紫扇貝、海灣扇貝及其正反雜交子代群體遺傳結構的AFLP分析
2013-10-13 08:14:34黃曉婷胡麗萍王春德包振民
海洋科學 2013年8期
孫 妍, 黃曉婷, 胡麗萍, 王 師, 王春德, 包振民
(1.海洋生物遺傳學與育種教育部重點實驗室, 中國海洋大學 海洋生命學院, 山東 青島 266003; 2.青島農業大學 海洋科學與工程學院, 山東 青島266109)
海灣扇貝(Argopecten irradians, Lamarck, 1819)原產于北美洲大西洋沿岸, 于 1982年引入中國[1],由于其生長迅速而受到養殖戶歡迎, 目前已成為中國重要的海水養殖貝類之一[2-3]。但是, 由于引種個體數目較少, 隨著養殖規模的不斷擴大, 遺傳多樣性降低, 近交衰退現象顯著, 種質資源的退化需要引種復壯和遺傳育種工作的不斷進行, 對產業的可持續發展造成了嚴重威脅[4-5]。與海灣扇貝同屬的紫扇貝(A. purpuratus,Lamarck, 1819), 是原產于南太平洋的速生型中型扇貝, 主要在智力北部和秘魯沿岸進行規模化人工養殖[6], 具有體型大、生長速度快、壽命長等優點。
雜交在改善海洋貝類種質資源中有許多成功的應用[3,7], 通過雜交育種的方式, 有望豐富現有扇貝種質資源并產生優于親本生產性狀的雜種優勢。青島農業大學王春德教授于2008年將紫扇貝引入中國,并開展了紫扇貝與海灣扇貝的雜交育種工作, 獲得了雜交子一代成體[8], 在生長率、成活率上優于親本,與海灣扇貝相比, 雜交子代成貝總質量增加125.9%~138.9%, 肉柱重增加 145.4%~156.2%, 具有明顯的雜種優勢[9]。在紫扇貝和海灣扇貝的雜交中,親本雙方在雜交育種中的貢獻率大小、種間雜交子代與種內交配子代遺傳上的差異、以及雜種優勢的遺傳基礎等問題都需要我們對雜種及其親本的遺傳結構進行分析。
本研究應用AFLP分子標記技術, 對紫扇貝、海灣扇貝、正交子一代(紫扇貝♀×海灣扇貝♂)及反交子一代(海灣扇貝♀×紫扇貝♂)群體進行了遺傳結構分析, 從分子水平探討雜交子代的遺傳組成, 同時為雜種優勢機理的闡述提供理論依據。
1 材料和方法
1.1 材料
本實驗所用紫扇貝(PP)、海灣扇貝(II)及其正反雜交后代(紫扇貝♀×海灣扇貝♂, PI; 海灣扇貝♀×紫扇貝♂, IP)均由青島農業大學王春德教授提供,其中紫扇貝為秘魯引進種, 海灣扇貝取自青島膠南市, 雜交實驗及雜種培育參見Wang等[9]的方法。
1.2 方法
基因組 DNA的提取采用常規酚-氯仿抽提法提取基因組DNA[10]。AFLP分析參照Vos等[11]的方法,引物及接頭序列參照潘潔等[12], 由上海生物工程合成, 序列見表1。選擇性擴增用4種EcoR I引物和3種Mse I引物, 共5對組合引物組合: E-AAC/M- CAG、E-AAC/M-CCA、E-AAG/M-CCA、E-ACA/M- CTG、E-ATC/M-CTG。擴增產物經4.5%聚丙烯酰胺凝膠電泳后, 進行銀染顯帶。

表1 AFLP分析中所用的接頭及引物序列Tab. 1 The adapter and primer sequence used in AFLP analysis
數據統計與分析銀染之后得到的帶譜應用Cross checker 2.8軟件進行統計分析, 有帶記為1, 無帶記為 0, 獲得 1、0矩陣。統計各群體擴增位點數和多態位點數。用Pop Gene32軟件計算群體內的多態位點比率、Nei’ s基因多樣性、香農氏多樣性指數和群體間遺傳相似度及遺傳距離。為評估遺傳變異來源于群體內還是群體間, 用Arlequin3.11進行分子方差分析(Analysis of Molecularof Variance, AMOVA), 計算群體間變異來源和遺傳分化指數。
2 實驗結果
2.1 AFLP 擴增帶譜分析
本研究用 E-AAC/M-CAG、E-AAC/M-CCA、E-AAG/M-CCA、E-ACA/M-CTG、E-ATC/M-CTG五對引物組合, 在紫扇貝(PP)、海灣扇貝(II)及其正反雜交子代(PI和IP)4個群體中共得到了433個位點,平均每對引物得到86.6個位點。
AFLP在親本和雜交后代 4個群體中的擴增帶譜可以分為4種, 如圖1: (I)雙親和正反雜交子代均有的擴增位點, 這種位點共有 169個, 占總位點數的 39.03%; (II)親本一方與雜交子代共享位點,其中, 紫扇貝與正反交子代共享的位點數為 82個,海灣扇貝與雜交子代共享的位點數為 181個; (III)在親本中出現, 而雜交子代中丟失的位點, 統計發現, 紫扇貝和海灣扇貝各有30和18個位點在子代中丟失; (Ⅳ)非親位點, 即親本中沒有, 而雜交子代中出現的位點, 這類位點共有 12個(正交子代中5個、反交子代中 8個)。由此可見, 正反交子代97.23%擴增得到的位點來源于紫扇貝和海灣扇貝,且來源于海灣扇貝的位點數多于來源于紫扇貝的位點數。本研究在分子水平上證實了雜交扇貝成貝的雜種身份。

圖1 紫扇貝、海灣扇貝及雜交子代AFLP選擇性擴增結果—E33M54Fig. 1 Polyacrylamide gel electrophoresis result of selective amplification
2.2 紫扇貝(PP)、海灣扇貝(II)及其正反雜交子一代(PI和IP)的群體遺傳結構分析
4個群體的多態位點比率、Nei’ s基因多樣性、香農氏指數見表 2。4個群體中, 紫扇貝(PP)群體的遺傳多樣性最低; 海灣扇貝(II)群體最高。雜交子代群體的遺傳多樣性介于兩親本之間, 且反交群體高于正交群體。紫扇貝群體內遺傳多樣性較低可能是由于目前引種數目較少引起的。

表2 4個群體內遺傳多樣性參數Tab. 2 Genetic diversityparameters withinfour groups
從群體間的遺傳距離(表 3)可以看出, 兩個親本(PP和II)間的遺傳距離為0.5464, 正反雜交F1代與兩親本的遺傳距離為 0.2186~0.2701, 表明兩個親本間、子代與親本間的遺傳差異較大。而正反交F1代間遺傳距離小, 遺傳相似度高, 表明正反交子代無群體分化。此外, 正反雜交F1代與母本的遺傳距離均比與父本的遺傳距離小, 表明雜交子代一定程度偏向母本, 從母本獲得的遺傳信息可能要多于從父本獲得的遺傳信息。
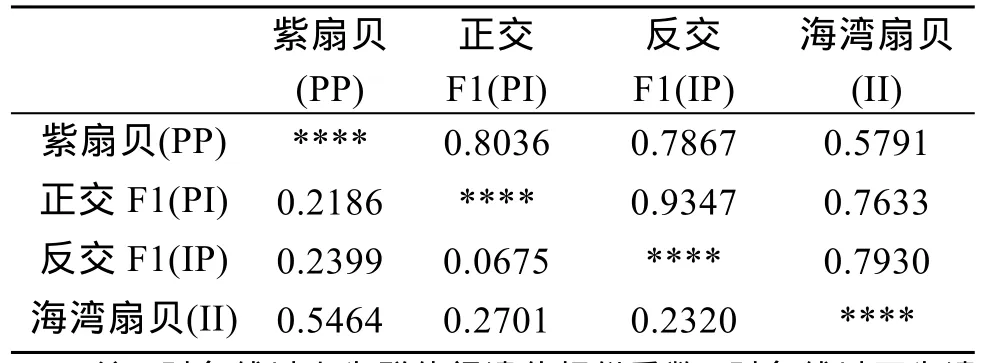
表3 4個群體間的遺傳相似性和遺傳距離Tab. 3 Genetic distances and similarities between four groups
應用分子方差分析(AMOVA)對4個群體兩兩間的遺傳多樣性來源進行分析(表 4), 兩親本群體間遺傳差異最大, 兩親本間和親本與雜交子代間遺傳分化顯著; 正反雜交子代間遺傳差異最小, 沒有明顯的遺傳分化。
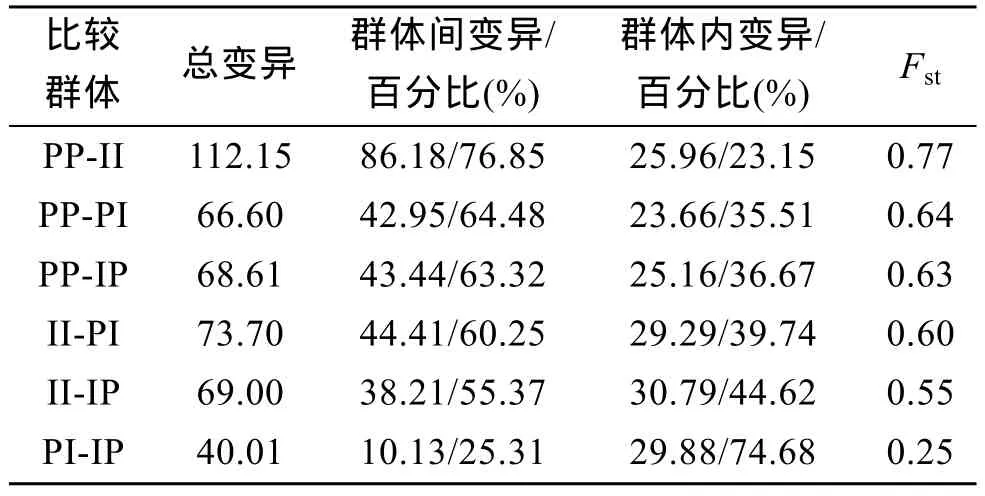
表4 4個群體總遺傳變異來源及遺傳分化指數Tab. 4 The source of variation and Fstabout four scallop groups
3 討論
紫扇貝與海灣扇貝同屬于海灣扇貝屬(Argopecten), 進化關系較近, 是當今世界上兩種重要的海水經濟養殖扇貝[13]。雖然紫扇貝在生產上做出了相當大的貢獻, 但針對其分子生物學的研究很少, 基因組信息相對于其他經濟養殖扇貝而言十分有限。AFLP標記對基因組的信息背景沒有要求, 通過改變限制性內切酶和選擇性堿基的種類與數目,可產生大量的理論上可以覆蓋整個基因組標記數目,該標記還具有豐富的多態性, 對遺傳關系相近的材料也可達到較強的多態分辨能力。本研究選用AFLP分子標記技術, 能夠從基因組的水平對紫扇貝、海灣扇貝及其正反雜交F1代4個群體之間的遺傳關系進行分析, 從而確證雜交子代的遺傳組成, 并為研究雜種優勢產生提供可能的遺傳基礎。
雜交作為一種改良生產性狀的重要途徑, 在海洋養殖扇貝的育種工作已有很多報道。扇貝種內不同地理種群之間雜交的研究較多也較為成熟, 常亞青和劉小林對櫛孔扇貝中國種群與日本種群及與俄羅斯群體的雜交組合的研究表明, 雜交后代表現出了不同程度的雜交優勢[14-15]。種間雜交方面, 國內在櫛孔扇貝(Chlamys farreri)、海灣扇貝(A. irradians irradians)、蝦夷扇貝(Patinopecten yessoensis)和華貴櫛孔扇貝(Mimachlamys nobilis)等不同種間雜交已經開展了很多工作[16-18]。但是, 種間雜交由于親本間遺傳差異較大, 常出現雜交不親和、幼體不成活等“遠緣雜交綜合征”, 導致在許多貝類的種間雜交中出現低幼蟲孵化率、幼蟲低成活率和高胚胎畸形率的現象[17,19]。僅在楊愛國等開展的櫛孔扇貝與蝦夷扇貝雜交研究中, 獲得的雜交子一代, 并在抗逆性能方面明顯優于親貝, 而通過細胞遺傳學分析發現, 該兩種扇貝的雜種身份在幼蟲時期是可以證實的, 在幼蟲發育過程中染色體組成上發生了一定的變異,成體時期的雜種身份仍需驗證[20-21]。王春德等[8-9]于2007和2008年將紫扇貝引進中國, 并開展了紫扇貝和海灣扇貝種間雜交研究, 成功獲得了雜交扇貝成體, 且雜交扇貝表現出一定的雜種優勢, 這在已有的扇貝雜交育種研究當中是鮮有報道的, 借助細胞遺傳學和分子生物學等技術手段, 作者已經證明雜種子代的確由雙親的遺傳物質組成, 證明的雜交一代的雜種身份[22]。
為了進一步了解雜種子代的遺傳組成, 本研究應用 AFLP技術對紫扇貝、海灣扇貝及正反雜交子代的遺傳結構進行了分析, 計算出兩親本遺傳距離0.5464, 遺傳分化指數Fst為0.77。Nei[23]估計亞種間的遺傳距離值約為 0.02~0.20; 種間遺傳距離的變幅為 0.10~2.00, Wright[24]提出,Fst>0.25 說明群體間的遺傳分化很大。由此可見, 本研究中兩親本扇貝群體遺傳分化顯著, 屬于兩個不同的物種, 它們的雜交屬于種間雜交。但是扇貝種間雜交子代, 尤其是雜交成貝, 可能有復雜的遺傳組成。本研究對雜交子代遺傳結構的分析表明, 正反交子代中的位點大部分來源于紫扇貝和海灣扇貝, 來源于海灣扇貝的擴增位點略多于來源于紫扇貝的擴增位點, 原因可能是海灣扇貝群體的遺傳多樣性高于紫扇貝; 此外,正反交F1與雙親的Fst指數都>0.25, 表明分化顯著,且F1與雙親的遺傳相似度和遺傳距離顯示其為介于雙親間的中間種; 雖然雜交子代與紫扇貝和海灣扇貝的遺傳關系上并不完全對等, 略偏向于母本一方,但并不表現出明顯的傾向性。本研究結果為紫扇貝和海灣扇貝種間雜交子代的遺傳組成提供了分子水平的證據。
紫扇貝和海灣扇貝雜交獲得的子代, 與海灣扇貝相比, 雜交子代成貝總質量增加了 125.9%~138.9%,肉柱質量增加了 145.4%~156.2%, 具有明顯的雜種優勢[9]。雜種優勢作為生物界普遍存在的現象, 在改良生產性狀和種質品質中得到了廣泛的應用, 但其理論基礎的研究卻相對薄弱, 理論研究的滯后也制約其在生產中的進一步應用。因此, 雜種優勢的遺傳機理研究一直是近百年來遺傳和育種學家研究的熱點和難點。早在 20世紀初, Bruce[25]和 Shull[26]就基于單基因遺傳效應相繼提出了雜種優勢的顯性假說和超顯性假說。隨著研究手段的改進和研究內容的深入, 認為雜種優勢的遺傳實質應是各種基因效應的綜合,特別是上位效應和連鎖效應在雜種優勢遺傳基礎中將占有重要地位。雜種優勢對包括貝類在內的海洋經濟動物進行遺傳改良研究和應用的歷史較短, 關于其雜種優勢遺傳機理的研究報道也很少[27]。AFLP作為一種顯性標記應用于紫海雜交貝的遺傳結構分析, 可以為海洋貝類雜種優勢產生的機理的探討提供一定的線索。在雜交過程中子代出現了非親遺傳的位點, 這些位點的產生可能是由于親本雙方的同源染色體相互作用對酶切位點引起的修飾或隱蔽, 或者個別酶切位點附近堿基突變等, 這些雜交子代中出現的種種非孟德爾遺傳變化可能與其在生產上表現出的雜種優勢有一定關系, 需要進一步深入研究加以證實。
[1]張福綏, 何義朝, 劉祥生, 等. 海灣扇貝(Argopecten irradians)引種、育苗及試養[J].海洋與湖沼, 1986,17(5): 367-374.
[2]張福綏, 何義朝, 楊紅生. 海灣扇貝引種工程及其綜合效應[J]. 中國工程科學, 2000, 2(2): 30-35.
[3]Guo X. Use and exchange of genetic resources in molluscan aquaculture [J]. Reviews in Aquaculture,2009, 1(3-4): 251-259.
[4]張國范, 劉述錫, 劉曉, 等. 海灣扇貝自交家系的建立和自交效應[J]. 中國水產科學, 2003, 10(6):441-445.
[5]Zheng H, Zhang G, Liu X, et al.Inbreeding depression for various traits in two cultured populations of the American bay scallop,Argopecten irradians irradiansLamarek (1819)introduced into China[J].Journal of Experimental Marine Biology and Ecology,2008,364(1):42-47.
[6]Brand E, Merino G E, Abarca A, et al. Scallop fishery and aquaculture in Chile [J]. Developments in Aquaculture and Fisheries Science, 2006, 35: 1293-1314.
[7]燕敬平, 孫慧玲, 方建光, 等. 日本盤鮑與皺紋盤鮑雜交育種技術研究[J]. 海洋水產研究, 1999, 20(1):35-39.
[8]王春德, 劉保忠, 李繼強, 等. 紫扇貝與海灣扇貝種間雜交的研究[J]. 海洋科學, 2009, 33(10): 84-87.
[9]Wang C, Liu B, Li J, et al. Introduction of the Peruvian scallop and its hybridization with the bay scallop in China[J]. Aquaculture, 2011, 310(3): 380-387.
[10]Sambrook J, Russell David W. Molecular cloning: a laboratory manual. Vol. 3[M]. Cold Spring Harbor Laboratory Press, 1989.
[11]Vos P, Hogers R, Bleeker M, et al. AFLP: a new technique for DNA fingerprinting[J]. Nucleic acids research, 1995, 23(21): 4407-4414.
[12]潘潔, 包振民, 趙洋, 等. 扇貝 AFLP分析中的引物選擇[J]. 青島海洋大學學報, 2002, 32(Sup.): 310.
[13]Gonzalez M L, Perez M C, Lopez D A. Breeding cycle of the northern scallop,Argopecten purpuratus(Lamarck,1819)in southern Chile [J]. Aquaculture Research, 2002,33(11): 847-852.
[14]常亞青, 劉小林, 相建海, 等.櫛孔扇貝中國種群與日本種群雜交子一代的早期生長發育[J]. 水產學報,2002, 26(5): 385-388.
[15]劉小林, 常亞青, 相建海, 等. 櫛孔扇貝不同種群雜交效果的初步研究.I中國種群與俄羅斯種群的雜交[J]. 海洋學報, 2003, 25(1): 93-99.
[16]張玉勇, 常亞青, 宋堅. 雜交育種技術在海水養殖貝類中的應用及研究進展[J]. 水產科學, 2005, 24(4):39-41.
[17]劉憲杰, 常亞青. 扇貝種間雜交的可行性及幼體早期發育的初步研究[J]. 大連水產學院學報, 2006, 4: 35-39.
[18]劉廣緒, 包振民, 胡景杰, 等. 櫛孔扇貝、華貴櫛孔扇貝及其種間雜交子代、種內交配子代的 ISSR分析[J]. 中國海洋大學學報, 2006, 36(1): 71-75.
[19]張國范, 劉曉. 關于貝類遺傳改良幾個問題的討論[J].水產學報, 2006, 30(1): 130-137.
[20]呂振明, 楊愛國, 王清印, 等. 櫛孔扇貝和蝦夷扇貝雜交子代的細胞遺傳學研究[J].高技術通訊, 2006,16(8): 853-858.
[21]呂振明, 楊愛國, 王清印, 等. 櫛孔扇貝和蝦夷扇貝雜交子代的GISH鑒定及其免疫學特性[J]. 中國水產科學, 2006, 13(4): 587-602.
[22]Hu L, Huang X, Mao J, et al. Genomic Characterization of Interspecific Hybrids between the ScallopsArgopecten purpuratusandA. irradians irradians[J]. PLOS ONE, 2013, 8(4): e62432.
[23]郭平仲. 種群間遺傳距離與進化趨異[J]. 北京師范學院學報. 1992, 13(2): 63-71.
[24]Weight S. Evolution and the Genetics of Population Variability Within and Among Natural Population[M].Chicago: University of Chicago Press, 1978.
[25]Shull G H. The composition of a field of maize[J].Journal of Heredity, 1908(1): 296-301.
[26]蘭進好, 張寶石, 周鴻飛. 作物雜種優勢遺傳基礎研究進展[J]. 中國農學通報2005, 21(1): 114-119.
[27]張國范, 劉曉, 闕華勇, 等. 貝類雜交及雜種優勢理論和技術研究進展[J]. 海洋科學2004, 28(7): 54-60.
