共表達(dá)PRRSV GP5和N蛋白重組桿狀病毒的構(gòu)建與鑒定
2013-11-23 05:57:04漆世華溫文生李玉萍王煥君謝紅玲
中國(guó)獸藥雜志 2013年12期
楊 雷,漆世華,溫文生,李玉萍,王煥君,謝紅玲
(武漢中博生物股份有限公司,武漢430070)
豬繁殖與呼吸綜合征病毒(Porcine Reproductive and Respiratory Syndrome Virus,PRRSV)主要引起妊娠母豬繁殖障礙和仔豬的呼吸道疾病及免疫抑制和持續(xù)性感染,死亡率不斷升高[1-2]。1996年郭寶清等[3]首次從國(guó)內(nèi)疑似豬繁殖與呼吸綜合征(Porcine Reproductive and Respiratory Syndrome,PRRS)感染豬群中分離出PRRSV,從而證實(shí)了本病在我國(guó)的存在。PRRSV是動(dòng)脈炎病毒科、動(dòng)脈炎病毒屬的單股正鏈RNA病毒。其全基因組長(zhǎng)約15 kb,含有9個(gè)開(kāi)放閱讀框(ORFs),0RFla和0RFlb編碼蛋白聚合酶,ORF 2~7分別編碼結(jié)構(gòu)蛋白GP2a、GP2b、GP3、GP4、GP5、M 和 N 蛋白[4]。其中,N蛋白也稱核衣殼蛋白,由0RF7編碼,在病毒粒子中含量較高,N基因高度保守。N蛋白約15 kD,至少有5個(gè)抗原決定簇,其中包括兩型的共同決定簇和特異性決定簇,具有一個(gè)糖基化位點(diǎn),但它不是?糖基化蛋白。N蛋白的C端在維持蛋白構(gòu)象上起重要作用。其N端半段是由Arg、Lys、His 3中堿性氨基酸組成,可能和RNA基因組間的相互作用有關(guān)。N蛋白之間形成同源二聚體,對(duì)病毒的組裝至關(guān)重要,可以誘導(dǎo)機(jī)體產(chǎn)生細(xì)胞免疫作用[5]。GP5蛋白約25 kD,由0RF5編碼,是一個(gè)糖基化蛋白,含有4個(gè)糖基化位點(diǎn),且非常不保守,三次跨膜,并有一個(gè)70個(gè)氨基酸殘基的胞外區(qū)。有6個(gè)抗原決定簇,有證據(jù)表明GP5在PRRSV的抗體識(shí)別過(guò)程中起關(guān)鍵作用,是誘導(dǎo)機(jī)體產(chǎn)生中和抗體的主要結(jié)構(gòu)蛋白,病毒中和作用與抗GP5抗體效價(jià)呈顯著相關(guān)性,同時(shí) GP5還可誘導(dǎo)細(xì)胞凋亡,是公認(rèn)的PRRSV的主要保護(hù)性抗原[6]。
目前用于預(yù)防PRRS的疫苗主要是傳統(tǒng)疫苗——弱毒苗和滅活苗。盡管弱毒疫苗能夠提供較好的免疫保護(hù),但存在毒力返強(qiáng)和散毒的危險(xiǎn)[7]。而滅活疫苗誘導(dǎo)產(chǎn)生的抗體水平很低,且極不穩(wěn)定。因此,研發(fā)確實(shí)、可靠、高效的新一代疫苗也是未來(lái)一個(gè)方向。本研究利用桿狀病毒雙表達(dá)載體(pFastBacTMDual)同時(shí)表達(dá)HP-PRRSV GP5蛋白和N蛋白,以期更好的發(fā)揮PRRSV GP5、N抗原活性及協(xié)同作用,為PRRSV基因工程疫苗的研究奠定基礎(chǔ)。
1 材料與方法
1.1 載體、細(xì)胞、菌株與病毒 桿狀病毒載體pFastBacTMDual、昆蟲(chóng) Sf9 細(xì)胞、E.coli DH10Bac購(gòu)于美國(guó)Invitrogen公司。高致病性豬繁殖與呼吸綜合征病毒廣東分離株(PRRSV GD)由中國(guó)獸醫(yī)藥品監(jiān)察所提供。
1.2 主要試劑 限制性內(nèi)切酶、PCR相關(guān)試劑為TaKaRa公司產(chǎn)品;Cellfectin?ⅡReagent為Invitrogen公司產(chǎn)品;抗PRRSV N蛋白的鼠單克隆抗體為美國(guó)VMRD公司,Prod:P080728-004;抗PRRSV GP5蛋白的兔多抗為英國(guó)ABCAM公司,Prod:RS-4504R;PRRSV GP5抗體檢測(cè)試劑盒為法國(guó)LSI公司,Batch:5-VRTPRA-028;PRRSV N抗體檢測(cè)試劑盒為美國(guó)IDEXX公司,Prod:Z411。
1.3 引物設(shè)計(jì)和PCR擴(kuò)增 參照PRRSV GD株(GenBank登錄號(hào):EU109503.1)ORF7和ORF5基因的序列,各設(shè)計(jì)一對(duì)針對(duì)PRRSV GD毒株N和GP5蛋白基因的特異性引物,擴(kuò)增N蛋白基因帶BamHⅠ酶切位點(diǎn)的上游引物 NF:5’-GGATCCATGCCAAATAACAACGGCAAGC-3’和帶 HindⅢ酶切位點(diǎn)的下游引物 NR:5’-AAGCTTTCATGCTGAGGGTGATGCTGTG-3’;擴(kuò)增 GP5蛋白基因帶XhoⅠ酶切位點(diǎn)的上游引物GP5F:5’-CTCGAGATGTTGGGGAAGTGCTTGACC-3’和帶 KpnⅠ酶切位點(diǎn)的下游引物 GP5R:5’-GGTACCCTAGAGACGACCCCATCGTTC-3’,引物由上海桑尼生物科技有限公司合成。
按照TakaR RNA試劑盒說(shuō)明書(shū)提取PRRSV GD株RNA,以oligo(dT)為引物,以M-MLV進(jìn)行反轉(zhuǎn)錄;然后以cDNA產(chǎn)物為模板,應(yīng)用PCR技術(shù)擴(kuò)增出PRRSV GD毒株的N和GP5全基因;N和GP5基因的擴(kuò)增條件均為:94℃ 5 min;94℃ 40 s,60 ℃ 40 s,72℃ 45 s,35個(gè)循環(huán);72℃延伸10 min。PCR產(chǎn)物進(jìn)行1%的瓊脂糖凝膠電泳。
1.4 重組桿狀Bac-N-GP5病毒的制備 將PCR擴(kuò)增的N蛋白基因和GP5蛋白基因通過(guò)酶切及酶連接的方式插入桿狀病毒載體pFBD(pFast-BacTMDual的縮寫(xiě))中,獲得含有 N蛋白基因和GP5蛋白基因的重組桿狀病毒載體pFBD-NGP5。N蛋白基因通過(guò)polyhedrin啟動(dòng)子調(diào)控,GP5基因由p10啟動(dòng)子直接調(diào)控,其構(gòu)建示意圖見(jiàn)圖1。
按照Invitrogen說(shuō)明書(shū)將上述重組載體pFBDN-GP5通過(guò) Bac-to-Bac系統(tǒng)(Invitrogen,Carlsbad,CA,USA)轉(zhuǎn)入 DH10Bac E.coli,進(jìn)行菌內(nèi)同源重組。用卡那霉素、慶大霉素和四環(huán)素篩選獲得重組病毒DNA,PCR鑒定,然后將正確的重組病毒DNA用Cellfectin?ⅡReagent傳染SF9細(xì)胞,培養(yǎng)3~4 d,獲得重組的桿狀病毒Bac-N-GP5。
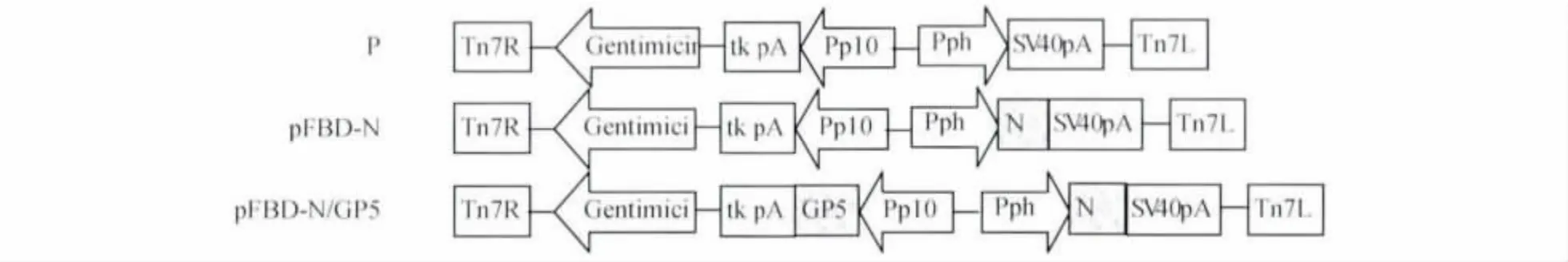
圖1 重組質(zhì)粒構(gòu)建示意圖
1.5 IFA檢測(cè)重組桿狀病毒Bac-N-GP5的表達(dá)
重組桿狀病毒Bac-N-GP5接種Sf9細(xì)胞72 h后,棄去培養(yǎng)基,用PBS洗滌細(xì)胞兩次,80%(V/V)冷丙酮固定細(xì)胞 20 min,PBS洗三次,加鼠抗PRRSV N/兔抗PRRSV GP5抗體 (1∶100)37℃作用1 h,PBS洗三次,F(xiàn)ITC標(biāo)記羊抗鼠二抗/羊抗兔二抗(1∶500)37 ℃作用1 h,PBS 洗三次,50%(V/V)甘油封片,熒光顯微鏡觀察。
1.6 Western blot檢測(cè) 將感染重組桿狀病毒的細(xì)胞裂解液通過(guò)12%SDS-PAGE電泳分離蛋白,采用半干轉(zhuǎn)移(Bio-Rad)轉(zhuǎn)印硝酸纖維膜,并設(shè)細(xì)胞裂解液作為空白對(duì)照。用含5%脫脂乳的PBST封閉過(guò)夜,分別以 N和 GP5蛋白的抗體為一抗,1∶1000稀釋,室溫孵育1 h;辣根過(guò)氧化物酶標(biāo)記的兔抗鼠IgG和羊抗兔IgG為二抗,1∶1000稀釋,室溫孵肓1 h。用DAB顯色液進(jìn)行顯色作用。
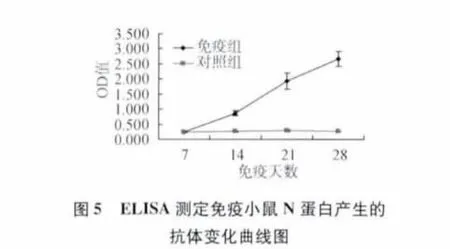
1.7 重組桿狀病毒在小鼠中的免疫原性測(cè)定 用感染重組桿狀病毒的細(xì)胞裂解液以鋁佐劑為佐劑,于0和14 d免疫小鼠,于免疫后7、14、21、28 d采集小鼠血清,用ELISA測(cè)定抗N蛋白和抗GP5蛋白的抗體水平。
2 結(jié)果與分析
2.1 重組桿狀病毒Bac-N-GP5的構(gòu)建與鑒定
通過(guò)RT-PCR擴(kuò)增得到了ORF7和ORF5基因,將其克隆入桿狀病毒表達(dá)載體pFBD中,獲得重組質(zhì)粒pFBD-N-GP5。經(jīng)PCR鑒定表明基因N和GP5克隆到桿狀病毒表達(dá)載體pFBD中(圖2),測(cè)序表明該重組質(zhì)粒的讀碼框正確。
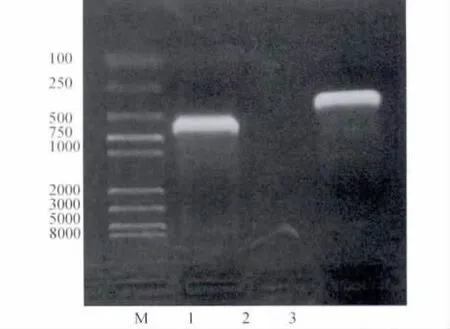
圖2 PCR檢測(cè)重組質(zhì)粒pFBD-N-GP5
2.2 IFA鑒定重組桿狀病毒的表達(dá) 用針對(duì)PRRSVN和GP5蛋白的抗體進(jìn)行IFA試驗(yàn),結(jié)果發(fā)現(xiàn)重組桿狀病毒感染的細(xì)胞出現(xiàn)明顯的熒光,而對(duì)照組則看不到熒光(圖3)。證明N蛋白和GP5蛋白可以在重組桿狀病毒中共同表達(dá)。
2.3 重組桿狀病毒W(wǎng)estern blot分析 重組桿狀病毒感染Sf9細(xì)胞后經(jīng)Western blot檢測(cè),結(jié)果見(jiàn)圖4。抗N蛋白抗體和抗GP5蛋白的抗體分別檢測(cè)到約為15 kD的N蛋白和25 kD的GP5蛋白條帶。證實(shí)N和GP5重組蛋白在Sf9細(xì)胞中表達(dá)成功。
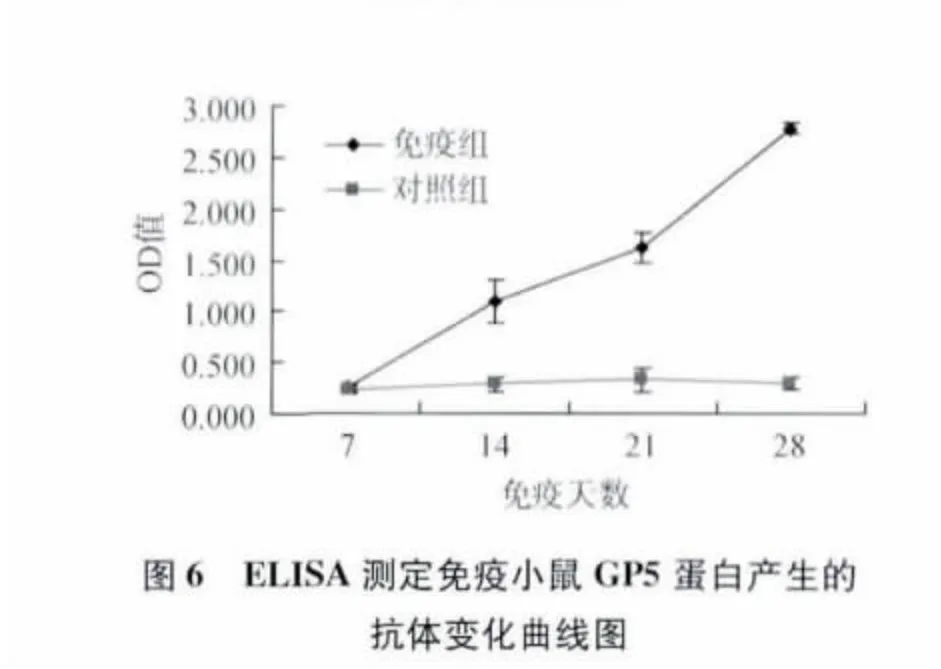
2.4 重組桿狀病毒在小鼠中的免疫原性測(cè)定 用感染重組桿狀病毒的細(xì)胞裂解液以鋁佐劑為佐劑,于0和14 d免疫小鼠,于免疫后7、14、21、28 d采集小鼠血清,分別用抗PRRSV N蛋白和GP5蛋白的抗體ELISA檢測(cè)試劑盒對(duì)免疫后的血清中的N蛋白產(chǎn)生的抗體和GP5蛋白產(chǎn)生的抗體進(jìn)行測(cè)定,結(jié)果表明免疫組血清抗N蛋白和抗GP5蛋白的抗體在14 d開(kāi)始升高,21和28 d的抗體值明顯高于對(duì)照組(圖5、圖6),說(shuō)明重組桿狀病毒表達(dá)的N和GP5蛋白對(duì)小鼠能夠產(chǎn)生很好免疫原性。關(guān)于重組桿狀病毒對(duì)本屬動(dòng)物豬的免疫和攻毒保護(hù)效果還有待于進(jìn)一步研究。

圖3 重組桿狀病毒表達(dá)N和GP5蛋白的間接免疫熒光檢測(cè)

3 討論
目前PRRS的防治主要依靠傳統(tǒng)疫苗,即弱毒疫苗和滅活苗,但由于這些疫苗均存在不同程度的缺陷,主要是免疫效果不理想。此外,活疫苗還存在毒力返強(qiáng)和散毒的危險(xiǎn)等。因此國(guó)內(nèi)外許多研究者都在努力探索各種形式的基因工程疫苗,使用不同的表達(dá)系統(tǒng)來(lái)表達(dá)主要免疫原PRRSV中的GP5、N和M蛋白基因及一些細(xì)胞因子。其中桿狀病毒一昆蟲(chóng)細(xì)胞表達(dá)系統(tǒng)是目前較好的真核表達(dá)系統(tǒng),此系統(tǒng)能夠提高外源蛋白產(chǎn)量,并且表達(dá)的蛋白能夠經(jīng)過(guò)正確的翻譯后修飾加工,更接近天然蛋白的構(gòu)象和活性,因此該系統(tǒng)被應(yīng)用于很多外源蛋白的表達(dá)。
本試驗(yàn)將PRRSV的ORF7基因和ORF5基因分別插入到polyhedrin啟動(dòng)子和p10啟動(dòng)子下,構(gòu)建重組桿狀病毒。結(jié)果表明,用重組病毒Bac-N-GP5感染Sf9細(xì)胞后,經(jīng)IFA和Western blot檢測(cè),能夠在細(xì)胞中檢測(cè)到N和GP5蛋白的共表達(dá)。動(dòng)物免疫試驗(yàn)表明,重組桿狀病毒免疫小鼠后血清中N和GP5蛋白的抗體水平明顯高于對(duì)照組,說(shuō)明重組蛋白能夠引發(fā)小鼠較強(qiáng)的體液免疫反應(yīng)。本試驗(yàn)利用桿狀病毒雙表達(dá)系統(tǒng)同時(shí)在Sf9細(xì)胞中表達(dá)PRRSV N蛋白和GP5蛋白,為下一步應(yīng)用重組桿狀病毒作為PRRSV亞單位疫苗奠定了基礎(chǔ)。
[1]Chung W B,Lin M W,Chang W F,et al.Persistence of Porcine Reproductive and Respiratory Syndrome Virus in Intensive Farrow-to-Finish Pig Herds[J].Can J Vet Res,1997,61(4):292-298.
[2]Van G S,Van R K,Pensaert M.Interaction between Porcine Reproductive Respiratory syndrome Virus and Bacterial Endotoxin in the Lungs of Pigs:Potentiation of Cytokine Production and Respiratory Disease[J].J Clin Microbiol,2003,4l(3):960-966.
[3]郭寶清,陳章水,劉文興,等.從疑似PRRS流產(chǎn)胎兒分離豬繁殖與呼吸綜合征病毒(PRRSV)的研究[J].中國(guó)畜禽傳染病,1996,87(2):1-5.
[4]Nelsen C J,Murtaugh M P,F(xiàn)aaberg K S.Porcine reproductive and respiratory syndrome virus comparison:divergent evolution on two continents[J].J Virol,1999,73(1):270-280.
[5]Nelson E A,Christopher-Hennings J,Drew T,et al.Differentiation of U.S.and European isolates of porcine reproductive and respiratory syndrome virus by monoclonal antibodies[J].J Clin Microbiol,1993,31:3184-3189.
[6]Zhou Y J,Yu H,Tian Z J,et al.Monoclonal antibodies and conserved antigenic epitopes in the C terminus GP5 protein of the North American type porcine reproductive and respiratory syndrome virus[J].Vet Microbiol,2009,138:1-10.
[7]Madsen K G,Hansen C M,Madsen E S,et al.Sequence analysis of porcine reproductive and respiratory syndrome virus of the American type collected from Danish swine herds[J].Arch Virol,1998,143(9):1683-1700.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
