基于COI和ITS-1基因?qū)酪吧后w和人工繁育子代的遺傳分析
2013-10-13 08:14:24邱兆星王鴻霞張秀梅劉保忠
海洋科學(xué) 2013年7期
陳 楨 , 邱兆星, 王鴻霞, 張秀梅 劉保忠
(1. 中國(guó)海洋大學(xué) 海水養(yǎng)殖教育部重點(diǎn)實(shí)驗(yàn)室, 山東 青島 266003; 2. 中國(guó)科學(xué)院 實(shí)驗(yàn)海洋生物學(xué)重點(diǎn)實(shí)驗(yàn)室, 山東 青島 266071; 3. 山東省海水養(yǎng)殖研究所, 山東 青島 266002)
魁蚶(Scapharca broughtoniiSchrenck)俗稱赤貝、血貝、大毛蚶, 隸屬軟體動(dòng)物門(Mollusca), 雙殼綱(Bivalvia), 翼形亞綱(Pterimorphia), 蚶目(Arcoida), 蚶科(Arcidae), 是一種大型經(jīng)濟(jì)貝類, 主要分布于中國(guó)、日本、朝鮮半島及俄羅斯東南部沿海。在我國(guó)以黃海北部較多, 主要在山東及遼寧沿海分布[1]。迄今為止,對(duì)魁蚶的研究工作已開展較多, 包括生殖生物學(xué)和苗種繁育[2-6]、資源生態(tài)[7-8], 群體遺傳結(jié)構(gòu)[9-12]等。
山東榮成瀕臨黃海北部, 沿岸淺海水域歷來就有魁蚶這一優(yōu)勢(shì)種質(zhì)資源的自然分布。20世紀(jì)80年代初, 海岸帶生物資源調(diào)查結(jié)果發(fā)現(xiàn), 魁蚶自然資源在榮成海域的集中分布面積約7000 ha, 其蘊(yùn)藏量超過2萬t , 平均生物密度達(dá)0.476個(gè)/m2, 是榮成近岸水域資源開發(fā)的重要經(jīng)濟(jì)種[13]。自 20世紀(jì) 90年代以來, 由于資源過度開發(fā), 使原有的魁蚶資源受到極大破壞,自然種群生物量顯著下降。目前, 榮成海域內(nèi)魁蚶自然資源分布區(qū)域面積銳減, 按原始資源面積計(jì)算, 自然生物量少于0.01個(gè)/m2[13]。因此, 榮成市海洋與漁業(yè)局廣泛組織、發(fā)動(dòng)沿海魁蚶資源引進(jìn)、增殖和修復(fù)技術(shù)。
近年來, 為滿足魁蚶資源保護(hù)和增殖的需要,魁蚶的底播增養(yǎng)殖研究日益受到重視, 并取得一定的生態(tài)和經(jīng)濟(jì)效益[14-17]。在資源增殖過程中, 人們更多地關(guān)注資源量的增加, 而對(duì)人工苗種對(duì)本底生物資源可能帶來的遺傳影響重視不夠。不合理的繁育和增殖方式可能導(dǎo)致近交快速積累、有效群體不斷減少等負(fù)面效果, 引起近交衰退, 使種苗遺傳多樣性下降, 表現(xiàn)為生長(zhǎng)減緩、抗病力減弱、品質(zhì)退化等。有報(bào)道指出, 群體的遺傳多樣性每喪失 10%, 就會(huì)對(duì)其繁育能力、存活率、生長(zhǎng)等重要性狀產(chǎn)生很大的負(fù)面影響[18]。
本研究采用線粒體COI基因和核糖體ITS-1基因序列, 對(duì)榮成和即墨 2個(gè)魁蚶野生群體進(jìn)行遺傳多樣性研究, 探討了即墨群體人工繁育子一代群體的遺傳多樣性變化及其在榮成底播增殖的可行性。
1 材料與方法
1.1 實(shí)驗(yàn)材料
于 2009年 9月, 從榮成(編號(hào) RC)采集 26個(gè)野生魁蚶個(gè)體; 從即墨采集一批野生魁蚶(數(shù)量>100個(gè))作為親本, 通過人工育苗的方式得到一批子代,從親本群體中隨機(jī)取37個(gè)個(gè)體(編號(hào)JM), 從子代中隨機(jī)取40個(gè)個(gè)體(編號(hào)JZ)(子代生長(zhǎng)到2010年5月進(jìn)行采集), 共對(duì)103個(gè)魁蚶個(gè)體進(jìn)行分析。
1.2 實(shí)驗(yàn)方法
1.2.1 基因組DNA提取及PCR擴(kuò)增
采用TIANGEN海洋動(dòng)物組織基因組DNA提取試劑盒提取魁蚶的基因組DNA后, 保存-20℃待用。
先用 COI通用引物(COI L1490: 5′-GGTCAACAAATCATAAAGATATTGG-3′和 COI H2198: 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′)擴(kuò)增得到 2 個(gè)魁蚶個(gè)體的COI序列; 再根據(jù)得到的序列, 用Primer Premier 5.0軟件設(shè)計(jì)COI特異引物, 序列為: COI-1:5′-CAAATGTTGGAATAGTAAAGGA-3′; COI-2: 5′-TATTTTCAGGATTCTGGGCA-3′。所用的ITS-1引物[19]為 : ITS-1A: 5′-GGTTCTGTAGGTGAACCTGC-3′;ITS-1B: 5′-CTGCGCCATCGACCC-3′。
PCR 反應(yīng)體系 25 μL: ddH2O 17.25 μL; DNA 模板1 μL; 10×PCR buffer 2.5 μL; MgCl2(25 mmol/L)1.5 μL;dNTPs(10mM)0.5 μL; 引 物 F(10mM)1 μL; 引 物R(10mM)1 μL; Taq DNA Polymerase(5U/μL)0.25 μL。PCR反應(yīng)程序?yàn)? 94℃變性5 min, 1個(gè)循環(huán); 94℃變性20 s, 50℃/58℃退火30 s, 72 ℃延伸30 s, 34個(gè)循環(huán);72℃延伸10 min, 1個(gè)循環(huán); 4℃保存。
所有PCR擴(kuò)增在Takara T650 PCR儀上進(jìn)行。
1.2.2COI基因PCR產(chǎn)物純化、克隆及測(cè)序
PCR產(chǎn)物經(jīng)瓊脂糖凝膠電泳確定為單一目的條帶。用上海生工SanPrep柱式DNA膠回收試劑盒回收純化。將純化產(chǎn)物與pMD19-T 載體(大連寶生物)連接, 連接反應(yīng)體系為 5 μL(solution I 2.5 μL, vector 0.4 μL, 純化產(chǎn)物2.1 μL), 16℃連接過夜。連接產(chǎn)物42℃轉(zhuǎn)化到大腸桿菌 TOP10感受態(tài)細(xì)胞中, 并涂布于 LB平板上 37℃過夜。挑取邊緣清晰的菌落作模板, 用 M13 通用引物(M13 F: 5′-AGCGGATAACAATTTCACACAGG-3′; M13 R: 5′-CGCCAGGGTTTTCCCAGTCACGAC-3′)進(jìn)行PCR擴(kuò)增, 篩選陽性克隆。將陽性克隆擴(kuò)大培養(yǎng), 送交北京六合華大基因科技有限公司測(cè)序。
1.2.3ITS-1基因PCR產(chǎn)物純化及測(cè)序
PCR產(chǎn)物經(jīng)瓊脂糖凝膠電泳確定為單一目的條帶。用上海生工SanPrep柱式DNA膠回收試劑盒回收純化。將純化產(chǎn)物送交北京六合華大基因科技有限公司測(cè)序。
1.2.4 數(shù)據(jù)分析
所有序列均由Mega 4.0軟件[20]進(jìn)行編輯、校對(duì)和比對(duì), 并對(duì)比對(duì)結(jié)果進(jìn)行分析和手工校正, 并去除序列中不確定的部分。一共得到63個(gè)魁蚶野生個(gè)體(榮成26個(gè)和即墨37個(gè))和40個(gè)即墨人工繁育子代的 605bp的 COI基因序列 (NCBI登錄號(hào)為:KC588467- KC588503)和相應(yīng)的354bp的ITS-1基因序列(NCBI登錄號(hào)為: KC555313- KC555327)。
采用Mega 4.0軟件按照Kimura等[21]雙參數(shù)進(jìn)化模型計(jì)算單倍型間的遺傳距離, 構(gòu)建單倍型的鄰接關(guān)系樹, 并采用1000次重抽樣檢驗(yàn)系統(tǒng)樹分支的可靠性。
用DnaSP 5.0[22]軟件計(jì)算序列中核苷酸組成、多態(tài)位點(diǎn)數(shù)(S)、核苷酸多樣性(π)、單倍型數(shù)(h)和單倍型多樣性指數(shù)(Hd), 分析群體的多樣性水平。
利用 Arlequin3.11軟件[23]計(jì)算單倍型間的關(guān)系和核苷酸替換數(shù), 手工繪制最小跨度樹(Minimum spanning tree)。采用Arlequin 3.11軟件計(jì)算魁蚶群體之間的遺傳分化指數(shù)Fst值來檢驗(yàn)群體遺傳結(jié)構(gòu)。Fst值的顯著水平通過 10000次重抽樣來檢驗(yàn), 并用Bonferroni方法[24]對(duì)其進(jìn)行校正。
2 結(jié)果
2.1 序列分析
實(shí)驗(yàn)共獲得長(zhǎng)度為605bp的COI基因序列片段和長(zhǎng)度為354bp的ITS-1基因序列片段各103條。COI基因片段的T、C、A、G堿基平均含量分別為23.7%,21.4%, 40.1%, 14.8%, A+T含量明顯高于C+G含量,而ITS-1基因片段的 T、C、A、G堿基平均含量分別為 25.6%, 26.9%, 20.1%, 27.4%, A+T含量低于C+G含量。
2.2 遺傳多樣性
在COI基因序列的605個(gè)位點(diǎn)中, 多態(tài)位點(diǎn)73個(gè), 其中簡(jiǎn)約信息位點(diǎn)29個(gè)。這些多態(tài)位點(diǎn)共定義了 37個(gè)單倍型(表 1), 其中有 Hap1出現(xiàn)頻率最高,為37.8%的即墨野生(親代)群體和77.5%的即墨子代群體所共享, 在榮成野生群體中未檢測(cè)到單倍型Hap1; Hap4為18.9%的即墨野生群體與61.5%的榮成野生群體共享, 而其他35個(gè)單倍型為各個(gè)群體私有且只在一個(gè)個(gè)體中發(fā)現(xiàn)。即墨野生群體中私有單倍型占總單倍型的 43.3%, 榮成野生群體的私有單倍型占總單倍型的 38.5%, 即墨子代群體的私有單倍型占總單倍型的22.5%。

表1 COI序列單倍型在各群體中分布Tab.1 Distribution of COI haplotypes of S. broughtonii among localities
在ITS-1基因序列的354個(gè)位點(diǎn)中, 檢測(cè)到插入/缺失3處, 多態(tài)位點(diǎn)10個(gè), 其中簡(jiǎn)約信息位點(diǎn)7個(gè)。這些多態(tài)位點(diǎn)共定義了15個(gè)單倍型(表2), 其中有5個(gè)單倍型為3個(gè)群體共有享, H1是出現(xiàn)頻率最高的單倍型, 有 2個(gè)單倍型為即墨野生群體和即墨子代群體共享, 其他 8個(gè)單倍型為各個(gè)群體私有且只在一個(gè)個(gè)體中發(fā)現(xiàn)。
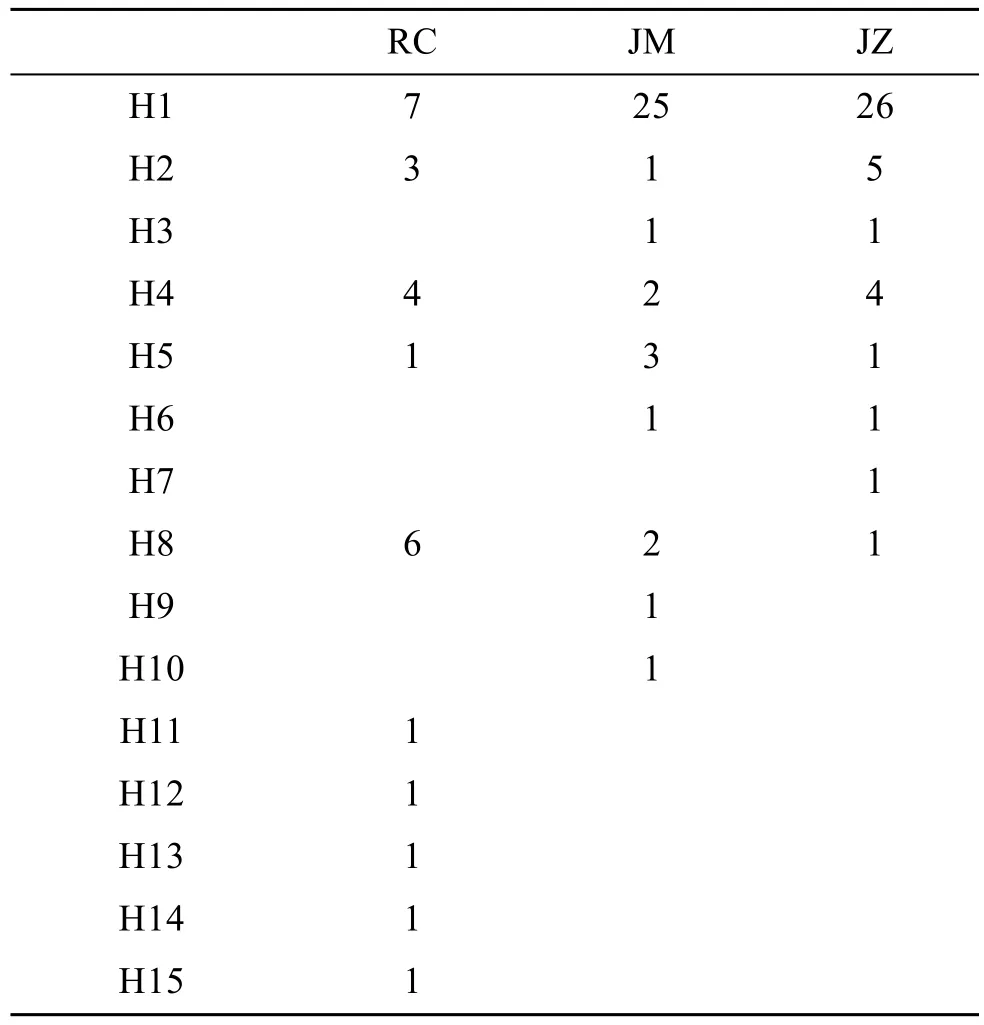
表2 ITS-1序列單倍型在各群體中分布Tab.2 Distribution of ITS-1 haplotypes of S. broughtonii among localities
基于多態(tài)位點(diǎn)得到各群體的核苷酸多樣性指數(shù)(π),多態(tài)位點(diǎn)數(shù)(S), 單倍型數(shù)(h)和單倍型多樣性指數(shù)(Hd)見表3和表4。分析COI基因序列發(fā)現(xiàn), 榮成和即墨野生群體的單倍型多樣性指數(shù)分別為0.631和0.832, 核苷酸多樣性指數(shù)分別為0.0015和0.0252; 即墨人工培育子代群體的單倍型多樣性指數(shù)為0.404, 核苷酸多樣性指數(shù)為 0.0008, 顯然比 2個(gè)野生群體的多樣性指數(shù)低。分析ITS-1基因序列的結(jié)果, 發(fā)現(xiàn)野生群體和人工培育即墨子代群體間的單倍型多樣性和核苷酸多樣性指數(shù)差異不大。
2.3 系統(tǒng)發(fā)育關(guān)系
基于COI基因片段定義的37個(gè)單倍型的鄰接關(guān)系樹顯示, 所有單倍型明顯分為 2個(gè)類群(圖 1), 并得到很高的置信度支持。全部榮成野生群體的單倍型和部分即墨野生群體單倍型聚成類群Ⅰ, 部分即墨野生群體的單倍型和全部即墨子代群體的單倍型聚成類群Ⅱ。而基于ITS-1基因片段定義的15個(gè)單倍型的鄰接關(guān)系樹置信度低, 單倍型之間的關(guān)系無法確定(圖2)。

表3 魁蚶樣品數(shù)和基于線粒體 COI基因的多態(tài)位點(diǎn)數(shù)(S), 單倍型數(shù)(h), 單倍型多樣性(Hd)和核苷酸多樣性(π)及其標(biāo)準(zhǔn)差(Standard deviation, SD)Tab.3 Sample size and number of variable sites (S), number of haplotype (h), haplotype diversity (Hd), nucleotide diversity (π)of S. broughtonii based on COI. SD: Standard deviation
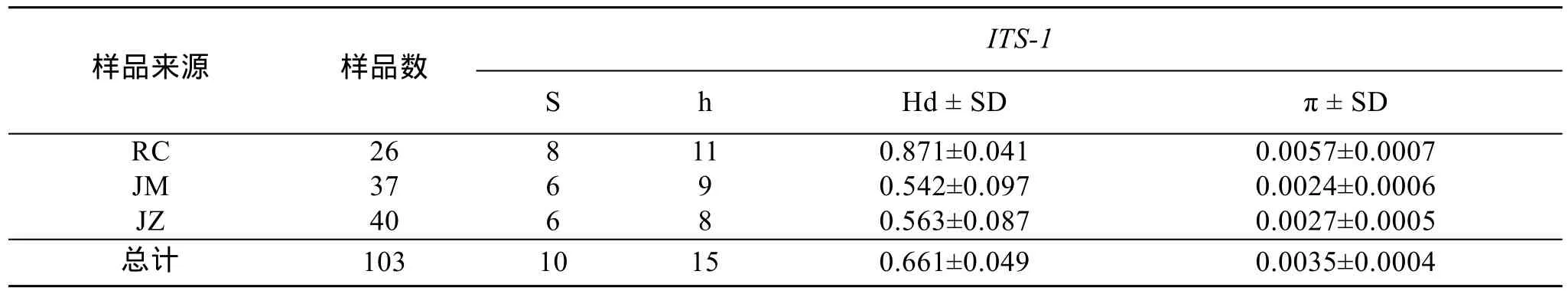
表4 魁蚶樣品數(shù)和基于核糖體內(nèi)轉(zhuǎn)錄間隔區(qū)ITS-1的多態(tài)位點(diǎn)數(shù)(S), 單倍型數(shù)(h), 單倍型多樣性(Hd)和核苷酸多樣性(π)及其標(biāo)準(zhǔn)差(Standard deviation, SD)Tab.4 Sample size and number of variable sites (S), number of haplotype (h), haplotype diversity (Hd), nucleotide diversity (π)of S. broughtonii based on ITS-1. SD: Standard deviation
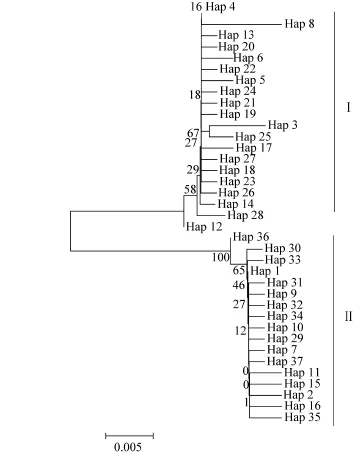
圖1 用Mega 4.0構(gòu)建的魁蚶COI基因單倍型間的鄰接關(guān)系樹Fig.1 Neighbor-joining tree of haplotypes of S. broughtonii based on COI sequences constructed by Mega 4.0
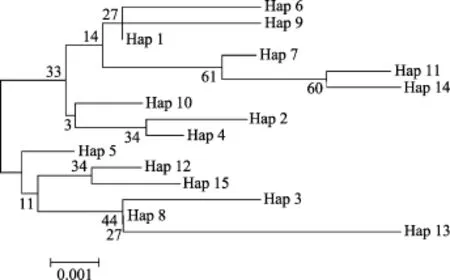
圖2 用Mega 4.0構(gòu)建的魁蚶ITS-1基因單倍型間的鄰接關(guān)系樹Fig.2 Neighbor-joining tree of haplotypes of S.broughtonii based on ITS-1 sequences constructed by Mega 4.0
基于COI和ITS-1基因構(gòu)建的單倍型最小跨度樹有相似的結(jié)構(gòu), 都呈星形輻射狀。處在星狀結(jié)構(gòu)中心的都為出現(xiàn)頻率最高的單倍型(基于COI和ITS-1基因分別為Hap1和H1), 如圖3和圖4與表1和表2。COI基因單倍型中, Hap1只在即墨野生和子代群體中出現(xiàn), 榮成群體中未出現(xiàn); 而ITS-1單倍型中,H1在3個(gè)群體中均有出現(xiàn)。
2.4 群體遺傳結(jié)構(gòu)
基于COI基因序列片段, AMOVA分析結(jié)果表明,魁蚶群體的群體間差異占 26.74%, 群體內(nèi)差異占73.26%, 分子差異主要在群體內(nèi)(表5)。基于ITS-1基因序列片段, AMOVA分析結(jié)果也表明分子差異主要在群體內(nèi), 群體間與群體內(nèi)分子差異分別占 6.86%和93.14%(表5)。2個(gè)基因序列分析得到的固定指數(shù)分別為 0.2674和 0.0686, 統(tǒng)計(jì)檢驗(yàn)結(jié)果都為極顯著(P<0.01), 表明魁蚶群體中存在明顯的遺傳結(jié)構(gòu)。表現(xiàn)群體間遺傳分化的Fst值見表 6。基于COI基因得到的榮成和即墨 2個(gè)野生魁蚶群體之間的Fst值為 0.168,基于ITS-1基因得到的兩群體間Fst值為0.1193, 表明2個(gè)野生群體之間存在一定程度的遺傳分化。
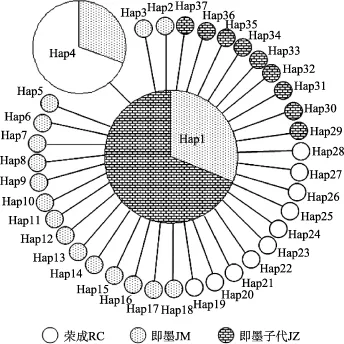
圖3 魁蚶37個(gè)COI基因單倍型的最小跨度樹Fig.3 Minimum spanning tree of the 37 haplotypes of the S.broughtonii based on COI sequences

圖4 魁蚶15個(gè)ITS-1基因單倍型的最小跨度樹Fig.4 Minimum spanning tree of the 15 haplotypes of the S.broughtonii based on ITS-1 sequences
3 討論
從本研究的結(jié)果看,COI基因的堿基含量分布不均勻, A+T含量高于C+G含量, 表現(xiàn)出明顯的堿基偏向性, 這與在其他無脊椎動(dòng)物的COI基因中觀察到的結(jié)果是一致的[25-26]。而ITS-1基因的堿基含量分布表現(xiàn)為A+T含量低于C+G含量, 陳琳琳[27]在日本、俄羅斯海參崴和中國(guó)威海 3個(gè)魁蚶地理群體中也發(fā)現(xiàn)類似結(jié)果, 這與其他貝類的ITS序列中觀察到的結(jié)果相似[28]。
遺傳多樣性降低在人工繁殖群體中是一個(gè)較普遍的現(xiàn)象。方耀林等[29]在采用 mtDNA控制區(qū)序列,對(duì)野生大鯢及其人工繁殖后代的遺傳多樣性進(jìn)行分析時(shí)發(fā)現(xiàn), 兩組親本的單倍型多樣性及核苷酸多樣性均高于各自的子代。王鴻霞等[30]發(fā)現(xiàn)凡納濱對(duì)蝦人工繁育的子代的等位基因數(shù)比親代低 11.11%, 平均期望雜合度、平均觀測(cè)雜合度和平均多態(tài)性信息含量等指標(biāo)均低于親代。已有群體遺傳學(xué)研究表明,遺傳多樣性的保持和有效群體大小密切相關(guān)。當(dāng)有效群體較小時(shí), 由于瓶頸效應(yīng), 野生或養(yǎng)殖群體傾向于近交水平的增加[31], 引起種群遺傳多樣性水平的下降。在本研究中, 雖然采用了100個(gè)個(gè)體以上規(guī)模的親本群, 但基于COI基因序列片段的結(jié)果看,即墨人工繁育的子代群體的單倍型多樣性和核苷酸多樣性明顯比即墨野生群體低。又有研究證實(shí), 在水產(chǎn)動(dòng)物的自然繁育中, 只有很少的親本參與了子代基因庫的構(gòu)成, 有效群體比繁育親本群體要小的多。如Sekino等[32]利用4個(gè)高度多態(tài)性的微衛(wèi)星位點(diǎn)對(duì)牙鲆子代群體進(jìn)行了家系鑒定, 結(jié)果表明不同的親本對(duì)子代的貢獻(xiàn)率不同, 18個(gè)親本中僅一半對(duì)后代基因有貢獻(xiàn)。Vandeputte等[33],王鴻霞等[30]在對(duì)子代群體進(jìn)行家系鑒定時(shí)也發(fā)現(xiàn)相似的現(xiàn)象。也就是說,親本數(shù)量和親本對(duì)子代的不同貢獻(xiàn)率可能是造成子代群體遺傳多樣性低于親代群體的原因。
在對(duì)COI基因序列分析時(shí)發(fā)現(xiàn), 即墨人工繁育群體的核苷酸多樣性比野生親代群體有明顯降低,但基于ITS-1基因序列分析時(shí)發(fā)現(xiàn), 兩群體的多態(tài)位點(diǎn)數(shù)和核苷酸多樣性指數(shù)相差不大。同樣, 在即墨群體與榮成群體之間, 基于COI基因序列分析結(jié)果發(fā)現(xiàn)兩者的多態(tài)位點(diǎn)數(shù)和核苷酸多樣性指數(shù)差異顯著,兩群體的多態(tài)位點(diǎn)數(shù)分別為56和12, 核苷酸多樣性分別為0.0252和0.0015, 呈現(xiàn)明顯的倍數(shù)關(guān)系。而基于ITS-1基因序列分析卻發(fā)現(xiàn), 兩群體的多態(tài)位點(diǎn)數(shù)和核苷酸多樣性指數(shù)相差不大。這可能是由于線粒體基因比核基因的進(jìn)化速率快[34]和ITS-1基因種內(nèi)變異小[35]的特點(diǎn)造成的。ITS-1基因種內(nèi)變異小的現(xiàn)象在其他研究中也有發(fā)現(xiàn), 如Philips等[36]對(duì)不同地理種群的紅點(diǎn)鮭的ITS-1進(jìn)行測(cè)序分析, 發(fā)現(xiàn)種內(nèi)不同種群之間的序列相似性較大; 林昕等[37]采用ITS-1基因序列片段, 發(fā)現(xiàn)福建沿海不同養(yǎng)殖區(qū)泥蚶群體具有較高的遺傳相似性。因此, 作者認(rèn)為基于COI基因序列檢測(cè)魁蚶群體水平多態(tài)性比ITS-1基因更為敏感。

表5 3個(gè)魁蚶群體的分子方差分析Tab.5 Analysis of molecular variance(AMOVA)of COI and ITS-1 haplotypesof S. broughtonii populations
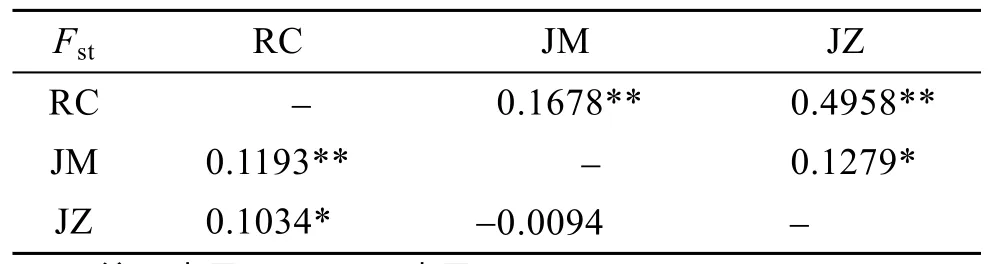
表6 基于COI(對(duì)角線以上)和ITS-1(對(duì)角線以下)基因序列單倍型分析得到的群體間遺傳分化指數(shù)Fst值Tab.6 Population pairwise(Fst)between S.broughtonii populations based on COI(above diagonal)和ITS-1(below diagonal)sequences
在單倍型鄰接關(guān)系樹中, 基于COI基因序列得到的單倍型以高置信度聚成2個(gè)類群, 而基于ITS-1基因序列得到的單倍型鄰接關(guān)系樹置信度低, 單倍型之間的關(guān)系無法確定, 該結(jié)果可能也與ITS-1基因的種內(nèi)變異小有關(guān)。在COI基因序列單倍型的鄰接關(guān)系樹中, 榮成野生群體和部分即墨野生群體聚在同一個(gè)類群(類群Ⅰ)中, 另一部分即墨野生群體與即墨子代聚在類群Ⅱ, 表明榮成和即墨野生群體之間存在某些程度的遺傳分化。根據(jù) Wright[38]對(duì)遺傳分化程度劃分的等級(jí),Fst為 0.05~0.15, 說明群體間存在中等程度的遺傳分化。而即墨和榮成野生群體之間的Fst值基于COI和ITS-1基因分別為0.1678和0.1193, 表明魁蚶2個(gè)野生群體之間的遺傳分化程度為中等水平。在本研究中, 基于COI和ITS-1基因構(gòu)建的單倍型最小跨度樹都為星狀結(jié)構(gòu), 這通常被認(rèn)為是種群經(jīng)歷過種群擴(kuò)張的結(jié)果[39], 處于星狀結(jié)構(gòu)中心的單倍型(COI和ITS-1基因序列中分別為Hap1和H1)推測(cè)為祖先單倍型。在COI單倍型分布中, 可能是祖先單倍型的 Hap1在榮成野生群體中未出現(xiàn),這可能與分析樣品數(shù)量有關(guān)。
2種基因序列片段的分子方差分析(AMOVA)結(jié)果都表明, 魁蚶 3個(gè)群體內(nèi)的分子遺傳變異明顯大于群體間的遺傳變異。研究中發(fā)現(xiàn), 相對(duì)于ITS-1基因的AMOVA分析結(jié)果, 基于COI基因序列分析時(shí)群體間的遺傳變異比例較大(26.74%)。進(jìn)行Fst分析時(shí)同樣發(fā)現(xiàn)這樣的問題, 基于COI基因序列得到的群體間Fst值比ITS基因序列得到的Fst值大。我們認(rèn)為這可能是由于2個(gè)基因的進(jìn)化速率不同和ITS-1種內(nèi)變異小的特性共同導(dǎo)致的結(jié)果。同時(shí)也注意到, 基于COI基因序列分析得到的榮成野生群體與即墨子代之間的Fst值最大(為 0.4958), 兩群體間遺傳差異較大, 這可能會(huì)對(duì)榮成魁蚶野生群體的遺傳結(jié)構(gòu)產(chǎn)生不可忽略的影響, 底播時(shí)應(yīng)慎重。方耀林等[29]在研究野生大鯢及其人工繁殖后代的遺傳多樣性分析時(shí),發(fā)現(xiàn)2個(gè)子代種群的Fst值較大, 認(rèn)為不同大鯢人工繁殖的后代, 遺傳分化有擴(kuò)大的趨勢(shì)。這提示人們也可進(jìn)一步對(duì)人工繁育的榮成群體子代和即墨群體子代的遺傳多樣性和遺傳結(jié)構(gòu)進(jìn)行研究, 以更準(zhǔn)確地揭示魁蚶不同地理群體的遺傳多樣性和遺傳分化關(guān)系, 為魁蚶的底播增殖工作提供科學(xué)的參考依據(jù)。
4 結(jié)論
榮成市魁蚶的底播增殖正逐步深入開展。底播苗種的選用是底播增殖能否取得顯著成效的關(guān)鍵。人工增殖是一種重要的資源保護(hù)與修復(fù)手段。但選擇適當(dāng)?shù)姆N苗和增殖地點(diǎn)是前提。若選擇不當(dāng), 不僅達(dá)不到保護(hù)資源或修復(fù)資源的作用, 反而可能對(duì)種群本身的遺傳特性造成破壞。在切實(shí)開展增殖工作之前, 應(yīng)采取必要的調(diào)研等相關(guān)工作, 確保增殖活動(dòng)能取得預(yù)期的成效。
本研究通過對(duì)即墨和榮成兩地的野生群體以及人工繁育即墨群體獲得的子代進(jìn)行遺傳多樣性和遺傳結(jié)構(gòu)的分析, 探討了即墨群體人工繁育的苗種在榮成海域底播增殖的可行性。研究結(jié)果發(fā)現(xiàn), 榮成和即墨2個(gè)野生群體本身就存在中等程度的遺傳分化。人工繁育的即墨子代群體遺傳多樣性比即墨野生群體有所降低。榮成群體與即墨子代之間的遺傳差異,較之與即墨野生群體的差異更大。如果采用即墨子代在榮成底播, 可能會(huì)對(duì)榮成野生魁蚶野生群體的遺傳結(jié)構(gòu)產(chǎn)生一定影響, 因此, 采用即墨人工繁育子代作為榮成魁蚶底播增殖的苗種來源時(shí)必須十分慎重, 并在苗種繁育時(shí)盡量采用大群體有效親本。
[1]王如才, 王昭萍, 張建中. 海水貝類養(yǎng)殖學(xué)[M]. 青島: 青島海洋大學(xué)出版社, 1993: 156.
[2]郭學(xué)武. 魁蚶生殖生物學(xué)初步研究[J]. 海洋水產(chǎn)研究, 1994, 15:55-63.
[3]于瑞海, 孫志剛, 王長(zhǎng)竹, 等. 魁蚶大水體升溫育苗技術(shù)研究[J]. 海洋科學(xué), 1995, 2: 5-7.
[4]于瑞海, 王如才, 姜春麗, 等. 魁蚶生產(chǎn)性間斷升溫育苗技術(shù)的研究[J]. 黃渤海海洋, 1997, 15(3): 40-45.
[5]Numaguchi K. Gonad development of the ark shellScapharca broughtonii(Schrenck)broodstock in farming grounds of Japan [J]. Nippon Suisan Gakkaishi,1996, 62(3): 384-392.
[6]鄭國(guó)富, 王曉波, 付卓. 魁蚶人工育苗生產(chǎn)中幾項(xiàng)技術(shù)探討[J]. 河北漁業(yè), 2008, 1: 41-42.
[7]陳覺民, 王恩明, 李何. 海水中某些化學(xué)因子對(duì)魁蚶幼蟲、稚貝及成體的影響[J]. 海洋與湖沼, 1989, 20(1):15-22.
[8]Park MS, Lim HJ, Kim PJ. Effect of environmental factors on the growth, glycogen and hemoglobin content of cultured ark shell,Scapharca broughtonii[J].J Korean Fish Soc, 1998, 31: 176-185.
[9]Lee JM, Park JW, Yoo MS, et al. Morphological characteristics and genetic diversity using the RAPD technique in the ark shell,Scapharca broughtonii(Schrenck)from Korea and China [J]. J Korean Fish Soc, 1997, 30: 297-304.
[10]Yokogawa K. Morphological and genetic differences between Japanese and Chinese red ark shellScapharca broughtonii[J]. Fish Sci, 1997, 63: 332–337.
[11]喻子牛, 孔曉喻, 楊銳, 等. 魁蚶(Scapharca broughtonii)等位基因酶遺傳變異研究[J]. 青島海洋大學(xué)學(xué)報(bào), 1998, 28(1): 51-58.
[12]Cho ES, Jung CG, Sohn SG, et al. Population genetic structure of the ark shellScapharca broughtoniiSchrenck from Korea, China, and Russia based on COI gene sequences [J]. Marine Biotechnology, 2007, 9 (2):203-216.
[13]宋云杰, 郭旭強(qiáng). 榮成市魁蚶底播增殖可行性探討[J]. 齊魯漁業(yè), 2010, 27(8): 53-54.
[14]唐啟升, 王俊, 邱顯寅, 郭學(xué)武. 魁蚶底播增殖的試驗(yàn)研究[J]. 海洋水產(chǎn)研究, 1994, 15: 79-86.
[15]孫鵬飛, 劉杰, 王衛(wèi)民, 等. 淺海底播養(yǎng)殖魁蚶試驗(yàn)[J]. 河北漁業(yè), 2009, 10: 42.
[16]于瑞海, 李琪. 無公害魁蚶底播增養(yǎng)殖穩(wěn)產(chǎn)新技術(shù)[J]. 海洋湖沼通報(bào), 2009, 3: 87-90.
[17]林培振, 劉德坤, 杜萌萌, 等. 魁蚶海區(qū)底播增養(yǎng)殖技術(shù)[J]. 中國(guó)水產(chǎn), 2010, 11:42.
[18]王曉偉, 牙鲆生長(zhǎng)相關(guān)性狀遺傳力估計(jì)[D]. 青島:中國(guó)科學(xué)院海洋研究所, 2008.
[19]陳琳琳, 孔曉瑜, 周立石, 等. 魁蚶核糖體DNA基因轉(zhuǎn)錄間隔區(qū)的序列特征[J]. 中國(guó)水產(chǎn)科學(xué), 2005,12(1): 104-108.
[20]Tamura K, Dudley J, Nei M, et al. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA)Software Version 4.0 [J]. Molecular Biology and Evolution, 2007,24 (8): 1596-1599.
[21]Kimura M, Yanagimoto T, Munehara H. Maternal identification of hybrid eggs inHexagrammosspp. by means of multiplex amplified product length polymorphism of mitochondrial DNA [J]. Aquatic Biology, 2007:187-192.
[22]Librado P, Rozas J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data [J].Bioinformatics, 2009, 25 (11): 1451-1452.
[23]Excoffier L, Laval G, Schneider S. Arlequin ver. 3.0:An integrated software package for population genetics data analysis. Evolutionary Bioinformatics Online 1,2005, 47-50.
[24]Rice W. Analyzing tables of statistical tests [J].Evolution, 1989, 43: 223-225.
[25]鄭文娟, 朱世華, 沈錫權(quán), 等. 基于線粒體 COI基因序列探討泥蚶的遺傳分化[J]. 動(dòng)物學(xué)研究, 2009,30(1): 17-23.
[26]陳麗梅, 孔曉瑜, 喻子牛, 等. 3種蟶類線粒體 16S rRNA和COI基因片段的序列比較及其系統(tǒng)學(xué)初步研究[J]. 海洋科學(xué), 2005, 29(8): 27-32.
[27]陳琳琳. 幾種雙殼貝類ITS區(qū)序列分析與遺傳多樣性研究[D]. 青島: 中國(guó)海洋大學(xué), 2004.
[28]許志強(qiáng), 葛家春, 李曉輝, 等. 基于rDNA ITS序列研究蚌科 6種類的系統(tǒng)發(fā)生關(guān)系[J]. 淡水漁業(yè), 2009,39(1): 16-20.
[29]方耀林, 張燕, 肖漢兵, 等. 野生大鯢及其人工繁殖后代的遺傳多樣性分析[J]. 水生生物學(xué)學(xué)報(bào), 2008,32(5): 783-786.
[30]王鴻霞, 吳長(zhǎng)功, 相建海. 凡納濱對(duì)蝦繁殖中不同親本對(duì)子代遺傳貢獻(xiàn)率的差異[J]. 動(dòng)物學(xué)報(bào), 2006,52(1): 175-181.
[31]Shikano T, Chiyokubo T, Taniguchi N. Effect of inbreeding on salinity tolerance in the guppy (Poeciliareticulata)[J]. Aquaculture, 2001, 202: 45-55.
[32]Sekino M, Saitoh K, Yamada T, et al. Microsatellite-based pedigree tracing in a Japanese flounderParalichthys olivaceushatchery strain: implications for hatchery management related to stock enhancement program [J].Aquaculture, 2003, 221: 255-263.
[33]Vandeputte M, Kocour M, Maugera S,et al.Heritability estimates for growth-related traits using microsatellite parentage assignment in juvenile common carp (Cyprinus carpioL.)[J]. Aquaculture, 2004, 335: 223-236.
[34]Brown W M. Evolution of Genes and Proteins [M].Sunderland: Sinauer, 1983, 62-88.
[35]王慶, 傅洪拓, 核rDNA ITS區(qū)序列在水產(chǎn)動(dòng)物研究中的應(yīng)用[J]. 安徽農(nóng)學(xué)通報(bào), 2007, 13(11): 99-101.
[36]Philips R B, Sajdak S L, Domanico M J. Relationships among charrs bases on DNA Sequences [J]. Nord J Frdabw Res, 1995, 71: 378-391.
[37]林昕, 王鵬, 杜琦, 李振華, 等. 福建沿海不同養(yǎng)殖區(qū)泥蚶的 ITS-1基因片段序列分析[J]. 福建水產(chǎn),2008, 1: 61-65.
[38]Wright S. The genetical structure of populations [J].Annals of Eugenics, 1951, 15: 323-354.
[39]Slatkin M, Hudson R R. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations [J]. Genetics, 1991, 129: 555-562.
