轉基因抗蟲水稻的研發與應用及在我國的發展策略
2013-11-04 06:58:40徐秀秀韓蘭芝彭于發侯茂林
環境昆蟲學報 2013年2期
關鍵詞:水稻
徐秀秀,韓蘭芝,彭于發,侯茂林
(中國農業科學院植物保護研究所/植物病蟲害生物學國家重點實驗室 北京 100193)
水稻是世界上最重要的糧食作物之一,全世界有30 多億人口以稻米為食。2008年全球水稻的種植面積已超過1.59 億公頃(http://faostat.fao.org/site/567/default.aspx),其中我國水稻種植面積已超過全球種植面積的18%。水稻也是蟲害最多的糧食作物之一,田間害蟲種類達624 余種,每年由稻螟所造成的經濟損失達115 億元(盛承發等,2003),由稻飛虱所造成的產量損失達150萬噸(黃世文,2010)。特別是近年來,隨水稻耕作制度的變更、品種更新、抗藥性產生及氣候變暖等因素,水稻害蟲在全國范圍內的發生數量和危害程度明顯上升,給我國的水稻生產造成嚴重威脅。傳統化學防治方法的長期使用,不但增加生產成本,造成環境污染,而且還破壞了生態平衡,因此選育抗蟲轉基因水稻是防治害蟲最經濟、有效的方法。
隨全球生物技術的發展,利用基因重組技術將外源抗蟲基因導入水稻,使水稻自身產生抗蟲蛋白從而達到防治害蟲的目的已成為現實。自1988年,第一例Bt 水稻成功轉化后,眾多研究者相繼將不同類型的Bt 殺蟲基因、蛋白酶抑制劑基因和植物或動物源凝集素基因轉入不同水稻品種,獲得一系列對靶標害蟲控制效果顯著的轉單、雙或多基因抗蟲品系,為水稻害蟲的防治提供了有效途徑(Bajaj and Mohanty,2005;Deka and Barthakur,2010)。至今,全球已研發了近50個轉基因抗蟲水稻品系,隨研發技術的進一步深入,新型抗蟲、多基因、復合性狀的轉基因水稻新品種將不斷涌現。
本文對全球轉基因抗蟲水稻的研發與應用現狀進行了系統綜述,分析了目前我國轉基因抗蟲水稻發展所存在的問題及面臨的機遇,并結合我國水稻生產的實際情況,提出了我國轉基因抗蟲水稻的發展策略,以期為抗蟲水稻的研發與應用、環境安全評價及風險管理提供科學依據。
1 轉基因抗蟲水稻的研發進展
由于水稻自身缺乏高效的抗蟲基因,轉基因抗蟲水稻轉化所需的目的基因主要來自外源基因。其中來自于蘇云金芽孢桿菌(Bacillus thuringiensis,Bt)的Bt 基因是目前世界上應用最廣和最高效的抗蟲基因,它對鱗翅目(Cohen et al.,2000)、雙翅目(Andrews et al.,1987)和鞘翅目害蟲(Krieg et al.,1983;Herrnstadt et al.,1986)均具有較高的抗性,且對人、動物和生態環境安全(表1)。除Bt 基因外,某些蛋白酶抑制劑、植物凝集素、核糖體失活蛋白、植物次生代謝物基因等對昆蟲也具有較好的抗蟲效果,已被廣泛應用在轉基因抗蟲水稻的研發上(表2)。
1.1 Bt 水稻
1981年第一個Bt 殺蟲基因被克隆;1993年第一個Bt 水稻品系研制成功;2000年起,Bt 水稻開始陸續進入田間試驗階段(Tu et al.,2000;Anonymous,2001;High et al.,2004)。研發前期,Bt 水稻主要以轉單個cry1A 基因為主,如cry1Ab、cry1Ac和cry1Ab/cry1Ac 融合基因等(陳浩等,2009)。在我國培育較為成功的為華中農業大學研發的“華恢1 號”、“Bt 汕優63”(轉cry1Ab/cry1Ac 基因)和浙江大學的“克螟稻”(轉cry1Ab基因),它們在整個生育期內對二化螟、三化螟和稻縱卷葉螟均表現較高的抗性(舒慶堯等,1998;Shu et al.,2000;Tu et al.,2000;Ye et al.,2001,2003)(表1)。印度也已培育出多個抗三化螟的轉基因水稻品種,但未看到其田間試驗的后續報道(Nayak et al.,1997;Khanna and Raina,2002;Ramesh et al.,2004)(表1)。巴基斯坦和地中海沿岸的一些國家也開展了相關的Bt 水稻研究,并獲得一些抗蟲效果優良的Bt 水稻品系(Breitler et al.,2000,2004;Marfa et al.,2002;Husnain et al.,2002;Bashir et al.,2004)(表1)。為延緩害蟲抗性發展,培育轉雙或多基因Bt 水稻是未來發展的必然趨勢。其他類別的Bt 基因如cry2A、cry1C和cry9C 與cry1A 共同氨基酸的同源性較低(<45%),不易產生交互抗性,適宜與cry1A 組合研制轉雙或多基因抗蟲水稻(Alcantara et al.,2004)。有研究用分別轉cry1Ab、cry1Ac、cry1C和cry2A 4個單基因的抗蟲明恢63 株系作親本,以5種組合方式分別正反交,將不同的Bt 基因兩兩聚合在一起,獲得10種雙價Bt 株系。室內抗蟲鑒定結果表明,大部分雙價Bt 基因組合有協同增效作用,其抗蟲性明顯高于單基因Bt 水稻(陳浩等,2009)。
隨著水稻轉基因技術的成熟,人們不再滿足僅僅將外源基因導入到水稻基因組中。根據不同需要,一些特殊的轉基因技術逐漸發展起來。出于延緩害蟲抗性和食品安全性的考慮,人們希望Bt 毒蛋白僅僅在水稻易受蟲害攻擊的莖、葉部位表達,在此基礎上發展了組織特異性或誘導性表達外源基因的方法。Ye et al.(2009)采用根癌農桿菌介導將綠色組織特異性表達的rbcS 啟動子驅動下的cry1C 基因導入粳稻品種中花11,獲得高效抗蟲且Bt 毒蛋白僅在綠色組織中高效表達的轉基因株系,該株系葉片中Bt 毒蛋白的表達量是使用玉米Ubiquitin 組成性啟動子的3 倍,而在胚乳中Bt 毒蛋白的表達量不到Ubiquitin 啟動子的千分之一(Tang et al.,2006;Ye et al.,2009)。利用傷口誘導型啟動子促進抗蟲基因在轉基因植株中受傷害的部位和時間段內表達是減少生物量損失和延緩靶標害蟲抗性產生的另一有效途徑(De Maagd et al.,1999;Sharma et al.,2004),玉米蛋白酶抑制基因的-689/+197 區域可為Bt 水稻中cry1B 基因提供高效愈傷誘導性表達,該轉基因水稻在田間高抗二化螟(Breitler et al.,2001)。

表1 全球已研發的Bt 水稻品系Table 1 The development of transgenic insect-resistant rice lines in the world

文獻來源:http://www.cera-gmc.org/index.php?action=gm_crop_database;http://www.isaaa.org/gmapprovaldatabase/default.asp;High et al.,2004;Bajaj and Mohanty,2005;Deka and Barthakur,2010.“SSB”表示二化螟;“YSB”表示三化螟;“RLF”表示稻縱卷葉螟;“BPH”表示褐飛虱,“LH”表示葉蟬,同下表2。Source from:http://www.cera-gmc.org/index.php?action=gm_crop_database;http://www.isaaa.org/gmapprovaldatabase/default.asp;High et al.,2004;Bajaj and Mohanty,2005;Deka and Barthakur,2010.Abbreviations:SSB,striped stem borer;YSB,yellow stem borer;RLF,rice leaffolder.LH,leaf hopper.It is the same to the following table 2.
1.2 非Bt 抗蟲水稻
雖然Bt 基因一直是轉基因植物應用最成功、最廣泛的抗蟲基因,但是將其與其它的抗蟲基因組合使用才能達到持久抗蟲的目的,我國的轉cry1Ac+CpTI 基因水稻及印度小規模試種的轉cry1Ab+Xa21+GNA 基因聚合的抗蟲水稻均是比較成功的實例(Bentur,2006;Han et al.,2006,2007)。有些植物本身就含有一些非Bt 的抗蟲基因,如蛋白酶抑制劑(Protease inhibitors)和植物凝集素(Plant lectins)等(Sharma et al.,2004)。蛋白酶抑制劑廣泛分布于植物組織和器官中,能削弱或阻斷蛋白酶對食物中蛋白質的消化,導致昆蟲非正常死亡。如轉蠶豆蛋白酶抑制劑(Mochizuki et al.,1999)、豇豆蛋白酶抑制劑(Xu et al.,1996)、大豆蛋白酶抑制劑、土豆蛋白酶抑制劑(PinⅡ)(Sharma et al.,2004)和抗象鼻蟲的大麥蛋白酶抑制劑(Alfonso-Rubi et al.,2003)基因水稻均已成功研制(表2)。其中轉豇豆胰蛋白酶抑制劑基因(CpTI)水稻在中國研究的最早且最為成熟,相關的環境安全性評價已完成,正在申請生物安全證書(http://www.stee.agri.gov.cn/biosafety/spxx/)。植物凝集素是一類能特異識別并可逆結合糖類復合物的非免疫性球蛋白,當被昆蟲吞食至消化道時,與昆蟲腸道周圍細胞壁膜糖蛋白結合,影響營養物質的吸收,并在消化道內誘發病變,使昆蟲致病而死亡。目前在基因工程方面應用最多的有雪花蓮凝集素(GNA)、豌豆凝集素(P-lec)和大蒜凝集素(ASAL)等。轉雪花蓮凝集素基因水稻對小褐飛虱和褐飛虱表現出明顯的抗性,既能降低其存活率又能降低其產卵能力并延遲發育(Nagadhara et al.,2003;Tang et al.,2001)。轉大蒜凝集素基因水稻IR64對跳蟲類、褐飛虱和綠葉蟬均具有較好的控制效果(Saha et al.,2006)(表2)。洋蔥凝集素(Allium Cepa agglutinin,ACA)對刺吸類害蟲具有較好的殺蟲效果(Hossain,2006)。
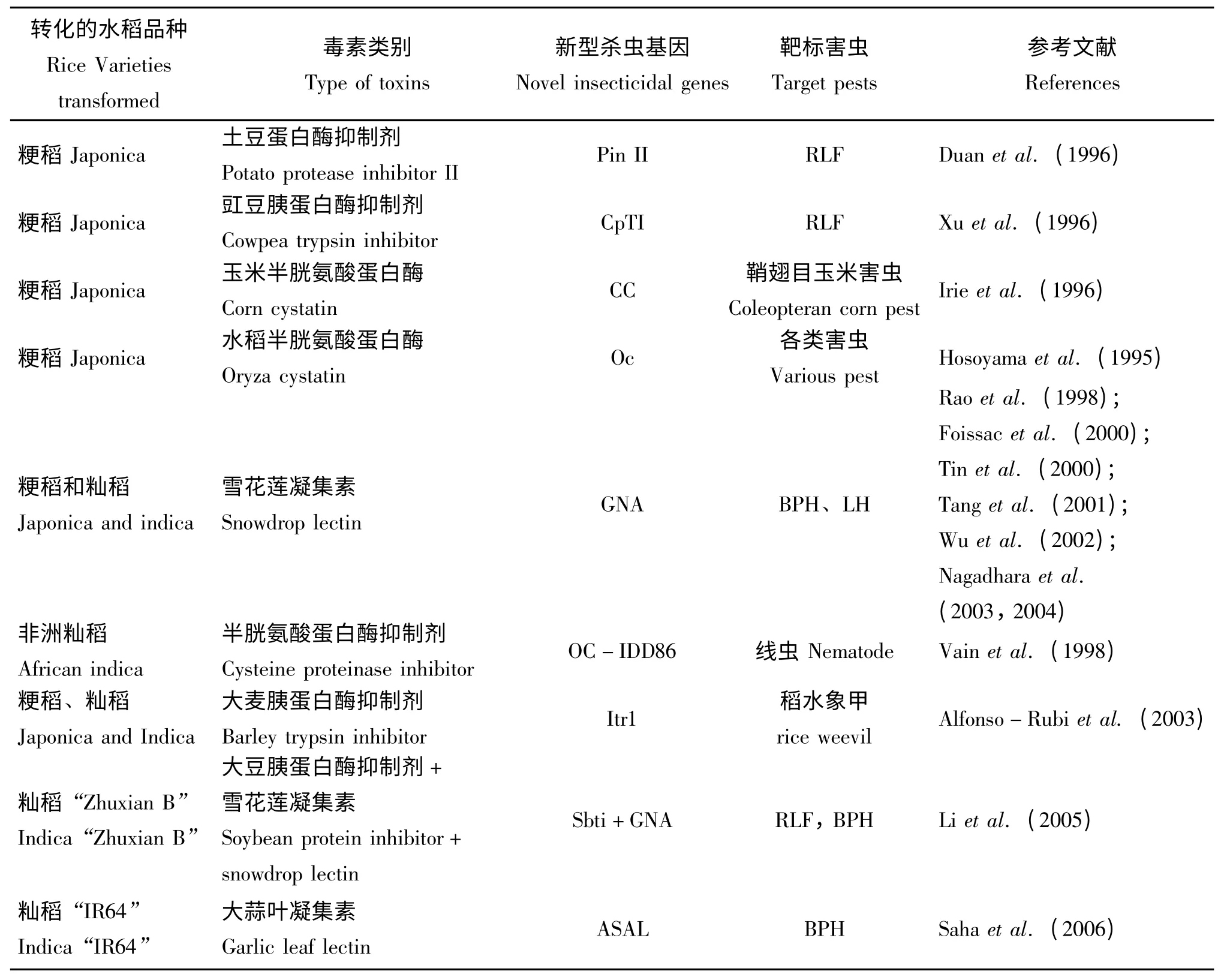
表2 全球用于抗蟲水稻品系研發的非Bt 基因Table 2 Insect resistance genes other than Bt used in development of transgenic rice lines in the world
2 轉基因抗蟲水稻的應用
盡管國內外正積極對一些抗蟲效果優異的轉基因抗蟲水稻進行中間試驗、環境釋放和生產性試驗。但目前還沒有一個國家的抗蟲水稻處于商業化應用階段。這主要因為人們對糧食問題過于敏感,轉基因抗蟲水稻的食品安全性還尚未定論,各國對轉基因水稻的產業化持謹慎態度。伊朗為種子增殖而在2005年批準種植了4000 公頃Bt 水稻(James,2009),但目前伊朗政府仍不允許進行商業化生產。中國于2009年8月17 日批準了兩個Bt 水稻品種華恢1 號及其雜交種Bt 汕優63 在湖北省生產應用的安全證書,但未進行商業化種植,這說明中國有望成為世界上第一個使用轉基因抗蟲水稻的國家(http://www.stee.agri.gov.cn/biosafety/spxx/)。此外,農業部轉基因生物安全委員會于2009年5月還批準克螟稻、T1c-9(轉cry1C 基因),T2A-1(轉cry2A 基因)、科豐6和科豐8 號五種轉基因水稻通過生產性試驗,等待生物安全證書的發放(http://www.stee.agri.gov.cn/biosafety/spxx/)。
除抗蟲水稻外,一些藥用或耐除草劑水稻也陸續被批準進口或商業化應用。1998年起,美國批準Ventra Bioscience 公司研發的轉溶菌酶、乳鐵蛋白、人血清白蛋白基因的3個藥用轉基因水稻的商業化種植,種植面積由最初的400 畝增加到2007年的3 萬畝,2006~2007年種植區域增加到堪薩斯州和北卡羅來納州。1999年,美國批準了安萬特公司的轉bar 基因耐除草劑水稻LL RICE 06和LL RICE 62 的商業化種植,2000年批準該水稻可以食用。2006年8月,美國又批準了轉bar 基因耐除草劑水稻LL RICE 601 的商業化種植(http://www.isaaa.org/gmapprovaldatabase/default.asp)。加拿大、墨西哥、澳大利亞分別于2006年、2007年、2008年批準了耐除草劑轉基因水稻LL RICE 62 的進口,并允許食用(http://www.isaaa.org/gmapprovaldatabase/default.asp)。
雖然轉基因抗蟲水稻尚未得到商業化種植,但其研發勢頭強勁。以中國為例,從1998年至今農業轉基因生物安全管理辦公室已批準2375份田間試驗的申請,包括1598份中間試驗,503份環境釋放及274份生產性試驗申請(http://www.stee.agri.gov.cn/biosafety/spxx/)。這 說 明,轉基因抗蟲水稻的發展勢頭良好,產業化前景廣闊。
3 我國轉基因抗蟲水稻面臨的機遇與挑戰
3.1 我國轉基因抗蟲水稻發展面臨的機遇
目前,我國水稻育種面臨的問題十分突出,主要表現在水稻品種抗性較差,害蟲危害日益嚴重,由于水稻及其近緣種中尚未發現對螟蟲有效的抗性基因,無法通過常規育種加以改良,必須通過轉基因技術引入外源抗性基因;對于褐飛虱,雖然在水稻中存在相應的抗性基因資源,但由于抗性基因大多為數量性狀,單個基因的效應微小,難以通過常規育種加以改良,必須依賴分子標記聚合育種或轉基因育種技術加以解決。而且我國是世界上最大的水稻生產地,約4 億農戶以種植水稻為生。據統計,Bt 水稻的種植能使產量提高8%,殺蟲劑使用量減少80%,每年可為我國帶來約40 億美元的收益,具有較大的經濟、生態和社會效益(Huang et al.,2005)。可見,我國水稻生產對轉基因抗蟲水稻具有迫切的需求。
同時,轉基因抗蟲育種研究也是我國農業發展的一項既定國策。80年代開始,我國一貫并高度重視轉基因技術的研究和應用,2004年以來連續7個中央一號文件都強調加快農業生物技術的發展;863、973 等國家科技計劃都將轉基因技術研究作為重大項目予以支持;2008年國家啟動實施“轉基因生物新品種培育重大專項”;2009年生物育種又被列入“戰略性新興產業規劃”,這均為我國轉基因抗蟲水稻的發展提供了良好的鍥機(http://world.people.com.cn/GB/41217/11975156.html)。
在國家政策扶植和重大項目的資助下,我國在功能基因克隆、水稻遺傳轉化、抗蟲品系培育等領域均獲得較大進展,為轉基因抗蟲水稻的研發提供了良好的技術儲備。截至2011年4月,全球已有701個Bt 殺蟲基因被克隆并命名,這些基因來自30 多個國家和地區,其中我國最多,有259個(張杰等,2011)。通過自主研發,我國已先后從水稻上克隆了10 多個有重要利用價值的農藝性狀基因用于水稻轉化。截至2006年8月,我國受理的與轉基因水稻相關的專利75 例,其中涉及功能基因克隆的有74 項(黃德林,2007)。在基因操作方面,已建立了轉化大DNA 片段和自動裝載外源基因序列的轉化載體技術,大大提高了載體構建的工作效率。在水稻遺傳轉化技術方面,已建立了高效、穩定、規模化的農桿菌介導轉化技術。抗蟲品系培育方面,我國被公認處于世界領先地位,已獲得“華恢1 號”、“Bt 汕優63”、“克螟稻”和“科豐”系列等抗蟲效果優異的轉基因水稻品系(朱禎,2010)。
3.2 我國轉基因抗蟲水稻研發存在的主要問題
雖然我國在轉基因水稻方面的研究與國際基本同步,在發展中國家居領先地位,但不可否認,與國際領先水平相比,我們仍存在較大差距,主要表現在:(1)抗蟲基因品種單一、缺乏有重要利用價值的自主知識產權基因。目前,我國抗蟲水稻以轉cry1A 單價基因為主,抗蟲譜窄,害蟲極易產生抗性。而cry1A 基因對螟蟲高抗,但對稻飛虱、葉蟬等刺吸類害蟲無控制作用。雖然我國已獲得一些殺蟲效果較好并具有自主知識產權的新型Bt 基因,如cry1Ah、cry1C和Cry2A 等(張杰等,2011),但大多數仍停留在同源序列擴增和候選基因獲得水平,特別是還未發現對稻飛虱效果較好的內源或外源基因,這嚴重制約了抗蟲水稻的研發與應用。(2)缺乏高效、穩定、安全、規模化的轉基因育種技術。我國目前雖已初步建立了安全、高效的轉基因技術體系,但規模小而分散,主要停留在實驗室水平,與國外跨國公司大規模轉化水平相比,存在巨大差距;組織特異性或葉綠體轉化等新型、高效轉化技術進展緩慢,多基因聚合轉化僅有初步研究;有效的配套育種技術體系還未建立,轉基因生物新品種培育效率偏低。(3)研究環節中的上、中、下游脫節,產業化能力薄弱。我國轉基因抗蟲水稻研發主要集中在高校和國家級科研機構,品種的培育和推廣則集中在地方農業科研院所,他們之間缺乏有機聯系。這種研究機制使上、中、下游脫節,優勢得不到有效互補,難以對我國轉基因研究成果進行系統整合,致使研究成果的轉化和產業化過程受阻,國際競爭力不足。(4)安全性評價研究相對滯后,檢測和監測水平有待提高。雖然我國在轉基因抗蟲水稻的環境和食品安全性評價中已做了大量工作,建立了相應的技術評價標準,但考慮到水稻的主糧地位,與其他抗蟲作物相比,我們對轉基因抗蟲水稻的安全性評價更應該采取嚴格和謹慎的態度。(5)公眾對轉基因技術及產品認知度較低,科普宣傳有待提高。
4 我國水稻生產特點及對轉基因抗蟲水稻種植的要求
目前,種植轉基因作物最多的國家是美國、巴西、阿根廷和加拿大。上述國家的農業為集約化和規模化的種植模式,作物一般實行一年一熟免耕種植,這種種植模式有利于轉基因作物的布局和監管(李云河等,2011)。而我國基本上是一家一戶式的微型家庭農場種植模式,且推行多熟制增加復種指數和采用間作套種的模式來提高土地利用率、促進作物高產、高效。這種種植模式決定了我國在轉基因作物種植技術上要求更加嚴格。從害蟲抗性治理的角度來說,高劑量/庇護所策略在西方集約化和規模化的農業生產中易于推行和使用,但在以小農經濟種植經營為主的中國很難實行。因此,對于高度雜食性害蟲的抗性治理,利用天然庇護所將是最佳選擇,我國轉Bt 基因棉花抗性成功治理的經驗也說明了這一點(Wu et al.,2002)。但水稻害蟲稻螟和稻飛虱均為寡食性害蟲,可供利用的天然庇護所非常少。因此,考慮到當前庇護所實施的困難性,通過合理的作物布局來建立適合我國國情的庇護所策略是非常必要的,如轉基因抗蟲水稻與抗除草劑或其他抗性性狀的轉基因水稻鄰作或間作,非抗蟲性狀水稻將作為抗蟲水稻的天然庇護所來延緩害蟲抗性發展。
5 轉基因抗蟲水稻在我國的發展策略
針對全球轉基因抗蟲水稻的研發、應用現狀和我國抗蟲水稻研究中存在的問題和面臨的機遇,并結合我國水稻自身生產特點,提出了目前我國在轉基因抗蟲水稻研發、安全評價及政府監管方面的發展策略。
5.1 我國轉基因抗蟲水稻的研發策略
目前,我國轉基因水稻育種的戰略部署是以“轉基因生物新品種培育重大專項”為主導,專項的發展目標為:近期以發展并改良單一性狀的抗蟲、抗病和抗除草劑轉基因水稻為主;中期以發展兩個或兩個以上的抗性性狀轉基因水稻為主;遠期以發展多基因、復合性狀的轉基因抗蟲水稻為重點。鱗翅目害蟲二化螟、三化螟、大螟和稻縱卷葉螟一直為水稻上的重要害蟲,年均發生面積高達6.1 億畝次,且近幾年呈逐年遞增趨勢(黃世文,2010)。為有效治理螟蟲為害,我國科學家已先后培育出“華恢1 號”、“Bt 汕優63”等多個抗蟲效果優良的轉基因抗蟲水稻品系,并由農業部于2009年8月為其頒發了在湖北省生產應用的安全證書;中國科學院遺傳與發育生物學研究所與福建農業科學院研制的“科豐”系列雙價抗蟲水稻和浙江大學研制的“克螟稻”對鱗翅目害蟲也具有較好的控制效果,相關的生物安全性評價工作已經完成,正在申請安全證書,這說明高抗螟蟲的Bt 水稻具有雄厚的產業化基礎,應作為重點、優先培育對象。
稻飛虱及葉蟬類等同翅目害蟲的發生地位與稻螟基本相同,稻飛虱大多為遷飛性害蟲,可遠距離遷飛危害,難以預測預防,經常會突發成災。如果抗鱗翅目昆蟲轉基因水稻得到推廣和應用,殺蟲劑的使用勢必減少,可能會造成稻飛虱和葉蟬種群的猖獗危害。因此,抗稻飛虱品系的培育是保障抗螟蟲轉基因水稻可持續應用的關鍵,也是繼螟蟲之后抗蟲水稻重點、優先培育的領域。雖然目前報道已定位的水稻抗褐飛虱基因共27個,鑒定的與抗性相關的重要QTL 有50 多個(楊震等,2011),但只有Bph14和Bph18 被克隆,且大多數抗性基因的定位精度較低,影響了目的基因的轉化效率。而且褐飛虱有多個生物型,極易發生變異,使抗性品種失去作用,所以我國抗稻飛虱轉基因水稻的研發仍面臨巨大挑戰。
為開辟新的育種途徑,拓展轉基因作物的功能,提高資源利用率和當前市場的需求,發展植保、功能和專用特性相結合的多基因、復合性狀的轉基因水稻是未來發展的必然趨勢。2011年,12個國家種植了復合兩種或三種性狀的轉基因作物,占全球轉基因作物種植面積的26%(James,2011)。美國種植的轉基因作物中有41%同時具備抗蟲和耐除草劑性狀,轉入八個基因的玉米SmartstaxTM同時擁有多種抗蟲和耐除草劑性狀(James,2010)。從害蟲抗性管理的角度來看,研究具有不同抗蟲性狀的轉基因作物對延緩害蟲抗性具有重要作用。在多基因抗蟲水稻中,每個基因的作用方式不同,故抗性治理只需較小的庇護所,且應用時間更持久。除轉cry1Ac+CpTI和轉GNA+SbTI 基因水稻研究較為成熟外,其它多基因抗蟲水稻如cry1Ab+cry1C,cry1Ab+cry2A,cry1Ac+cry2A,cry2A+cry1C和cry1Ab+vip3H+G6-epsps 仍在研究階段(Chen et al.,2011)。不可否認,它們的成功研制將極大地促進了我國轉基因抗蟲水稻的產業化發展。
與此同時,作為產業儲備項目,我們還必須加強對新的Bt 及植物源抗蟲基因的挖掘和利用研究,以防抗性種質資源過分單一而出現抗蟲性喪失的問題。一般認為,微效多基因的利用有助于保持水稻對稻飛虱的持久抗性。因此,育種工作者可充分利用含抗性基因的野生稻‘突變體資源'加強對稻飛虱內源新抗性基因的發掘與利用(楊震等,2011)。除進行內源基因發掘外,一些抗蟲效果優良的外源基因的利用也是非常必要的。在轉基因操作技術方面,我國應建立和完善高效的基因表達調控技術;發掘適于不同轉基因需求的啟動子元件;構建具有自主知識產權和應用價值的新型轉化載體,提高外源基因表達水平;開發適合農桿菌介導法的多基因聚合轉化技術,建立規模化轉基因體系;將轉基因育種與常規育種、分子標記輔助育種等技術緊密結合,大規模創建遺傳穩定性好的轉基因新材料,培育作物新品種。
5.2 我國轉基因抗蟲水稻安全性評價的發展策略
轉基因抗蟲水稻最受關注的生態風險是害蟲抗性,它決定著抗蟲水稻的使用壽命。因此,在產業化應用之前,靶標害蟲對外源殺蟲基因的抗性風險評價應貫穿整個研發過程。在室內建立抗性汰選品系,通過繼代汰選、抗性遺傳力測定、抗性預測系統評價害蟲的抗性風險;在轉基因水稻環境釋放地,長期監測靶標害蟲抗性基因頻率的變化,建立抗性風險評估模型。產業化應用之后,應建立靶標害蟲早期抗性的監測與預警技術。同時,針對我國水稻生產的實際情況和靶標害蟲的取食特點,建立相應的害蟲抗性管理策略,具體措施包括:(1)針對水稻產區害蟲的發生特點制定科學的生產種植和病蟲害綜合防治策略;(2)建立健全抗蟲水稻商業化監測體系,保障農民種植高水平、穩定表達殺蟲蛋白的抗蟲水稻,根據害蟲種群結構變化動態和靶標害蟲抗性演化趨勢,調整害蟲綜合防治措施;(3)通過合理的作物布局和頂層設計,將非抗蟲性狀水稻作為抗蟲水稻的庇護所,延緩害蟲抗性發展;(4)研發并應用具有不同作用機制的雙價和多價抗蟲水稻,增強抗蟲效率和擴大殺蟲譜,同時并作為替代產品適時推出(彭于發等,2011)
多基因、復合性狀轉基因抗蟲新品種的出現要求建立與之相同步的環境安全評價技術。與單個抗蟲性狀不同的是,復合性狀抗蟲作物涉及多個轉基因,基因之間可能存在非關聯、關聯、代謝等相互作用關系,可能引發協同效應。因此,復合性狀轉基因植物的安全性評價應考慮轉基因性狀間的關聯和互作。目前,我國現有的法規體系主要針對單性狀轉基因作物,對于復合性狀還沒有統一的安全評價原則和方法,這勢必影響我國轉基因作物的產業化應用。隨轉基因技術的進一步發展,新功能、新用途、多基因復合性狀抗蟲水稻新品種將不斷涌現,建立適合我國國情的轉基因水稻環境安全評價體系將是一項非常重要而迫切的任務。
5.3 政府層面的監管、決策與宣傳
政府是社會行為的主體,它的參與、協調和引導將對我國抗蟲作物的產業化應用具有重要推動作用。為有效延緩害蟲抗性發展,政府在品種審批和作物布局方面應統籌規劃和頂層設計,嚴禁批復具有相同作用機制的抗蟲水稻的商業化應用,積極推動多基因、復合性狀抗蟲水稻的研發和應用;避免同一抗蟲基因/機制轉基因水稻的鄰作或連作。對水稻種子市場進行嚴格監管,嚴禁殺蟲蛋白低劑量表達的抗蟲水稻種子進入市場。
由于我國研發過程中的上、中、下游脫節,優勢得不到有效互補,難以對轉基因研究成果進行系統整合及轉化,產業化過程受阻。建議從政府層面,進行引導和協調,鼓勵大型企業早期介入,逐步成為轉基因作物研發、投入和開發的主體,提高我國轉基因技術產業的國際競爭力。
為消除公眾對轉基因技術和產品的疑慮和恐慌,增進其對國家生物技術發展戰略的了解和對轉基因技術的認知,政府應積極擴大與公眾間的信息交流,牽頭組織并開展形式多樣的科普宣傳活動,為公眾解疑釋惑,引導公眾科學、理性地對待轉基因技術,積極創造有利轉基因技術發展的輿論氛圍。
References)
Alcantara EP,Aguda RM,Curtiss A,Dean DH,Cohen MB,2004.Bacillus thuringiensis delta endotoxin binding to brush border membrane vesicles of rice stem borers.Arch.Insect Biochem.Physiol.,55(4):169-177.
Alfonso-Rubi J,Ortego F,Castanera P,Carbonero P,Diaz L,2003.Transgenic expression of trypsin inhibitor CMe from barley in indica and japonica rice,confers resistance to the rice weevil Sitophilus oryzae.Transgenic Res.,12:23-31.
Andrews RW,Fausr R,Wabiko MH,Roymond KC,Bulla LA,1987.Biotechnology of Bt:a critical review.Bio/Technology,6:163-232.
Anonymous,2001.Consensus statement on sustainable use of transgenic rice for pest management.Mol.Breed,7:99-100.
Bajaj S,Mohanty A,2005.Recent advances in rice biotechnology-towards genetically superior transgenic rice.Plant Biotechnol.J.,3(3):275-307.
Bashir K,Husnain T,Fatima T,Latif Z,Mehdi SA,Riazuddin S,2004.Field evaluation and risk assessment of transgenic indica basmati rice.Mol.Breed.,13(4):301-312.
Bentur JS,2006.Host plant resistance to insects as a core of rice IPM.Science,Technology and Trade for Peace and Prosperity(IRRI,ICAR).McMillan India Ltd,419-435.
Breitler JC,Cordero MJ,Royer M,Meynard D,San Segundo B,Guiderdoni E,2001.The-689/+197 region of the maize protease inhibitor gene directs high level,wound-inducible expression of the cry1B gene,which protects transgenic rice plants from stem borer attack.Mol.Breed.,7:259-274.
Breitler JC,Marfa V,Royer M,Meynard D,Vassal JM,Vercambre B,Frutos R,Messeguer J,Gabarra R,Guiderdoni E,2000.Expression of a Bacillus thuringiensis cry1B synthetic gene protects Mediterranean rice against the striped stem borer.Plant Cell Rep.,19:1195-1202.
Breitler JC,Vassal JN,del Mar Catala M,Meynard D,Marfa V,Mele E,Royer M,Murillo L,San Sequndo B,Guiderdoni E,Messequer J,2004.Bt rice harbouring cry genes controlled by a constitutive or wound-inducible promoter,protection and transgene expression under Mediterranean field conditions.Plant Biotechnol.J.,2(5):417-30.
Chen H,Lin YJ,Zhang QF,2009.Review and prospect of researches on transgenic rice.Chin.Sci.Bull.,54(18):2699-2717.[陳浩,林擁軍,張啟發,2009.轉基因水稻研究的回顧與展望.科學通報,54(18):2699-2717]
Chen M,Shelton A,Ye GY,2011.Insect-resistant genetically modified rice in China:from research to commercialization.Ann.Rev.Entomol.,56:81-101.
Cohen MB,Gould F,Bentur JS,2000.Bt rice:practical steps to sustainable use.Int.Rice Res.Notes,25:5-10.
De Maagd,RA,Bosch D,Stiekema W,1999.Bacillus thuringiensis toxin-mediated insect resistance in plants.Trends Plant Sci.,4:9-13.
Deka S,Barthakur S,2010.Overview on current status of biotechnological interventions on yellow stem borer Scirpophaga incertulas(Lepidoptera:Crambidae)resistance in rice.Biotechnol.Adv.,28:70-81.
Han LZ,Wu KM,Peng YF,Wang F,Guo YY,2006.Evaluation of transgenic rice expressing Cry1Ac and CpTI against Chilo suppressalis and intrapopulation variation in susceptibility to Cry1Ac.Environ.Entomol.,35:1453-1459.
Han LZ,Wu KM,Peng YF,Wang F,Guo YY,2007.Efficacy of transgenic rice expressing Cry1Ac and CpTI against the rice leaffolder,Cnaphalocrocis medinalis.J.Invertebr.Pathol.,96:71-79.
Herrnstadt C,Soares GG,Wilcox ER,Edwards DL,1986.A new strain of Bacillus thuringiensis with activity against coleopteran insects.Bio/Technology,4:305-308.
High SM,Cohen MB,Shu QY,Altosaar I,2004.Achieving successful deployment of Bt rice.Trends Plant Sci.,9:286-292.
Hossain MA,Maiti MK,Basu A,Sen S,Ghosh AK,Sen SK,2006.Transgenic expression of onion leaf lectin gene in Indian mustard offers protection against aphid colonization.Crop Sci.,46:2022-2032.
Huang DL,2007.Researches on Patent Strategy of Transgenic Rice.Beijing:Chinese Agricultural Press,70-177.[黃德林,2007.轉基因水稻專利戰略研究.北京:中國農業出版社,70-177]
Huang JK,Hu RF,Rozelle S,Pray C,2005.Insect-resistance GM rice in farmers' fields:assessing productivity and health effects in China.Science,308:689-690.
Huang SW,2010.Key Technical Analysis of Main Rice Plant Diseases and Pest Control.Beijing:Jindun Press,20-55.[黃世文,2010.水稻主要病蟲害防控關鍵技術解析.北京:金盾出版社,20-55]
Husnain T,Asad J,Maqbool SB,Datta SK,Riazuddin S,2002.Variability in expression of insecticidal cry1Ab gene in indica basmati rice.Euphytica.,128:121-128.
James C,2009.Global Status of Commercialized Biotech/GM Crops:2009.ISAAA Brief No.41.ISAAA:Ithaca,NY,USA.
James C,2010.Global Status of Commercialized Biotech/GM Crops:2010.ISAAA Brief No.42.ISAAA:Ithaca,NY,USA.
James C,2011.Global status of commercialized biotech/GM Crops:2011.ISAAA Brief No.43.ISAAA:Ithaca,NY,USA.
Khanna H,Raina S,2002.Elite indica transgenic rice plants expressing modified cry1Ac endotoxin of Bacillus thuringiensis show enhanced resistance to yellow stem borer.Transgenic Res.,11:411-423.
Krieg A,Huger AM,Langenbruch GA,Schnetter W,1983.Bacillus thuringiensis var tenebrionis:a new pathotype effective against larvae of coleopteran.J.Appl.Entomol.,96:500-508.
Li YH,Li XJ,Peng YF,2011.Global development of herbicide-tolerant transgenic crops and a strategic prospect for China.Plant Prot.,37(6):32-37.[李云河,李香菊,彭于發.2011.轉基因耐除草劑作物的全球開發與利用及在我國的發展前景和策略.植物保護,37(6):32-37]
Marfa V,Mele E,Gabarra R,Vassal J,Guiderdoni E,Meaaeguer J,2002.Influence of the developmental stage of transgenic rice plants expressing the Cry1B gene on the level of protection against the striped stem borer,Chilo suppressalis.Plant Cell Rep.,20:1167-1172.
Mochizuki A,Nishizawa Y,Onodera H,Tabei Y,Toki S,Habu Y,Ugaki M,Ohashi Y,1999.Transgenic rice plants expressing a trypsin inhibitor are resistant against rice stem borers,Chilo suppressalis.Entomol.Exp.Appl.,93:173-178.
Nagadhara D,Ramesh S,Pasalu IC,Rao YK,Krishnaiah NV,Sarma NP,Bown DP,Gatehouse JA,Reddy VD,Rao KV,2003.Transgenic indica rice resistant to sap-sucking insects.Plant Biotechnol.J.,1(3):231-240.
Nayak P,Basu D,Das S,Basu A,Ghosh D,Ramakrishnan NA,Ghosh M,Sen SK,1997.Transgenic elite indica rice plants expressing cry1Ac δ endotoxin of Bacillus thuringiensis are resistant against Scirophaga incertulas.Proc.Natl.Acad.Sci USA.,94:2111-2116.
Peng YF,Wu KM,Li YH,Lu YH,Yang XG,Ye GY,Li XH,2010-2011.Advances in biosafety subject of GMO.In:China Association for Science and Technology and China Society of Plant Protection eds.Report on Advances in Plant Protection.Beijing:Chinese Scientific and Technical Press,146-157.[彭于發,吳孔明,李云河,陸宴輝,楊曉光,葉汞銀,李新海,2010-2011.轉基因生物安全學學科發展研究.見:中國科學技術協會和中國植物保護學會編著.植物保護學學科發展報告.北京:中國科學技術出版社,146-157]
Ramesh S,Nagadhara D,Reddy VD,Rao KV,2004.Production of transgenic indica rice resistant to yellow stem borer and sap-sucking insects,using super-binary vectors of Agrobacterium tumefaciens.Plant Sci.,166:1077-1085.
Saha P,Majumder P,Dutta I,Ray T,Roy SC,Das S,2006.Transgenic rice expressing Allium sativum leaf lectin with enhanced resistance against sap-sucking insect pests.Planta,223(6):1329-1343.
Sharma HC,Sharma KK,Crouch JH,2004.Genetic transformation of crops for insect resistance:potential and limitations.Crit.Rev.Plant Sci.,23:47-72.
Sheng CF,Wang HT,Gao LD,Xuan WJ,2003.Occurrence state,damage assessment and control strategies.Plant Prot.,29(1):37-39.[盛承發,王紅托,高留德,宣維健,2003.我國水稻螟蟲大發生現狀、損失估計及防治對策.植物保護,29(1):37-39]
Shu QY,Ye GY,Cui HR,Cheng XY,Xiang YB,Wu DX,Gao MW,Xia YW,Hu C,Sardana R,2000.Transgenic rice plants with a synthetic cry1Ab gene from Bacillus thuringiensis were highly resistant to eight lepidopteran rice pest species.Mol.Breed.,6:433-439.
Tang KX,Sun XF,Hu QN,Wu AZ,Lin CH,Lin HJ,Twyman RM,Christou P,Feng TY,2001.Transgenic rice plants expressing the ferredoxin-like protein(API)from sweet pepper show enhanced resistance to Xanthomonas oryzae.Plant Sci.,160:1035-1042.
Tang W,Chen H,Xu CG,Li XH,Lin YJ,Zhang QF,2006.Development of insect-resistant transgenic indica rice with a synthetic cry1C gene.Mol.Breed,18:1-10.
Tu JM,Zhang GA,Datta K,Xu CG,He YQ,Zhang QF,Khush GS,Datta SK,2000.Field performance of transgenic elite commercial hybrid rice expressing Bacillus thuringiensis δ-endotoxin.Nat.Biotechnol.,18:1101-1104.
Wu KM,Guo YY,Gao SS,2002.Evaluation of the natural refuge function for Helicoverpa armigera(Lepidoptera:Noctuidae)within Bacillus thuringiensis transgenic cotton growing areas in north China.J.Econ.Entomol.,95:832-837.
Xu D,Duan X,Wang B,1996.Expression of a late embryogenesis abundant protein gene,Hval,from barley confers tolerance to water deficit and salt stress in transgenic rice.Plant Physiol.,110:249-257.
Yang Z,Peng XM,Peng WZ,Pang AJ,Pang BL,2011.Advances on resistance genes to rice brown planthopper.Biotechnol.Bull.,8:15-20.[楊震,彭選明,彭偉正,龐愛軍,龐伯良,2011.水稻褐飛虱抗性基因研究進展.生物技術通報,8:15-20]
Ye GY,Shu QY,Yao HW,2001.Field evaluation of resistance of transgenic rice containing a synthetic cry1Ab gene from Bacillus thuringiensis Berliner to two stem borers.J.Econ.Entomol.,94:271-276.
Ye GY,Yao HW,Shu QY,2003.High levels of stable resistance in transgenic rice with a cry1Ab gene from Bacillus thuringiensis Berliner to rice leaffolder,Cnaphalocrocis medinalis(Guenée)under field conditions.Crop Prot.,22:171-178.
Ye RJ,Huang HQ,Yang Z,Chen TY,Liu L,Li XH,2009.Development of insect-resistant transgenic rice with cry1C-free endosperm.Pest Manag.Sci.,65:1015-1020.
Zhang J,Shu CL,Zhang CG,2011.Current state and trends of patent protection for insecticidal genes from Bacillus thuringiensis.Plant Prot.,37(3):1-12.[張杰,束長龍,張春鴿,2011.Bt 殺蟲基因專利保護現狀與趨勢.植物保護,37(3):1-12]
Zhu Z,2010.Progress in research and development of transgenic rice.J.Agri.Sci.Technol.,12(2):9-16.[朱禎,2010.轉基因水稻研究進展.中國農業科技導報,12(2):9-16]
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
