匍枝根霉TP-02 內切葡聚糖酶基因eg2 的克隆表達及功能分析*
2013-10-30 03:34:20湯斌張瑩瑩楊亞平
食品與發酵工業 2013年7期
湯斌,張瑩瑩,楊亞平
(安徽工程大學 微生物發酵安徽省工程技術研究中心,安徽 蕪湖,241000)
木質纖維素在地球上的分布十分廣泛且蘊藏量豐富,是最廉價的可再生性生物質能源[1]。纖維素酶為多組分酶系,主要包括內切葡聚糖酶(endoglucanases,EG)、外 切 葡 聚 糖 酶(cellobiohydralases,CBH)和β-葡萄糖苷酶(β-glucosidases,BG),三者通過協同作用,分解纖維素獲得各種寡糖以及纖維二糖,最終水解生成葡萄糖等單糖,繼而可運用于工業生產制取酒精、醫藥用品、食品以及其它化工原料等,具有良好的應用前景[2-3]。其中,內切葡聚糖酶主要對纖維素內部的非結晶區起作用,通過隨機切割斬斷β-1,4-糖苷鍵,釋放出長度不等的短鏈低聚糖,從而在纖維素的降解過程中起著至關重要的作用[4-5]。目前,如何提高內切葡聚糖酶活力及產量已成為了研究的一大熱點。國內外對內切葡聚糖酶的研究主要集中于基因克隆表達和產酶條件優化,現已從多種真菌如里氏木霉、斜臥青霉、米根霉等克隆得到內切葡聚糖酶基因,并成功構建基因工程菌[6-8]。而對于內切葡聚糖酶結構與功能的研究,尤其是其催化過程中關鍵基團的報道還較少。
纖維素酶分子通常由纖維素結合域(CBD)、催化結構域(CD)及長短各異的連接片段(linker)組成[9-10]。CBD 由各種纖維素結合模塊(CBM)組成,其功能為吸附于纖維素表面并將CD 呈遞到底物之上,以疏解結晶纖維素的結構[11];CD 呈現出對底物的特異性,可以獨立發揮催化作用[12]。本文從匍枝根霉TP-02 的cDNA 文庫中克隆得到eg2 基因,并對其基因產物EGⅡ的CBM 及CD 的結構功能進行分析,在此基礎上確定參與酶解過程的重要殘基,并對其進行突變分析,以期通過基因修飾的手段提高內切葡聚糖酶的催化活力。
1 材料與方法
1.1 菌株、質粒和引物
匍枝根霉TP-02 菌株由本實驗室從黃山生態林的腐木中分離并保存;大腸桿菌(Escherichia coli)BL21 菌株和pET28a 質粒均由本實驗室保存。引物均由上海生工合成。
1.2 試劑及培養基
Trizol 試劑、mRNA 抽提試劑盒、PCR 產物純化試劑盒、質粒抽提試劑盒均購自上海生工,cDNA 反轉錄試劑盒、T4 DNA 連接酶及各種限制性內切酶均購于TaKaRa 公司(大連)。
誘導培養基:CMC-Na 0.4%,KH2PO40.2%,(NH4)2SO40.14%,尿素、MgSO4和CaCl2各0.03%,微量元素(FeSO45 mg/L,MnSO41.5 mg/L,ZnSO41.4 mg/L,CoCl22.0 mg/L,蛋白胨0.75 mg/L,酵母膏0.25 mg/L)。篩選培養基(CMC-Na):CMC-Na 1.5%,(NH4)2SO40.3%,KH2PO40.1%,MgSO40.05%,瓊脂2%。含剛果紅篩選培養基:配制10 mg/mL 剛果紅溶液,滅菌后按1 mL/200 mL 的比例加到CMC-Na 篩選培養基中,混勻后倒平板。
PDA 培養基及LB 培養基配方參見分子克隆指南。
1.3 PCR 擴增eg2 基因
將匍枝根霉TP-02 擴大培養后轉入誘導培養基中,誘導72 h 后離心收集菌體,進行冷凍干燥。取100 mg 菌絲體在預冷的研缽中快速研磨,利用Trizol試劑提取總RNA,并根據試劑盒法純化mRNA,以其為模板利用cDNA 試劑盒反轉錄合成cDNA 第一鏈。設計引物擴增目的基因,其上下游引物分別為F1 和R1(下劃線所示為酶切位點):
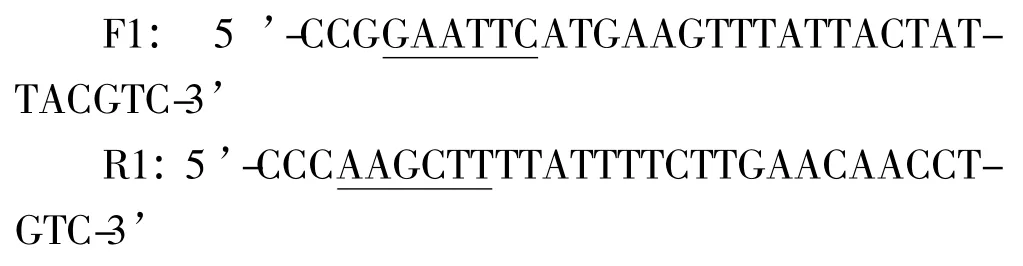
反應條件為:95℃預變性5 min;94℃變性30 s,56℃退火30 s,72℃延伸75 s,共30 個循環;72℃終延伸10 min。
1.4 重組質粒的構建及陽性克隆篩選
將純化的PCR 產物和pET28a 質粒分別經EcoRⅠ和Hind Ⅲ雙酶切后連接轉化,構建表達質粒pET28a-eg2。將重組質粒導入BL21 中,加入IPTG 至終濃度為1 mmol/L 涂布于含Kan+的LB 平板上,將形成的單菌落依次點種于含剛果紅的CMC-Na 篩選培養基上,并加入IPTG,37℃培養至長出單菌落,挑取透明圈較大的單菌落保種,用于酶切驗證、測序及發酵性能的研究。
1.5 重組菌株的誘導表達
挑取陽性克隆單菌落,接種于LB 液體培養基中,37℃,200 r/min 振蕩培養至OD600達0.8 左右,加入100 mmol/L 的IPTG 使其終濃度為1 mmol/L,進行誘導表達。每3 h 取樣,分別取1 mL 菌液,5 000 r/min 離心10 min,去上清液收集菌體,用0.05 mol/L檸檬酸緩沖液(pH 5.0)重懸細胞,-80℃反復凍融進行破胞,5 000 r/min 離心30 min,取上清液測酶活,并進行SDS 聚丙烯酰胺凝膠電泳(5%濃縮膠,12%分離膠)。
1.6 酶活測定
采用DNS 法測定內切葡聚糖酶的活力[13]。酶活定義:在酶的催化下,每分鐘水解CMC-Na 生成1 μmol 葡萄糖所需的酶量為1 個酶活力國際單位IU。
1.7 eg2 基因編碼蛋白質的生物信息學分析
利用PROSITE 數 據 庫(http://prosite. expasy.org/)預測EGII 的主要功能位點,并借助CDD 數據庫(http://www. ncbi. nlm. nih. gov/Structure/cdd/wrpsb.cgi)對EGII 氨基酸序列的保守結構域進行分析。利用ClustalX1.8 軟件對EGII 各保守結構域及其相應的同源序列進行多序列比對,并用BioEdit 軟件進行分析。利用Phyre2 在線分析引擎(http://www.sbg.bio.ic.ac.uk/ ~phyre/index.cgi)完成EGII三級結構的同源建模,并用Discovery Studio 2.5 軟件進行分析。
1.8 eg2 的基因修飾
根據eg2 生物信息學分析結果,確定突變位點并設計突變組。以pET28a-eg2 為模板,利用重疊PCR完成eg2 基因的修飾。各突變組如表1 所示;各組引物見表2,下劃線為突變位點,其中A1,B1 參與各突變組的PCR 過程。PCR 條件如下:95℃預變性5 min;95℃變性30 s,54℃/55℃退火45 s,72℃延伸50 s,共22 ~25 個循環;72℃終延伸10 min。將突變體酶切后與pET28a 連接,測序并轉染大腸桿菌BL21,測定各突變株的酶活。
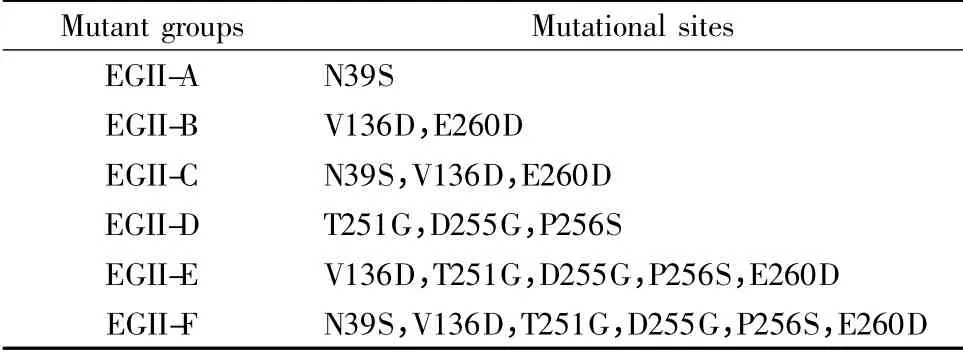
表1 突變組設計Tabel 1 Design of mutant groups
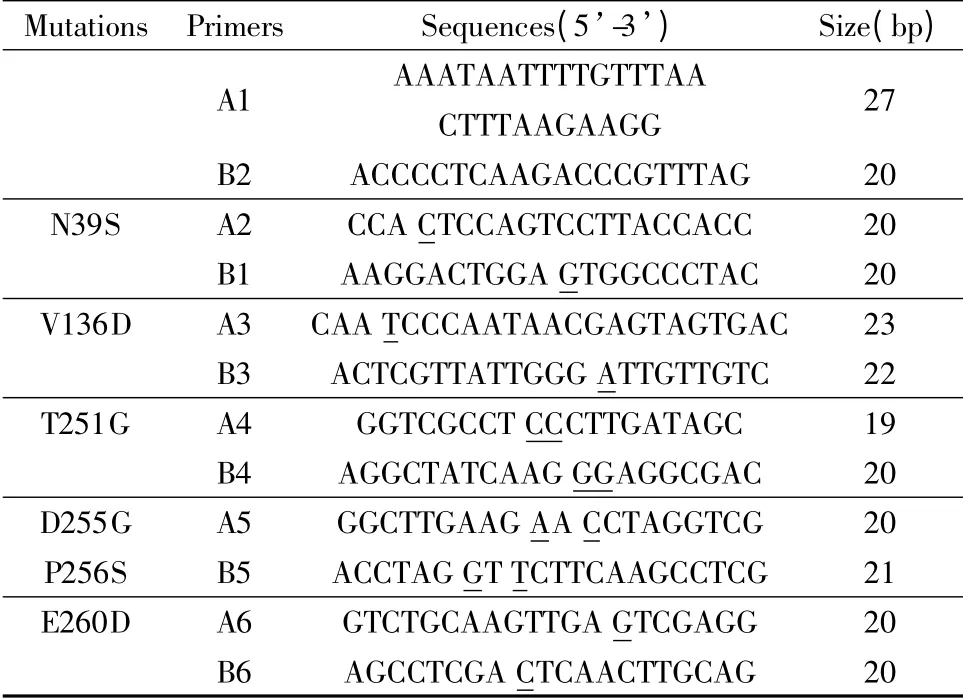
表2 重疊PCR 的引物序列Tabel 2 The sequences of primers for overlapping PCR
2 結果
2.1 eg2 基因的克隆
提取得到高質量的總RNA,并從中純化獲得mRNA,經逆轉錄酶作用,成功合成匍枝根霉TP-02的cDNA 單鏈,并從中克隆得到一段954 bp 的新型內切聚糖酶基因,稱為eg2 基因,將其上傳至Gen-Bank,獲取登錄號為JX315341。雙酶切驗證結果見圖1。
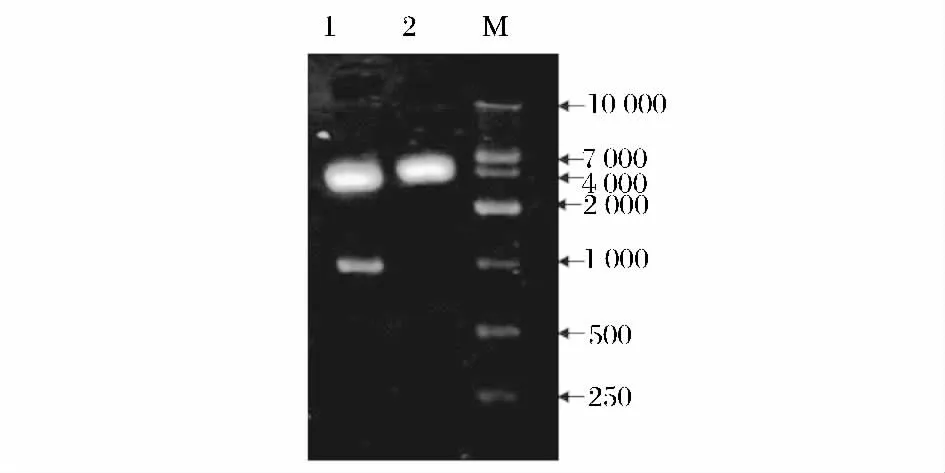
圖1 重組質粒雙酶切驗證Fig.1 Identification of recombinant plasmid
2.2 陽性克隆篩選
剛果紅與大分子多糖可牢固結合,隨著多糖分子量的變化,剛果紅結合程度就會發生變化,導致顏色從深紅變為淡黃。隨著菌落的長大,CMC 被不斷降解為寡糖,在菌落周圍就會形成透明圈,其大小由CMC 酶的降解能力所決定。透明圈越大,酶活越高。故而能夠快速地分辨出陽性克隆。圖2 所示為陽性克隆所形成的透明圈,空白對照A 在篩選平板上無法正常生長,未形成透明圈。

圖2 剛果紅染色結果Fig.2 The result of Congo red stain
2.3 EGII 功能位點及保守結構域預測
經PROSITE 分析,EGII 序列中含有1 個碳水化合物結合區(CBM1),該功能區的位置為氨基序列的25 ~61 位。CDD 數據庫的分析結果顯示,該蛋白質歸屬于糖基水解酶第45 家族(Glycoside Hydrolase Family 45),包含2 個GH45 家族的特征序列,并含有1 個典型的真菌纖維素結合區域即CBM1,該纖維素結合區域由4 個半胱氨酸構成,與PROSITE 預測結果一致。
2.4 EGII 氨基酸序列分析結果
2.4.1 CBM 模塊的比對分析
利用ClustalX 對EGII 的CBM1 序列進行同源比對,發現其在第15 位(即全序列的39 位)上的絲氨酸保守殘基被天冬酰胺所取代,該殘基參與CBM 親水性平面的形成[14],故該位點的突變可能會導致纖維素酶結合區域的結構改變,而對其酶活產生影響。以此確定CBM 的突變位點為N39S。
2.4.2 催化結構域GH45 特征序列分析結果
內切葡聚糖酶的催化區域通常包含兩個天冬氨酸催化殘基[1,15],通過序列比對發現EGII 中相對應的位置為136 位和260 位,均被其他殘基所取代,故確定其突變位點為V136D 和E260D。此外,文獻中報道在催化中心的一側存在一段loop 環,該環能增強催化基團周圍環境的疏水性,阻止除水分子外的各種親核試劑進入活性部位[15]。經序列比對后,確定該loop 環的突變位點為T251G、D255G 和P256S。利用Phyre2 完成EGII 的催化區GH45 及其突變體的三維建模,結果如圖3 所示。催化殘基V136D 及E260D 的突變形成拉近了兩者的距離,以其為中心的活性區域發生改變。此外,對位于V136 和E260 中間的一段loop 環進行改造,T251G、D255G 及P256S的突變形成使得該loop 環移至活性中心一側,同時改變催化殘基D260 的空間位置。
2.5 發酵性能實驗
將含eg2 基因的大腸桿菌重組菌及各突變株接到LB 液體培養基中進行發酵培養,每隔3 h 取樣測CMC 酶活,結果如圖4 所示。EGⅡ-A 至EGⅡ-F 的最高酶活分別為0.897 IU/mL,1.114 IU/mL,1.286 IU/mL,0.868 IU/mL,1.215 IU/mL 和1.321 IU/mL。各突變株的酶活均高于原始菌株,其中EGⅡ-F 的酶活比EGⅡ(0.723 IU/mL)提高了82.7%。含CBM1突變位點N39S 的突變株EGⅡ-A、EGⅡ-B 和EGⅡ-F比其他重組菌更早達到峰值,而所有含V136D 及E260D 的突變株均具有較高的酶活。單獨對loop 環進行突變的重組菌EGⅡ-C 酶活較低。
2.6 SDS-PAGE
取以上酶活達到峰值時的發酵液進行SDS 聚丙烯酰胺凝膠電泳,以未誘導的EGⅡ為空白對照,結果如圖5 所示。表達條帶的分子量大小約為45 kDa,與理論值相符。其中,EGⅡ-E 和EGⅡ-F 的表達條帶較粗,表達量相對較高。
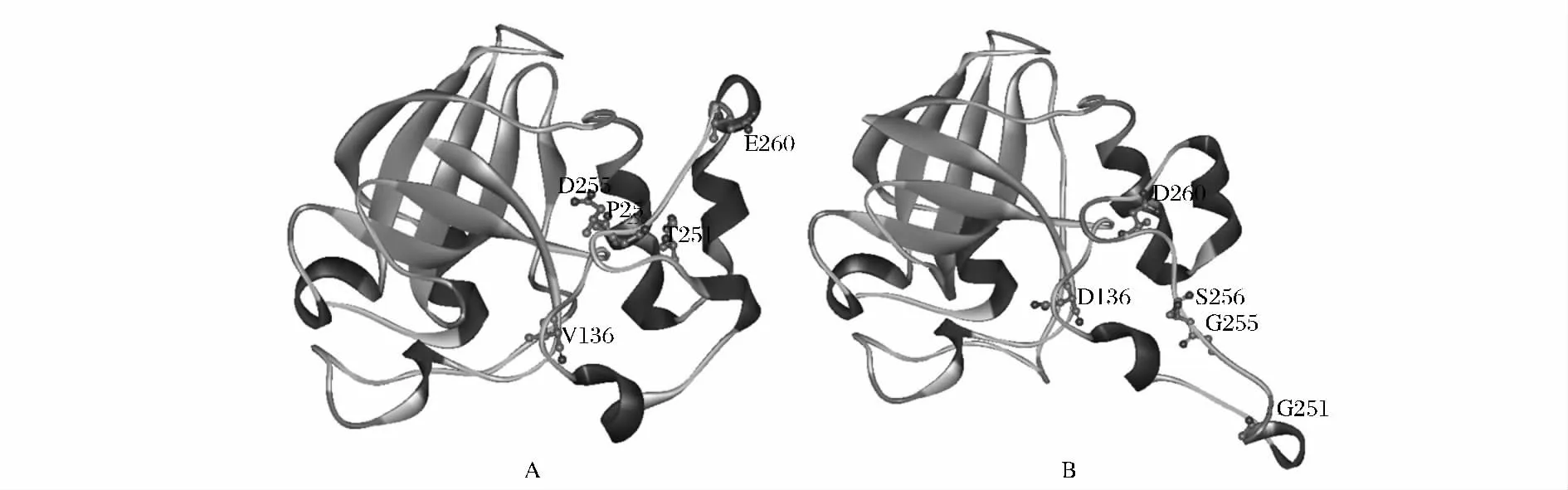
圖3 GH45 三級結構模型Fig.3 Tertiary structure of GH45 domain
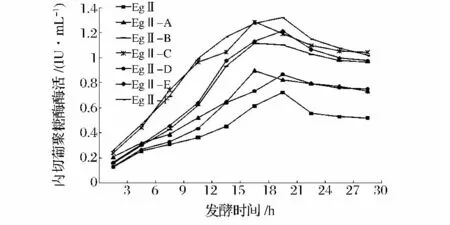
圖4 重組菌的發酵特性Fig.4 Fermentation characteristics of different recombinant E. coli BL21
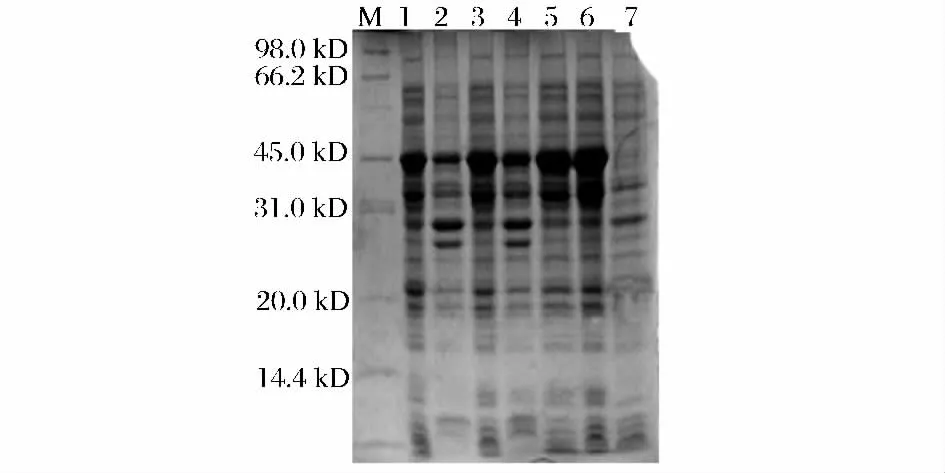
圖5 SDS-PAGE 結果Fig.5 Results of SDS-PAGE.
3 討論
本文成功克隆了匍枝根霉的內切葡聚糖酶基因,并實現了其在大腸桿菌中的表達,對其結構功能進行了初步探討,發現纖維素結合模塊CBM1 和催化結構域GH45 區在纖維素降解過程中起著關鍵作用。突變分析結果顯示,N39S 突變的形成可能會對CBM1與纖維素結合能力產生影響,從而改變內切葡聚糖酶的水解效率。V136D 及E260D 兩個催化位點的突變形成對酶活有明顯的提高作用,而單獨對loop 環進行突變只能對酶活產生微弱的影響,即在酶解過程中該loop 環只起輔助作用。然而,對于CBM1 和GH45結構域的具體作用機理還不甚明了,后續研究將著眼于纖維素酶的水解機制,及各保守殘基在催化過程中所扮演的角色。
[1] Gowen CM,Fong SS. Exploring biodiversity for cellulosic biofuel production[J]. Chemistry & Biodiversity,2010,7(5):1 086 -1 097.
[2] Takashima S,Ohno M,Hidaka M,et al. Correlation between cellulose binding and activity of cellulose-binding domain mutants ofHumicola griseacellobiohydrolase Ⅰ[J].FEBS Letters,2007,581(30):5 891 -5 896.
[3] 韋小敏. 斜臥青霉胞外蛋白質組學分析與纖維素酶合成調控機制研究[D]. 山東大學,2011:1 -2.
[4] Nakazawa H,Okada K,Onodera T,et al. Directed evolution of endoglucanaseⅢ(Cel12A)fromTrichoderma reesei[J]. Applied Microbiology Biotechnology,2009,83(4):649 -657.
[5] 鄭海英,黃平,蔡少麗,等. 特異腐質霉內切葡聚糖酶Ⅱ基因在畢赤酵母中的表達及酶學性質[J]. 微生物學通報,2012,39(2):145 -153.
[6] Samanta S,Basu A,Halder UC,et al. Characterization ofTrichoderma reeseiendoglucanase Ⅱ expressed heterologously in Pichia pastoris for better bionfinishing and biostoning[J]. J Microbiol,2012,50(3):518 -525.
[7] 劉韞滔,韓學鳳,羅澤宇,等. 斜臥青霉L-06 內切葡聚糖酶Ⅰ基因的克隆與表達[J]. 微生物學通報,2012,39(5):696 -701.
[8] Moriya T,Murashima K,Nakane A,et al. Molecular cloning of endo-beta-D-1,4-glucanase genes,rce1,rce2,and rce3,fromRhizopus oryzae[J]. J Bacteriol,2003,185(5):1 749 -1 756.
[9] Hirvonen M,Papageorgiou AC. Crystal structure of a family 45 endoglucanase fromMelanocarpus albomyces:mechanistic implications based on the free and cellobiose-bound forms[J]. Journal of Molecular Biology,2003,29(3):403 -410.
[10] 方詡,秦玉琪,李雪芝,等. 纖維素酶與木質纖維素生物降解轉化的研究進展[J]. 生物工程學報,2010,26(7):864 -869.
[11] Carrard G,Koivula A,Soderlund H,et al. Cellulosebinding domains promote hydrolysis of different sites on crystalline cellulose[J]. Proc Natl Acad Sci USA,2000,97(19):10 342 -10 347.
[12] Warner CD,Camci-Unal G,Pohl NLB,et al. Substrate binding by the catalytic domain and carbohydrate binding module ofRuminococcus flavefaciensFD-1 xyloglucanase/endoglucanase[J]. Industrial & Engineering Chemistry Research,2013(2):30 -36.
[13] 陳紅漫,張欣,李艷秋,等. 綠色木霉葡聚糖內切酶(EGⅠ)基因克隆及在釀酒酵母中的表達[J]. 食品與發酵工業,2012,38(6):48 -52.
[14] Kraulis PJ,Clore GM,Nilges M,et al. Determination of the three-dimensional solution structure of the C-terminal domain of cellobiohydrolase I fromTrichoderma reesi. A study using nuclear magnetic resonance and hybrid distance geometry-dynamical simulated annealing[J]. Biochemistry,1989,28(18):7 241 -7 257.
[15] Valjakka J,Rouvinen J. Structure of 20K endoglucanase fromMelanocarpus albomycesat 1.8 resolution[J]. Biological Crystallography,2003,59:765 -768.
