍而得名,實為早竹之變種。自然狀態下雷竹,稈高7—11m,徑達4—6cm。節間較短而均勻,幼竿密被白粉生長季以1a為1個周期,連年出筍能力強,在出筍期與筍芽分化期要求有充足降水[7]。
人工高效經營的雷竹主要措施是充足肥料供給、充足水分灌溉、鉤梢剪枝、間伐老竹、覆蓋增溫等。其中冬季采用雙層法覆蓋增溫是關鍵技術,用礱糠、稻草雙層覆蓋,其厚度30cm(下面鋪稻草10—15cm,上面覆蓋礱糠15—20cm)。自然狀態下,2月下旬到3月上旬土壤溫度10℃以上,竹出筍最低溫度需9—10℃,最適溫度20—24℃[8],有效積溫200℃。4月份覆蓋的稻草腐爛入土,礱糠及時移除。覆蓋增溫后,11月底竹林覆蓋稻草礱糠保持地溫10℃以上,使雷竹提前發筍12月底開始萌動。1月中下旬大量出土,恰趕春節前后上市,價格較高。雷竹林每年都要收獲竹筍留養新竹,伐走等量的4a老竹,相當于間伐的管理措施,間伐量約占竹林的20%—30%,始終保持一定的立竹密度和1—3a的竹齡,這樣竹林生態系統處于發育的青幼時期,其碳匯能力較森林和農田系統有明顯優勢。
1.2 觀測儀器
觀測竹林建有高20m的微氣象觀測塔,開路渦度相關系統的探頭安裝在17 m高度上,由三維超聲風溫儀(CAST3, Campbell Inc., USA) 和開路CO2PH2O 分析儀(Li-7500, LiCor Inc., USA) 組成,原始采樣頻率為10Hz,數據傳輸給數據采集器(CR1000, Campbell Inc., USA) 進行存儲,同時根據渦度相關原理在線計算并存儲30min 的CO2通量(FC)、潛熱通量(LE)和顯熱通量(HS)等統計量。常規氣象觀測系統,包括3層風速(010C, met one, USA) 、3層大氣溫度和濕度(HMP45C,Vaisala, Helsinki, Finland),安裝高度分別為1m,5m, 17m,2個SI-111紅外溫度分別置于1.5m和5m,用于采集地表和冠層溫度,凈輻射儀(CNR4,Kipp&Zonen)傳感器安裝高度17m,用于采集上行/下行的長波/短波輻射、凈輻射的數據,此外還有土壤熱通量(HFP01,Hukseflux)觀測深度3cm,5cm;土壤含水量(CS616,Campbell,USA)觀測深度5,50,100cm;土壤溫度(109,Campbell,USA)觀測深度5,50,100cm。常規氣象觀測系統數據采樣頻率為0.5 Hz,通過數據采集器(CR1000,Campbell Inc,USA) 每30 min自動記錄平均風速、溫度、氣壓、凈輻射等常規氣象信息。常規氣象數據用于插補通量觀測數據的缺失。7層CO2/H2O廓線觀測系統(LI-840,Li CorInc,USA)觀測CO2/H2O從土壤到植被再到大氣的擴散過程,為閉路系統,采樣頻率為0.2Hz,同時輸出30min平均值。全部觀測數據保存到數據采集器的TF卡上,同時配置無線傳輸模塊利用遠程計算機加載虛擬串口通過手機信號連接數據采集器,可以實時監測傳感器狀態,30min數據可以實時傳輸。
1.3 研究方法
開路渦度系統數據采集器可在線計算虛溫訂正和空氣密度變化訂正。但是未考慮地形和儀器傾斜影響,當下墊面有傾斜度時,由于地球引力作用,順著山坡走向大氣會發生匯流、漏流現象,此時平均垂直風速并不為零。很多研究都是通過旋轉風向坐標軸來計算通量[9-11]。在中尺度大氣環流時,可以旋轉坐標軸迫使平均垂直風速為零[12]。根據風向、儀器底座、主風向地形坡度等建立一新的坐標軸參考系統,經過坐標旋轉后,凈生態系統交換量即為渦度相關通量、儲存項、水平和垂直方向的平流3項的和。常見的有二次坐標旋轉、三次坐標旋轉和平面擬合,本文采用二次坐標旋轉(DR),使坐標系x軸與平均水平風方向平行,從而使平均側向風速度和平均垂直風速度為零。
本文的觀測方法采用的是渦度相關的微氣象觀測方法,通過測定大氣中湍流運動產生的風速脈動和氣體濃度脈動,計算二者協方差求解通量值[13]。
凈生態系統碳交換量(NEE)主要是指生態系統中植物光合作用、植被冠層空氣的碳儲存和生態系統呼吸消耗的碳排放引起的生態系統碳儲量變化[14]
微氣象學研究以大氣為對象,當CO2從大氣進入到生態系統時,定義NEE符號為負;當CO2從生態系統排放到大氣中時,定義NEE符號為正。生態學家研究的重點在生態系統的變化,對于通量的符號規定為:氣體由大氣圈進入生態系統的通量值符號為負,反之為正[15]。
凈生態系統生產力(NEP),定義的符號剛好和NEE符號相反,生態系統總交換量(GEE)與生態系統總初級生產力(GPP)符號也是相反的,這樣陸地和大氣之間的氣體交換過程中的關系,可用下列方程描述:
GEE=NEE-RE
(2)
NEE=Fc+Fs
(3)
RE=REnight+REday
(4)
式中,Fc為大氣和生態系統冠層的碳通量,即渦度探頭觀測值,Fs為冠層內的碳儲存通量(雷竹林高度不足8m年際變化可忽略);式中RE生態系統呼吸包括植物自養呼吸以及土壤微生物分解土壤有機質和凋落物呼吸通量,分為白天與夜晚計算。夜間生態系統完全為CO2排放狀態,NEE數值上就等于生態系統呼吸值RE。白天的生態系統呼吸是通過擬合半小時時長的5cm土壤溫度數據與夜間NEE數據,建立溫度與生態系統呼吸的關系式,推算REday。根據公式(2),(4)。分解NEE可以分解成GEE和RE,進而計算出GEE。
1.4 數據校正與插補
基于渦度原理開路系統采集的10Hz原始數據,因為用紅外氣體儀觀測的CO2氣體濃度是相當于干空氣的質量混合比,大氣的溫度、壓力、濕度發生變化會引起CO2質量濃度的變化,需要根據理想氣體狀態方程校正氣體密度為摩爾質量比。需要進行水汽校正,即WPL校正。根據垂直平均風速為零假設,做坐標軸旋轉校正。以上校正應用EdiResoftwar設定參數,加載模塊,計算成為30min步長數據。EdiRe software是由愛丁堡大學編寫,專門處理渦度觀測數據,并做相關校正。
在實際觀測中由于受到降水、凝水、昆蟲以及隨機電信號異常等的影響,需要對通量數據進行質量控制,結合渦度相關法通量觀測原理和ChinaFLUX推薦篩選標準,目前通用的渦度數據處理,有平均日變化法(MDV),查表法(LookUp-table),非線性回歸法(NLR)。本文中應用非線性的經驗方程,光合有效輻射數據偏差較大,插補數據的結果不理想相關性很小,因此采用平均日變化法。平均日變化法是對缺失數據用相鄰幾天同時刻數據的平均值進行查補。該方法首先確定平均時段的長度,研究發現白天取14d、夜間取7d的平均時間長度時偏差最小[16]。
渦度相關開路系統數據易受降水、電信號、儀器故障影響造成數據缺失或者不合理,一般來說文獻報道的國際通量網絡各個站點數據缺失和不合理比例在17%—50%[17]。美洲通量網(AmeriFlux)的站點白天缺失和不合理數據在20%—35%[18]。本站點白天數據有效率在80%左右;夜間湍流不充分CO2過多的沉積在林冠下部不能被探頭檢測到,篩選風摩擦系數較大的數據,有效數據在48%左右。表1說明除8月份數據有效率偏低外,其余各月數據有效率要高于通量觀測網絡系統的平均水平。
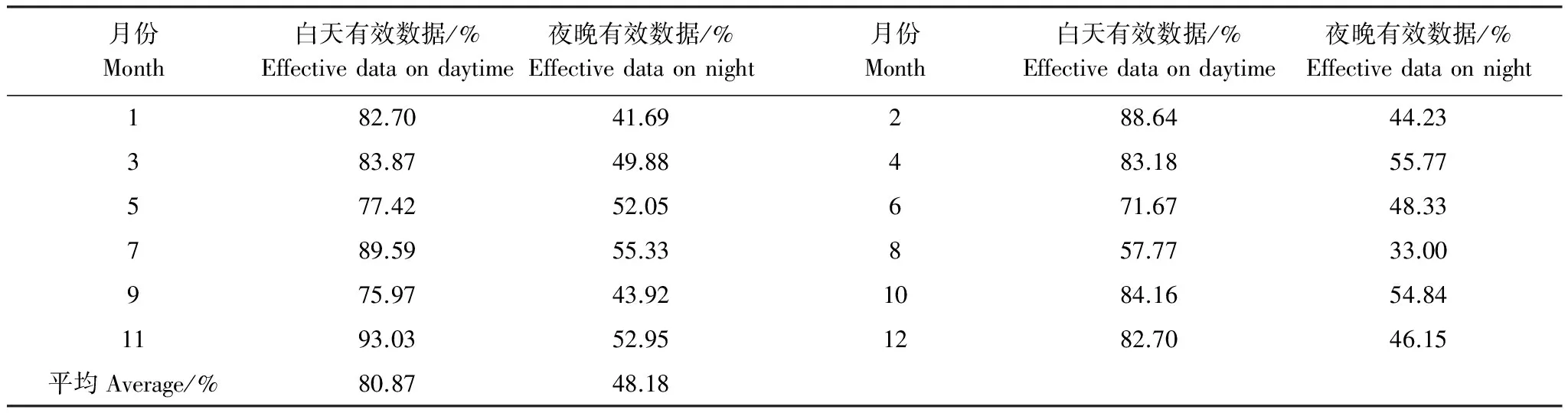
表1 各月有效數據統計
2 結果與分析
2.1 常規氣象因子的季節變化
觀測數據從2010年10月到2011年9月,構成全年數據。日平均最高溫2011年7月8日30.21℃,最低溫2011年1月17日-5.50℃。月平均氣象因子見表2。全年最高氣溫7月,最低氣溫1月;光合有效輻射變化與氣溫相同;降雨量最大6月,最小12月,降雨與該地區往期持平,降雨量變化較氣溫變化年際差異大。
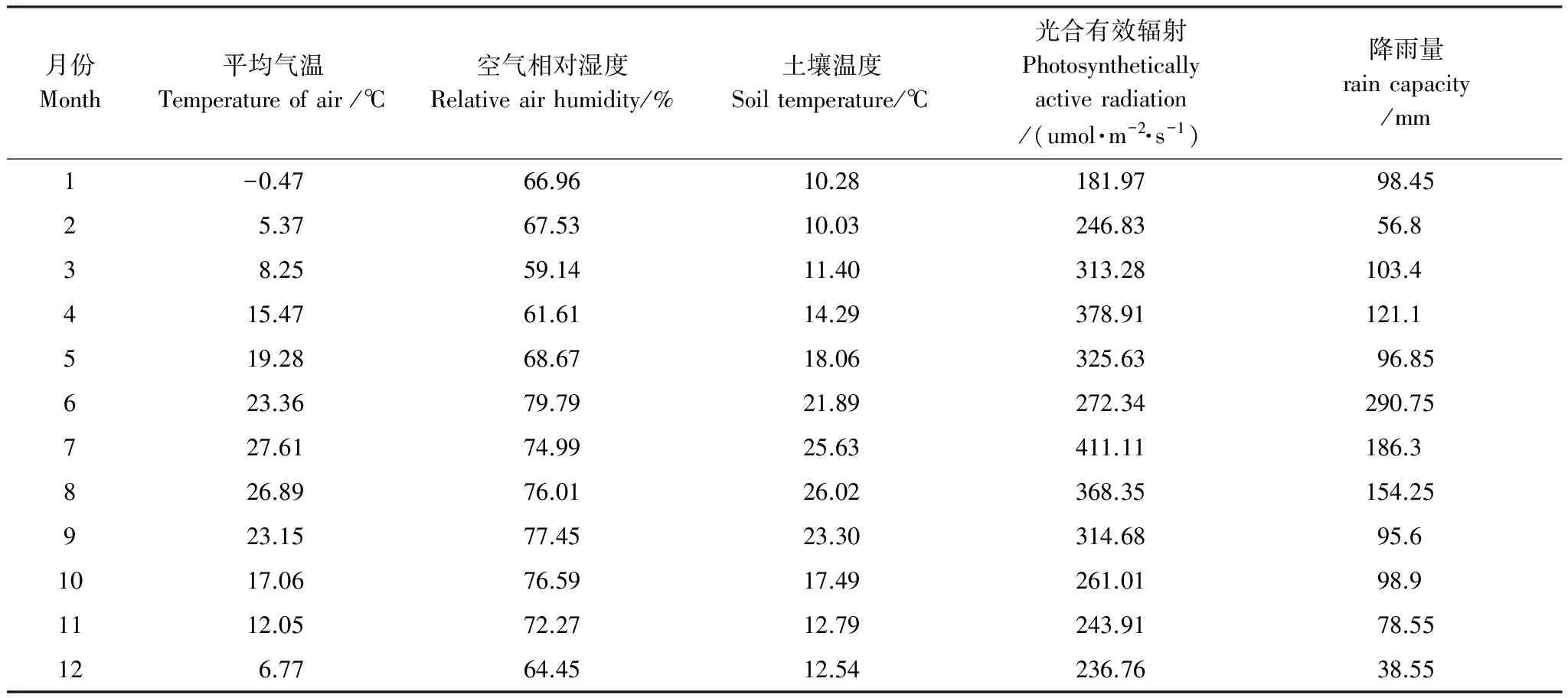
表2 各月氣象因子變化
2.2 凈生態系統碳交換量
從圖1月每時刻平均的日進程可以看出,NEE呈“U”形變化,表現為在白天通量負值,夜間通量正值,雷竹林生態系統月尺度上的NEE符號變化時刻和最大的碳排放、碳吸收時刻都有明顯的差異。各月通量符號變化,由正值轉為負值在日出1—1.5h,春冬季與夏秋季相比延遲1h;由負值轉為正值時刻較集中,發生在17:00—18:00。9—11月NEE由正轉負發生在6:30—7:00,NEE最大負值出現在11:00前后。在12月—翌年2月NEE由正轉負發生在7:30—8:00,NEE最大負值出現在12:00前后。3—8月的通量符號變化時刻和最大負值時刻介于秋季與冬季之間,NEE由正轉負發生在7:00—7:30,NEE最大負值出現在11:00—11:30。

圖1 CO2通量各月平均日變化
從全年NEE 變化看,年碳吸收總量126.303gC·m-2·a-1,總體是碳匯。這要小于周國模應用生物量調查法對毛竹固碳能力509gC·m-2·a-1,杉木為348gC·m-2·a-1的估算[19]。李靜潔等研究江西紅壤丘陵地區雙季稻碳收支[20],在不同施肥措施下估算早稻田兩季稻的碳匯能力為311—629gC·m-2·a-1,雷竹年固碳能力要小于水稻,同時也小于北方農田系統198—318gC·m-2·a-1 [21]年吸收能力。具體季節分析12—2月即冬季覆蓋月份NEE為正,雷竹林表現為碳源,春季、夏季、秋季均為碳匯。全年碳通量,吸收量表現為雙峰變化曲線,第1峰出現在6月(-21.39gCm-2·月-1);第二峰出現在11月(-35.89gC·m-2·月-1)。最高排放量1月(16.061gC·m-2·月-1)。
6月全月凈積累總量要小于11月,這可能是由于亞熱帶6月份開始進入梅雨期,影響雷竹的光合生產能力。同緯度氣候下千煙洲站點6月出現的凈積累量也小于11月的凈積累量[22]。7—8月受高溫影響有明顯的“午休”現象,CO2通量一般在10:0—11:00達到峰值,之后降低,09—11月適宜的水熱條件使碳吸收持續增加可為冬季竹筍萌發積蓄有機物。7—8月份雖然有較強的初級生產力,但由于呼吸過程排放的碳也比較大,碳凈吸收低于5—6月和9—11月。這在很多類型的生態系統普遍存在,在竹類系統也得到了驗證。12月進入冬季開始覆蓋增溫,人為經營措施干擾NEE開始減小。
從圖2可以看出RE呈單峰變化,在夏季較高冬季較低。圖5中顯示夜間NEE與土壤5cm溫度的散點圖,擬合曲線顯示夜間NEE與土壤5cm溫度成指數關系變化,相關系數0.3。7月土壤溫度最高月份,RE達到峰值151.05 gC·m-2·月-1,在氣溫較低的春冬季呼吸較低。從季節上分析,分為冬季(12—2月),春季(3—5月),夏季(6—8月),秋季(9—11月)。各季節RE占全年總量的比例分別為16.7%,21.2%,37.0%,25.1%;各季節NEE占全年比例分別為-21.3%,26.6%,25.8%,68.9%;各季節GEE占全年比例為12.8%,21.8%,35.8%,29.6%。全年RE貢獻最大為夏季,夏季生態系統呼吸是冬季的2.3倍;全年NEE貢獻最大為秋季超過全年其余月份之和,而冬季為負貢獻降低了全年碳匯總量,春夏季NEE貢獻為年平均水平;全年GEE最大貢獻為秋季,最小為夏季,變化與RE一致。
全年GEE季節變化特征明顯,峰值出現在6—7月,可能與溫度、水分影響有關;之后開始下降,但秋季生長明顯大于春季,冬季GPP最小。根據雷竹的生長特性,竹筍出土吐葉到成竹經30—40 d,5月大量新竹開始光合生產,6月下旬到7月梅雨期,為雷竹生長提供充足的水分供給。冬季的防雪鉤梢和溫度影響,可能是生產力開始下降的主要影響因素。覆蓋月份為12—3月,出筍是1—3月,竹筍在適宜的水熱條件下爆發性生長,竹筍出土呼吸加大,同時覆蓋后土壤和覆蓋物分解呼吸大大超出覆蓋前的土壤呼吸,有研究發現覆蓋后是覆蓋前的5—10倍[23],造成冬季CO2排放不降。土壤呼吸、覆蓋物呼吸以及植物呼吸都土壤溫度密切相關,1月溫度降低,生態系統呼吸隨之降低。6月GEE與RE同時上升出現峰值,新竹不斷增強的光合作用吸收了一定CO2,同時生態系統呼吸達到了較大值,造成NEE不為最高值。9月GEE與RE同時開始下降時,NEE開始增加,并在11月達到全年的最高峰值。在春季NEE隨GEE與RE的升高而增加,在秋季NEE隨GEE與RE的下降而增加,出現兩個峰值。由此可見人工高效經營下,溫度是主要的影響因子,光照次之。雷竹喜濕怕澇,在受水分脅迫時人工會及時補水排澇,水分影響這里先不考慮。

圖2 全年各月累積凈生態系統CO2交換量(NEE)、生態系統呼吸(RE)和總生態系統CO2交換量(GEE)變化過程
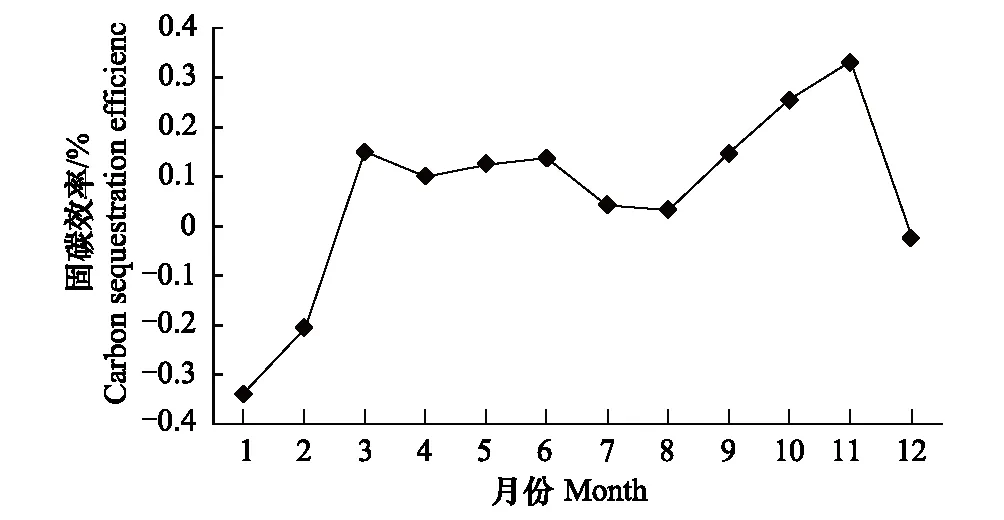
圖3 固碳效率變化
2.3 竹林固碳效率
在一定的時間尺度上,生態系統碳交換通量與碳固定和排放速率根據選定的研究界面和對象能夠相互直接轉化[24],在數值上GEE可等同為生態系統固碳速率,RE即為生態系統碳排放速率,NEE即生態系統凈固碳速率,量綱不變仍為gC·m-2·a-1。相對于植物生理研究,定義有碳利用效率(CUE),就是植物凈積累的碳占總光合生產中固定碳的比例[25],即有CUE=NPP/GEP[26],該指標表示植物碳同化能力的強弱,但是對于整個生態系統的碳收支研究缺失土壤呼吸和植物凋落物等異養呼吸碳排放計算,不能代表整個生態系統碳收支水平。如果將大氣-植被-土壤看作一個連續的整體,研究在生態系統氣體交換過程中生態系統的碳收支,將生態系統凈固碳速率與生態系統固碳速率比值,可以暫時定義為生態系統碳固定效率(ECSE),反映生態系統氣體交換中固定下來碳的比例,即ECSE=NEE/GEE。從圖3中看出,生態系統的碳利用效率從1月份開始為負值表明,是極強的碳釋放,冬季雷竹雖然有一定的光合能力但生態系統呼吸大大超過光合生產的凈積累量;3月開始,變為正值轉,為固碳并保持在10—16%;夏季7—8月固碳能力下降,碳排放的比例增加,進入秋季達到峰值,有生態系統氣體交換過程有33%被固定下來,全年固碳效率為11%。
2.4 高效經營覆蓋措施對CO2通量的影響

圖4 土壤5cm溫度與空氣1m溫度的比較
冬季覆蓋技術是雷竹高效經營的關鍵技術,覆蓋的主要作用是增加土壤溫度。夏季土壤表層溫度高于深層溫度,在冬季則相反,土壤溫度隨深度增加而升高。在冬季增加覆蓋措施后5cm土壤溫度要高于1m處氣溫5—10℃。從圖4中看出在12—2月,地表5cm土壤溫度明顯高出空氣溫度,平均溫度都維持在10℃以上,這就加大了生態系統的呼吸,同時冬季光合有效輻射為全年最低,形成冬季月份碳源。
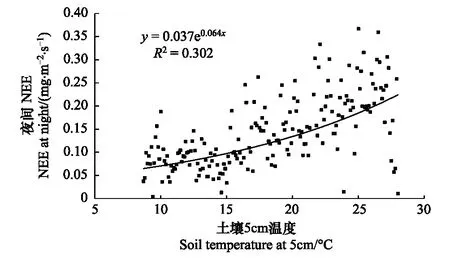
圖5 土壤5cm溫度變化與夜間NEE的關系
全年土壤溫度變化范圍9—26℃,波動范圍不大,夜間NEE可以代表生態系統呼吸即包括植物暗呼吸、土壤微生物呼吸以及凋落物分解的CO2排放,最高呼吸在0.6—0.7mg·m-2·s-1。全年的土壤5cm溫度與經摩擦風速篩選后夜間NEE關系并不明顯,因為點數太多,將各個溫度下的夜間NEE做平均處理,得到圖5。從圖5中,可以看出生態系統與土壤5cm成指數關系,但相關系數只有0.3,成指數關系這與之前有關文獻報道的研究結果較一致[16,27]。ChinaFLUX站點也有相似的報道,陳述悅研究華北麥田發現5cm地溫同土壤呼吸相關性最好[28],劉允芬等研究千煙洲人工針葉林土壤溫度與夜間NEE數值呈指數關系相關系數0.38[29]。相關系數不大的原因可能有兩個,首先冬季的覆蓋增溫措施干擾了土壤自然的呼吸,使溫度變化范圍減小,生物菌肥的使用加大了呼吸;再有溫度升高使植物自養呼吸增加,生態系統呼吸也隨之增加。
假定雷竹光合能力不變,數據顯示冬季無覆蓋時近地1m空氣溫度與土壤5cm溫度相差不大,用1m處空氣溫度代替無覆蓋的5cm土壤溫度,根據與夜間NEE的指數方程模擬無覆蓋的生態系統呼吸。以溫度為影響因子比較無覆蓋與有覆蓋的碳排放,模擬結果說明12—2月單位面積上,覆蓋后分別多增加21.56g,26.52g,20.53g的排放(表3)。因此若雷竹林無覆蓋可少排放CO2,相應冬季月份可由碳源轉匯。
冬季覆蓋增加的碳排放部分來自覆蓋的有機物分解。從表4中可以計算不包括地下部分生長計算碳收支平衡,其中雷竹每株干重計以3kg計算,每年每畝伐300株,地上部分碳密度參照周國模研究毛竹碳密度[19]以500g/kg計算,竹筍收獲以1500kg,含水90%計算,總年收獲量為525kg碳,即787.50Cg·m-2·a-1。竹筍收獲占14%,竹材收獲占86%,竹材的碳可以作為碳庫多年儲存,竹筍的碳運出系統后又將排放到大氣中,則雷竹林系統為十分微弱的碳源。雷竹林NEE較水稻田、北方農田都小,但收獲量缺明顯高出其他農田系統,可見高效經營措施對收獲量產出的影響。
覆蓋物投入的有機碳含量以及礱糠收回,稻草則腐爛入土的比例需要進一步實驗,對竹林系統碳庫影響都還不甚清楚。目前有關覆蓋物對雷竹林碳儲量影響、雷竹土壤有機質轉化研究的文獻還很少見,初步估計,每年每公頃投入覆蓋的稻草及礱糠等有機物料共100 t左右,大量的有機質輸入對土壤有機碳庫的組成分解及轉化應產生重要影響[30-32]。有文獻研究15a種植雷竹有機碳轉化變化,覆蓋經營后極大地提高了土壤有機質含量,每年每畝表層有機質增加達到616 kg,占投入物料的23.8%。該值略低于南方紅壤區作物秸稈的平均腐殖化系數(0.22—0.34,平均 0.28),可能是由于礱糠 C/N 比較高,不易腐殖化[33]。每畝1.6t有機碳投入,以23.8%轉化系數計算,將有380kg有機碳進入系統,即增加了碳排放571Cg·m-2·a-1,這些有機碳在系統中如何分配的有待深入研究。

表3 模擬覆蓋與無覆蓋碳排放變化

表4 高效經營中投入產出(以666.7m2計)
3 結論與展望
本文分析了太湖源人工經營的雷竹林碳通量各月的平均日變化趨勢,季節碳通量的變化趨勢,同時探究了冬季覆蓋增溫過程中對碳通量變化以及碳平衡的影響。研究表明:
人工經營的雷竹林生態系統全年各月碳通量平均日變化趨勢明顯;2010年10月到2011年9月碳收支為-126.303gC·m-2·a-1,RE為1108.845 gC·m-2·a-1,GEE為-1235.15gC·m-2·a-1。整體為碳匯,冬季覆蓋的12—2月份為碳源,其余月份為碳匯。通量過程具有明顯的季節性,雷竹林生態系統月尺度上的NEE符號變化時刻和最大的碳排放、碳吸收時刻都有明顯的差異,通量符號由正值轉為負值在日出1h—1.5h,春冬季與夏秋季相比延遲1h;由負值轉為正值時刻較集中,發生在17:00—18:00;最大碳匯時刻秋季在11:00,冬季在12:00,春夏介于11:00—12:00之間。溫度是影響碳通量的主要因子,5cm處土壤溫度與生態系統呼吸存在明顯的指數關系。在春季NEE隨NEE與RE的同時升高而增加,在秋季NEE隨NEE與RE的同時下降而增加,出現兩個峰值并有碳匯總量秋季大于春季。秋季竹林NEE貢獻達68.9%,同時固碳效率最高為33%。冬季覆蓋通過投入有機物料,提高地溫,同時增大了生態系統呼吸,增加了碳排放。
竹林具有很強的森林碳匯潛力,合理的耕作措施,科學采伐老竹,保持生態系統活力,可為陸地生態系統維持碳氧平衡應對氣候變化做出積極的貢獻。另一方面,對于竹林生態系統碳循環研究,本文僅對雷竹林的碳源匯做了結論,對與影響碳吸收、排放的各個分量未做計算。同時本站點僅對雷竹林一年的碳通量過程研究進行初步觀測研究,時間較短,因碳通量過程有明顯的年際變化易受氣候變化影響,要通過長期的連續觀測和相應的輔助實驗,來做深入研究。
[1]Tans P P,Fung I Y,Takahashi T.Observation constraints on the global atmospheric CO2budget.Science,1990,247(4949):1431-1438.
[2]Berbigier P,Bonnefond J M,Mellmann P.CO2and water vapour fluxes for 2 years above Euroflux forest site.Agricultural and Forest Meteorology,2001,108(3):183-197.
[3]Yu G R,SunX M.Flux Measurement and Research of Terrestrial Ecosystem in China.Beijing:Science Press,2008.
[4]Yu G R,Sun X M.Principles of Flux Measurement in Terrestrial Ecosystems.Beijing:Higher Education Press,2006:4-4.
[5]Payeur-Poirier J L,Coursolle C,Hank A,Giasson M A.CO2fluxes of a boreal black spruce chronosequence in eastern North America.Agricultural and Forest Meteorology,2012,153:94-105
[6]FangW,He J C,Lu K X,Chen J H.Cultivation techniques of early shooting and high yielding for Lei Bamboo Sprout.Journalof Zhejiang Forestry College,1994,11(2):121-128.
[7]Wang Z T,FangW,He J C.High Yield Cultivation Techniques of Lei Bamboo shoots.Beijing:China Forestry Press,1993.
[8]Xu L L,Hu Y B,Hu Y C,Sun C M,Yu M C,Liang K,Lang J B.Lei bamboo production development and covered technology.Shanghai Agricultural Science and Technology,2008,(2):85-86.
[9]Baldocchi D D,Hicks B B,Meyers T P.Measuring biosphere-atmosphere exchanges of biologically related gases with micrometeorological methods.Ecology,1988,69(5):1331-1340.
[10]McMillen R T.An eddy correlation technique with extended applicability to non-simple terrain.Boundary-Layer Meteorology,1988,43(3):231-245.
[11]FokenTh,Wichura B.Tools for quality assessment of surface-based flux measurements.Agricultural and Forest Meteorology,1995,78(1/2):83-105.
[12]Paw U K T,Baldocchic D D,Meyers T P,Wilson K B.Correction of eddy-covariance measurements incorporating both advective effects and density fluxes.Boundary-Layer Meteorology,2000,97(3):487-511.
[13]Reynolds O.On the dynamical theory of incompressible viscous fluids and the determination of the criterion.Philosophical Transactions of the Royal Society of LondonA,1895,186:123-164.
[14]Garratt J R.Limitations of the eddy-correlation technique for the determination of turbulent fluxes near the surface.Boundary-Layer Meteorology,1975,8(3/4):255-259.
[15]Song X,Liu Y F,Xu XF,Yu GR,Wen X F.Comparison study on carbon dioxide,water and heat fluxes of the forest ecosystem in red earth hilly zone over winter and spring.Resources Science,2004,26(3):96-104.
[16]FalgeE,Baldocchi D,Olson R,Anthoni P,Aubinet M,Bernhofer C,Burba G,Ceulemans R,Clement R,Dolman H,Granier A,Gross P,Grünwald T,Hollinger D,Jensen N O,Katul G,Keronen P,Kowalski A,Ta Lai C,Law B E,Meyers T,Moncrieff J,Moors E,Munger J W,Pilegaard K,Rannik ü,Rebmann C,Suyker A,Tenhunen J,Tu K,Verma S,Vesala T,Wilson K,Wofs S.Gap filling strategies for defensible annual sums of net ecosystem exchange.Agricultural and Forest Meteorology,2001,107:43-69.
[17]Katul G,Lai C T,Sch?fer K,Vidakovic B,Ellsworth D,Oren R.Multiscale analysis of vegetation surfacefluxes:from seconds to years.Advances in Water Resources,2001,24(9/10):1119-1132.
[18]Wilson K B,Baldocchi D D.Comparing independent estimates of carbon dioxide exchange over 5 years at a deciduous forest in the southeastern United States.Journal of Geophysical Research,2001,106(D24):34167-34178.
[19]Zhou G M.Carbon Storage,Fixation and Distribution in Mao Bamboo(Phyllostachyspubescens) Stands Ecosystem.Zhejiang:Zhejiang University,2006.
[20]Li J J,Pan G X,Li LQ,Zhang X H.Estimation of net carbon balance and benefits of rice-rice cropping farm of a red earth paddy under long term fertilization experiment from Jiangxi,China.Journal of Agro-Environment Science,2009,28(12):2520-2525.
[21]LiJ,YuQ,Sun X M,Tong X J,Ren C Y,Wang J,Liu E M,Zhu Z L,Lin G R.The north China plain farmland ecological system and its environmental regulation mechanism carbon exchange.Science in ChinaSeries D:Earth Sciences,2006,36(Supplement):210-223.
[22]Liu Y F,Song X X,Sun X M,Wen X F,Chen Y R.CO2flux seasonal changes and environmental factors of influence form Qianyanzhou of Coniferous.Science in ChinaSeries D:Earth Sciences,2006,36(Supplement):109-117.
[23]Zhou G M,Jiang P K,Xu Q F.Bamboo Forest Ecosystem Carbon Fixation and Transformation.Beijing:Science Press,2010.
[24]Yu G R,Wang Q F,Liu Y C,Liu Y H.Conceptual framework of carbon sequestration rate and potential increment of carbon sink of regional terrestrial ecosystem and scientific basis for quantitative carbon Authentification.Progress in Geography,2011,30(7):772-774.
[25]Cabral O M R,Gash J H C,Rocha H R,Marsden C,Ligo M A V,Freitas H C,Tatsch J D,Gomes E.Fluxes of CO2above a plantation of Eucalyptus in southeast Brazil.Agricultural and Forest Meteorology,2011,151(1):49-59.
[26]ChoudhuryB J.Modeling radiation-and carbon-use efficiencies of maize,sorghum,and rice.Agricultural and Forest Meteorology,2001,106(4):317-330.
[24]LloydJ,Taylor J A.On the temperature dependence of soil respiration.Functional Ecology,1994,8(3):315-323.
[27]LloydJ,Taylor J A.On the temperature dependence of soil respiration.Functional Ecology,1994,8(3):315-323.
[28]ChenSS,LiJ,Lu P L,Wang Y H,Yu Q.Soil respiration characteristics in winter wheat field in North China Plain.Chinese Journal of Applied Ecology,2004,15(9):1552-1560.
[29]Liu Y F,Yu G R,Wen X F,Wang Y H,Song X,Li J,Sun X M,Yang F T,Chen Y R,Liu Q L.The season of CO2flux variation characteristics form Subtropical plantation ecological system.Science in ChinaSeries D:Earth Sciences,2006,36(Supplement):91-102.
[30]Jiang P K,Zhou GM,Xu Q F.Effect of intensive cultivation on the carbon pool of soil inPhyllostachyspraecoxstands.Scientia Silvae Sinicae,2002,(6):6-11
[31]Jiang P K,Xu Q F.Effects of fertilization on soil active organic carbon underPhyllostachyspraecoxstand.Chinese Journal of Applied Ecology,2005,(2):253-256.
[32]Cai R R,HuangF,Sun D,Qin H,Yang F,Zhuang S Y,Zhou G M,Cao Z H.Temporal and spatial variation of soil organic matters inPhyllostachyspraecoxstands with intensive cultivation management.Journal of Zhejiang Forestry College,2007,(4):450-455.
[33]Li G D,Liu G Q,Zhuang S R,Gui R Y.Changes of organic matter in soils planted Lei Bamboo with different years.Chinese Journal of Soil Science,2010,41(4):847-848.
參考文獻:
[3]于貴瑞,孫曉敏,等.中國陸地生態系統碳通量觀測技術及時空變化特征.北京:中國科學出版社,2008.
[4]于貴瑞,孫曉敏.陸地生態系統通量觀測的原理與方法.北京:高等教育出版社,2006:4.
[6]方偉,何鈞潮,盧可學,陳健華.雷竹早產高效栽培技術.浙江林學院學報,1994,11(2):121-128.
[7]汪祖潭,方偉,何鈞潮.雷竹筍用林高產栽培技術.北京:中國林業出版社,1993.
[8]徐繆暢,胡永斌,胡余楚,孫春明,俞茂昌,梁侃,朗進寶.雷竹生產發展與竹園覆蓋技術.上海農業科技,2008,(2):85-86.
[15]宋霞,劉允芬,徐小鋒,于貴瑞,溫學發.紅壤丘陵區人工林冬春時段碳、水、熱通量的觀測與分析.資源科學,2004,26(3):96-104.
[19]周國模.毛竹林生態系統中碳儲量、固定及其分配與分布的研究[D].浙江:浙江大學,2006.
[20]李潔靜,潘根興,李戀卿,張旭輝.紅壤丘陵雙季稻稻田農田生態系統不同施肥下碳匯效應及收益評估.農業環境科學學報,2009,28(12):2520-2525.
[21]李俊,于強,孫曉敏,同小娟,任傳友,王靖,劉恩民,朱治林,于貴瑞.華北平原農田生態系統碳交換及其環境調控機制.中國科學D輯:地球科學,2006,36(增刊):210-223.
[22]劉允芬,宋霞,孫曉敏,溫學法,陳永潤.千煙洲人工針葉林CO2通量季節變化及其環境因子的影響.中國科學D輯:地球科學,2004,34(增刊):109-117.
[23]周國模,姜培坤,徐秋芳.竹林生態系統中碳的固定與轉化.北京:科學出版社,2010.
[24]于貴瑞,王秋鳳,劉迎春,劉穎慧.區域尺度陸地生態系統固碳速率和增匯潛力概念框架及其定量認證科學基礎.地理科學進展,2011,30(7):772-774.
[28]陳述說,李俊,陸佩玲,王迎紅,于強.華北平原麥田土壤呼吸特征.應用生態學報,2004,15(9):1552-1560.
[29]劉允芬,于貴瑞,溫學法,王迎紅,宋霞,李菊,孫曉敏,楊風亭,陳永瑞,劉琪瓃.千煙洲中亞熱帶人工林生態系統CO2通量的季節變異特征.中國科學D輯:地球科學,2006,36(增刊):91-102.
[30]姜培坤,周國模,徐秋芳.雷竹高效栽培措施對土壤碳庫的影響.林業科學,2002,(6):6-11.
[31]姜培坤,徐秋芳.施肥對雷竹林土壤活性有機碳的影響.應用生態學報,2005,(2):253-256.
[32]蔡榮榮,黃芳,孫達,秦華,楊芳,莊舜堯,周國模,曹志洪.集約經營雷竹林土壤有機質的時空變化.浙江林學院學報,2007,(4):450-455.
[33]李國棟,劉國群,莊舜堯,桂仁意.不同種植年限下雷竹林土壤的有機質轉化.土壤通報,2010,41(4):847-848.
