前置庫凈化系統中水生植物的選擇
2013-08-17 03:50:32高松峰楊倩琪
水土保持研究 2013年6期
高松峰,楊倩琪
(1.江蘇省環境科學研究院,南京210036;2.南京理工大學,南京210094)
隨著我國對飲用水水源地保護的日益重視,水源地周邊的點源污染得到了有效的遏制,但面源污染的影響比例和危害越來越突出,特別是以水庫型水源地為主的中小城鎮,其周邊的農村生活污水、稻田排水等農業面源污染通過降雨或地表徑流的方式對水源地水質造成嚴重的危害[1-2]。該類污染的特點主要體現在面源污染具有突發性強、污染物種類復雜多變、污染負荷變化大,難采取單一集中處理等特點。初期來水,污染物濃度高,負荷大,需要凈化處理,后期來水水量較大,污染程度低可直接排放進入水源地,前置庫系統蓄混放清的特點對水源地面源污染防控起著重要作用[3-4]。通過在水源地水庫前設置前置庫,前置庫排水廊道內設置植生帶,沉水(浮)水植生帶,利用這些植物對初期來水中的有機物、氮鹽及磷酸鹽等營養鹽物質,通過截留、沉降、吸附、降解等方式凈化后排入水源地水庫[5-7]。
水生植物是水體中的初級生產者,也是前置庫水生生態系統中物質與能量流的主要傳遞者,其種類和種群數量變動對庫區生態及水域環境有著重大影響,氮和磷的吸收主要受濕地植物生物量的影響[8]。Gumbricht等[9]認為在不收割的情況下,水生植物主要通過促進微生物的硝化與反硝化作用達到去除N的效果,而在收割的情況下,或在低溫時期,對氮和對磷的去除則主要是通過主動吸收,但傳統型前置庫的氮磷凈化效果欠佳,尤其冬季低溫條件下銨態氮(NH+4-N)、總氮(TN)去除速率遠低于夏季,僅為夏季的1/4左右[10-12]。水生植物體內的氮、磷含量受到水體營養狀況和植物生長狀況的影響,而且由于沉水植物根、莖、葉都沉沒于水中,水質對植株營養鹽的影響更為敏感。在一些營養鹽濃度較低的天然湖泊中,氮和磷成為植物生長的限制性因素[13-14]。國內外研究人員已相繼篩選出一批能高效去除水中各種污染物質的植物,一些漂浮植物如鳳眼蓮、浮萍等已大量用于氧化塘等工程中,蘆葦等則多用于人工濕地,但這些應用多著重植物根部微生物功能的發揮,主要靠植物自身的吸收功能來進行凈化水體的應用還較少,同時并未根據前置庫內不同分區的水質情況,確定水生植物的種群結構,特別是冬季低溫條件下植物種群,從而提高水中氮磷等營養鹽的去除效果[15-18]。因此,本文結合溧陽市塘馬水庫水質安全保障工程背景及水質特征,從技術集成角度出發,因地制宜地構建復合型前置庫凈化系統,在實驗室尺度下探討確定不同植物組合凈化效果及污染物去除規律,確定復合型前置庫去處污染物最優的工況參數,為前置庫凈化系統在溧陽市塘馬水庫水質安全保障工程中應用提供科學依據。
1 實驗材料與方法
1.1 夏秋季植物篩選試驗
水生植物包括挺水植物、漂浮植物、浮葉植物、沉水植物等。結合示范工程開展地區的氣候、水文、植被等狀況和室內試驗結果,選擇挺水植物:香蒲、千屈菜、水蔥、蓮藕、水芹、菖蒲、空心菜、西伯利亞鳶尾和沉水植物:伊樂藻、菹草、輪葉黑藻、苦草、狐尾藻,作為前置庫的備選植物,部分為冬季低溫條件下生長水生植物。本實驗采用靜態實驗方案在夏秋季節進行。試驗裝置為容量為40L的塑料桶,上部直徑(內徑)為39cm,底部直徑(內徑)為31cm。桶中栽種植物,桶底鋪一層洗凈的石英砂,以滿足挺水植物固根要求。實驗從8月中旬開始到九月底結束。植物栽種10d后,每隔3d定期取水樣測定其中的氮磷營養鹽含量,計算氮磷的去除率。定期采用自來水補充水量,以彌補因蒸發、植物吸收、取樣造成的水量損失。
實驗用水采用人工配水,初始水質指標為:TN 1.67mg/L、TP 0.41mg/L、CODMn6.75mg/L。根據《地表水環境質量標準基本項目標準限值》(GB3838-2002),實驗用水屬Ⅳ~Ⅴ類。
1.2 冬季植物篩選實驗
在冬季,大部分植物已經進入凋落期,對水質凈化作用不明顯,選擇合適的適宜于冬季生長的植物并研究其對水質的凈化規律是保證前置庫冬季長效運行的重要工作。以上述夏秋季植物凈化效果的研究為基礎,選用蘇南地區常見的能夠耐受冬季低溫的水生植物進行冬季生長試驗以及凈化效果研究。挺水植物:水芹、西伯利亞鳶尾、香蒲;沉水植物:伊樂藻、菹草以及狐尾藻。
(1)冬季植物的生長規律。試驗選擇在冬季進行,試驗時間為45d,自然條件下水溫的波動范圍為0~5℃。生長的水質條件采用人工配水方式,試驗為L9(33)的正交試驗。本試驗采用氮磷比、氮濃度、氮形態來表征不同富營養化水體的特征,以上每個因素各設計3個水平。氮形態的水平為硝態氮,混合態氮(銨態氮和硝態氮各占50%)和銨態氮,各處理組的水質情況見表1。
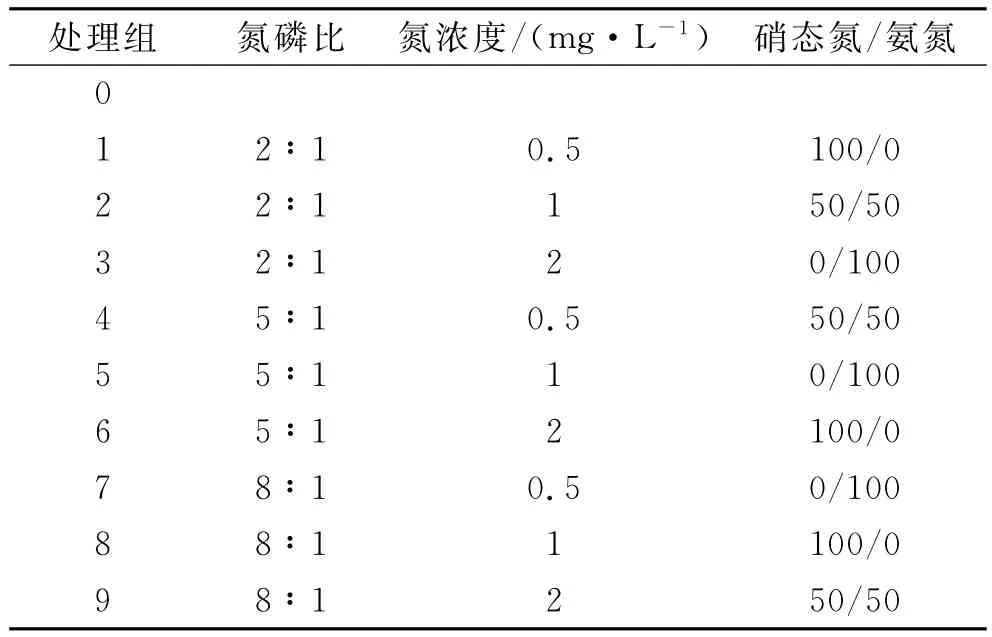
表1 不同處理組的水質參數
試驗裝置為容量為40L的塑料桶,上部直徑(內徑)為39cm,底部直徑(內徑)為31cm。試驗開始時,把植物洗凈稱重,0—9號桶中栽種植物,0號桶中不加營養液,底部鋪設洗凈的石英砂用于固定挺水植物為對照樣。試驗用氫氧化鈉溶液和稀硫酸調節pH值,pH值控制在7,確保營養鹽濃度變化范圍不超過20%,采用相對生長率表征植物生長情況:
相對生長率R=(Wt-W0)/W0(1)式中:R——相對生長率;Wt——試驗結束后生物量(濕重);W0——試驗前生物量(濕重)。
(2)冬季植物對水質的凈化效果。實驗用水采用人工配水,初始水質指標為:TN 1.67mg/L、TP 0.41mg/L、CODMn6.75mg/L。根據《地表水環境質量標準基本項目標準限值》(GB3838-2002),實驗用水屬Ⅳ—Ⅴ類。
采用靜態試驗,將植物栽種在盛有一定量自來水的大型塑料桶(挺水植物用洗凈的石英砂固定),容量為40L的塑料桶,上部直徑(內徑)為39cm,底部直徑(內徑)為31cm。試驗用水體積30L,植物質量約100g。所有試驗桶均放在室外自然光照的地方,但要避免雨淋。試驗期間記錄每天的水溫變化情況,水溫是取3個時間點的平均值(8:00,12:00,20:00),采樣前要按照每個試驗組的蒸發量先補充蒸餾水。根據《水和廢水監測分析方法(第四版)》[19]測定SS、TN、NH4-N、COD、TP、TDP 指標。其中 NH4-N、NO3-N、TDP為過0.45μm醋酸纖維濾膜后測定。
2 結果與分析
2.1 夏秋季植物對水質的作用
挺水植物對總氮的去除率為水芹>香蒲>水蔥,去除效果較好,平均去除率達到了26%以上;對氨氮的去除率為水芹> 水蔥>香蒲,去除效果較好,平均去除率達到了30%以上;對總磷的去除率為水芹>菖蒲>西伯利亞鳶尾,去除效果較好,平均去除率達到了30%以上(圖1);沉水植物中對總氮和氨氮去除率大小次序為伊樂藻>狐尾藻>輪葉黑藻,去除效果較好,平均去除率達到了20%以上;對總磷的去除率為伊樂藻>狐尾藻>輪葉黑藻,去除效果較好,平均去除率達到了25%以上(圖2)。

圖1 夏秋季節挺水植物對TP的去除率

圖2 夏秋季節沉水植物對TP的去除率
初期,植物對NH+4-N的去除效果較TN好,這主要是因為水體中的NH+4-N較少部分通過植物吸收和揮發作用而去除,大部分則是通過硝化作用和反硝化作用的連續反應而去除,這種反應過程會增加水體中NO-3-N的量,從而使TN的降解幅度變小。但隨著NH+4-N濃度下降并趨于穩定后,植物開始以吸收NO-3-N為主,從而不斷降低水體TN含量,導致后期TN的去除率逐漸高于NH+4-N。
植物對磷的去除率表現出先升高,后降低的現象。對磷的去除一方面是以磷酸鹽沉降并固定在基質上,另一方面是可溶性磷被植物吸收。在試驗初期對磷的去除效果上升非常顯著,這是因為磷被底部的石英砂吸附而使得其濃度大幅度降低,但也有部分磷會逐漸從石英砂中釋放出來,造成后期去除率出現下降的現象。綜上所述,在夏秋季節可以選擇的挺水植物主要為水芹、西伯利亞鳶尾、香蒲;沉水植物主要有伊樂藻、輪葉黑藻、狐尾藻。
2.2 冬季水生植物生長
試驗周期內植物的冬季相對生長率如圖3和4所示。從圖3中可以看出,在相同的水質及溫度條件下,挺水植物中水芹相對生長率為48.5%~92.2%生長狀況最好,香蒲相對生長率為10.2%~31.5%生長狀況最差;試驗期間,水溫在0~5℃,伊樂藻在低溫季節仍然繼續生長。由圖4可見,伊樂藻相對生長率為43%~94.2%,而在同樣的試驗條件下水溫在0~5℃,菹草普遍生長緩慢,相對生長率為-23.9%~39.6%,因此沉水植物中伊樂藻的生長狀況最好,狐尾藻的生長狀況最差。
通過方差分析(表2),發現氮磷比影響植株含磷量的顯著性因素,影響植物相對生長率的因素主次順序為:氮磷比>氮形態>氮濃度。氮磷比為2∶1與8∶1之間存在顯著差異(P<0.05),不同水平的氮濃度與氮形態沒有對伊樂藻相對增長率產生顯著影響(P分別為0.320,0.077),氮磷比為8∶1時植物的相對生長率最大,氮濃度越高,相對生長率越大。混合態氮最能促進伊樂藻的生長,其次為硝態氮,供應混合態氮與銨態氮的處理組間存在顯著差異(P<0.05)。由表3知,氮磷比、氮濃度、氮形態都不是菹草相對生長率的主要影響因素,各因素不同水平之間也不存在顯著差異,說明了雖然菹草屬于低溫生態位,但其最適生長溫度范圍為10~15℃,耐低溫能力沒有伊樂藻強,在0~5℃條件下,菹草基本不生長。
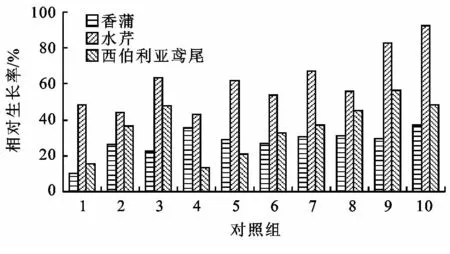
圖3 挺水植物冬季相對生長率對比
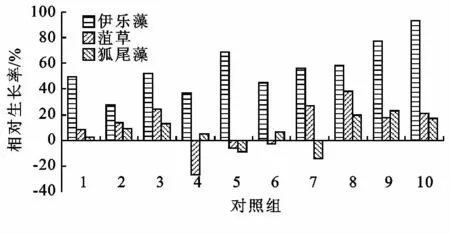
圖4 沉水植物冬季相對生長率對比

表2 挺水植物植株相對增長率顯著性分析
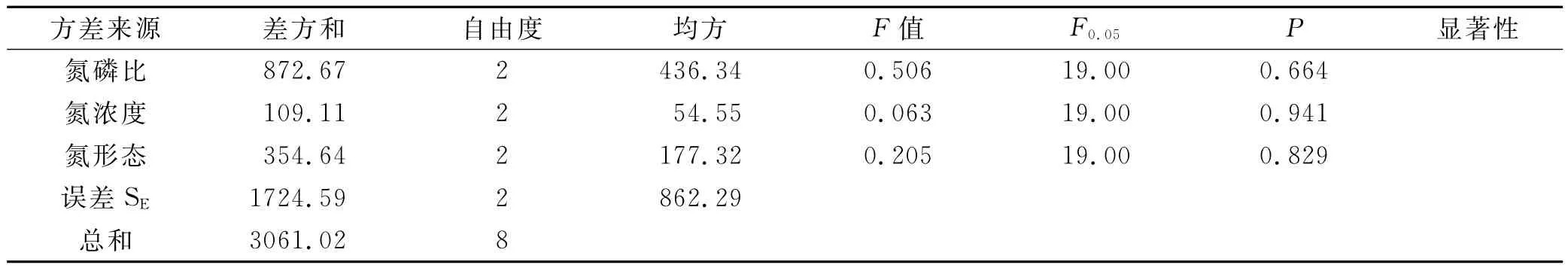
表3 沉水植物植株相對增長率顯著性分析
2.3 冬季水生植物對水質的作用
冬季水生植物對營養鹽的去除與其生長狀況有著較為密切的聯系,在保證沉水植物良好生長的前提下,冬季挺水植物中對總氮和總磷去除率大小順序分別為:水芹>西伯利亞鳶尾>香蒲,香蒲>水芹>西伯利亞鳶尾,去除效果較好;沉水植物對總氮和總磷去除率大小順序均為伊樂藻>菹草>狐尾藻,去除效果較好(圖5—6)。沉水植物對污染水體產生的主要效應是降低濁度,穩定水質,促進水中營養鹽的降低。這種改善效應主要是通過植物自身對污染物質的吸收和增強水體的自凈能力來實現的。沉水植物由于其生境結構的特殊性,其根、莖、葉都能從水中吸收營養鹽。伊樂藻對水中氮的吸收主要通過生物量的增長與富集作用,對磷的吸收主要通過富集作用;菹草對水中氮的吸收主要通過生物量的增長,對磷的吸收主要通過富集作用。沉水植物對營養鹽的去除效果伊樂藻最好,結合植物的生長狀況與對營養鹽的去除效果,可以在冬季選擇水芹作為前置庫的凈化挺水植物,伊樂藻作為前置庫凈化的沉水植物。
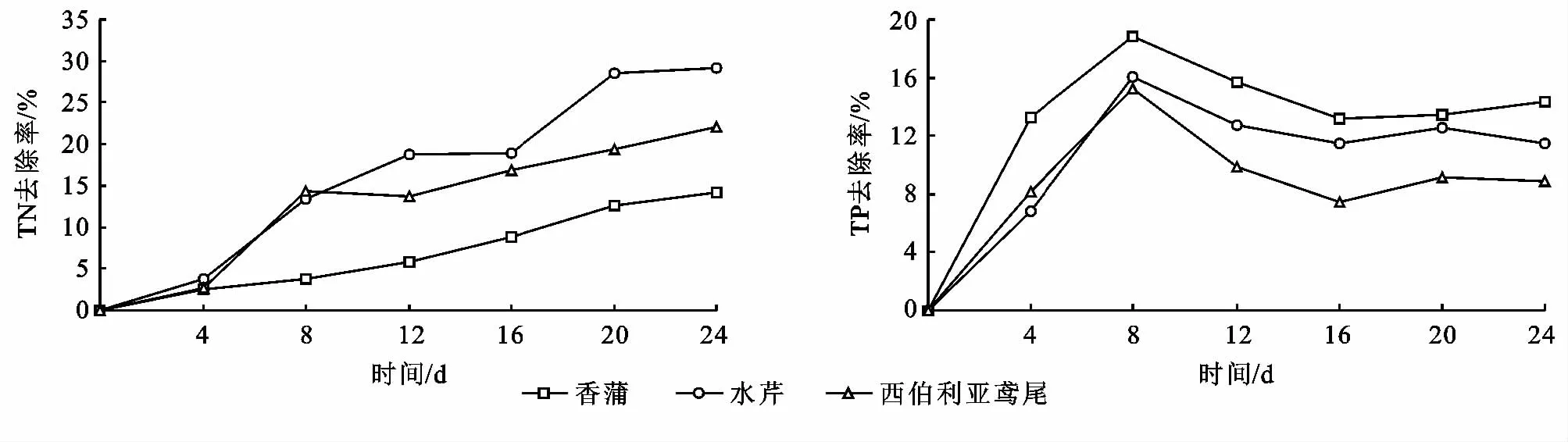
圖5 冬季挺水植物對TN、TP的去除率
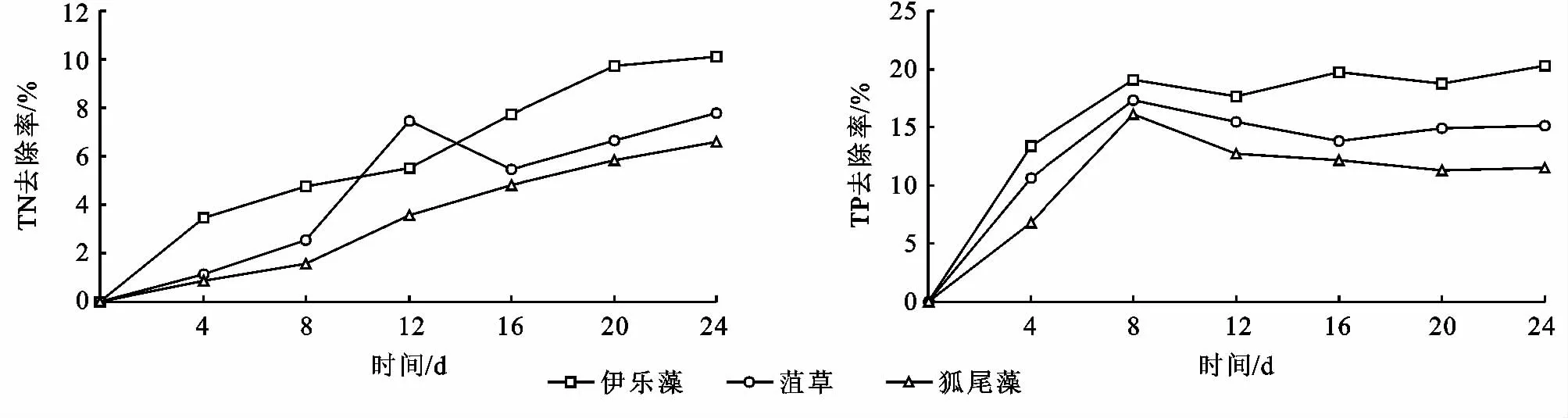
圖6 冬季沉水植物對TN、TP的去除率
2.4 伊樂藻對其他水質指標的作用
確定沉水植物伊樂藻,研究其對其他水質指標的影響,伊樂藻生長要求水深小于2~2.2倍水體透明度,最適溫度為25℃左右。在水溫為5~30℃都能處于正常的營養生長狀態,當溫度低于5℃或高于30℃時,其生長就會受到抑制。由圖7可知,有植物的池子,濁度在短期內下降效果明顯,但在試驗后期,兩者的差別減少,這是因為經過長時間的靜置,對照樣中的懸浮物大部分能沉淀到底泥中。在第43小時,兩個池子的濁度有較大幅度的上升是由于當天風力很大,水體受到擾動,對照池的濁度較第19小時增加了28NTU,但是有植物的池子濁度卻只增加了15NTU,這說明了伊樂藻具有較好的穩定底質的作用,能降低沉積物質再懸浮的風險。試驗結束后采伊樂藻,鮮重為33.5g(含水率約90%),用蒸餾水沖洗莖、葉,將沖洗下來的懸浮態物質烘干,為0.54g,即每千克伊樂藻鮮重所吸附的懸浮態物質為16g,說明伊樂藻能吸附大量的懸浮物,從而使渾濁的水體較快清澈。伊樂藻水樣中的濁度降低效果明顯,濁度的最高凈降低率達到87.9%。
在115小時前,伊樂藻水樣中DO比對照水樣低(圖8),這主要與以下兩點有關:(1)測定時間,試驗值都是在早上的10:00測定的,此時,植物的光合作用不強烈;(2)原水狀態,原水表面有大量的油污,油污阻礙了光線和氧氣的進入,使水面下的水體處于缺氧狀態,從而使伊樂藻的光合作用受阻。當植物適應了這種生境后,水中的DO開始大幅度上升,遠高于對照樣,在216h和235h監測到的溶氧接近飽和值,并伴有小氣泡的逸出。因此只有當伊樂藻經過一段時間適應了沒有超過其生長閾值的逆境后,才會對水體產生明顯的富氧效果,在本試驗中,從115h開始,有伊樂藻的池子開始表現出明顯的富氧效果。

圖7 不同處理組濁度的變化
3 結 論
沉水植物對總氮、氨氮和總磷的去除率伊樂藻最好,平均去除率達到了20%以上,同時伊樂藻能吸附大量的懸浮物,水質的濁度降低效果明顯,最高凈降低率達到87.9%,并對水體產生明顯的富氧效果。
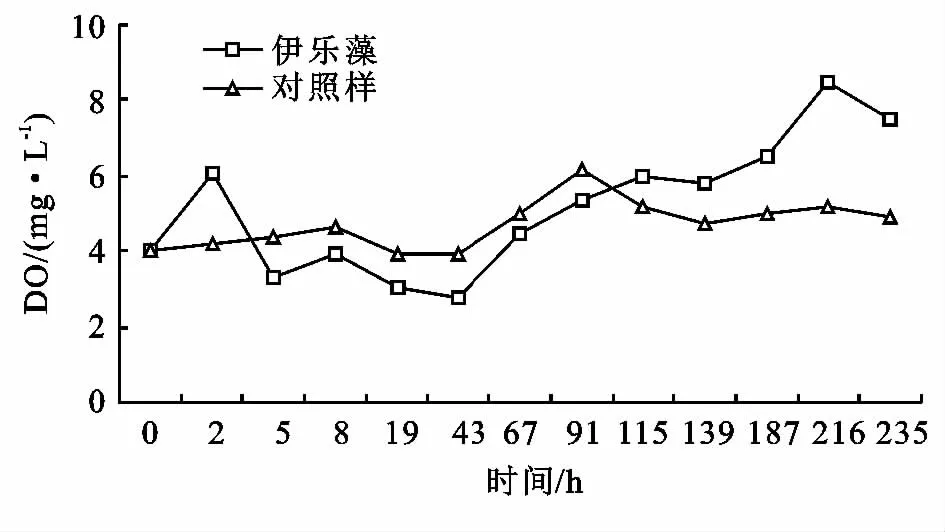
圖8 不同處理組DO的變化
冬季挺水植物中水芹相對生長率為48.5%~92.2%生長狀況最好,香蒲相對生長率為10.2%~31.5%生長狀況最差,冬季挺水植物對總氮、氨氮和總磷的去除率水芹最好;通過植株相對增長率顯著性分析,發現氮磷比是影響植株含磷量的顯著性因素,影響植物相對生長率的因素主次順序為:氮磷比>氮形態>氮濃度。冬季伊樂藻相對生長率為43%~94.2%,同等試驗條件下菹草生長緩慢,相對生長率為-23.9%~39.6%,伊樂藻的生長狀況最好,狐尾藻的生長狀況最差,對總氮、氨氮和總磷的去除率伊樂藻最好,平均去除率達到了20%以上。
綜上,建議水生植物以水芹和伊樂藻為全年物種并適當搭配其他植物作為前置庫凈化系統的植物組合。研究結果可為前置庫工程建設提供技術支撐。
[1] 李仰斌,張國華,謝崇寶.我國農村飲用水源現狀及相關保護對策建議[J].中國農村水利水電,2007(11):1-4.
[2] 柴世偉,斐曉梅,張亞雷,等.農業面源污染及其控制技術研究[J].水土保持學報,2006,20(6):192-195.
[3] Uhlmann D,Benndorf J.The use of primary reservoirs to control eutrophication caused by nutrient inflows from non-point sources[C]∥Land Use Impact on Lake and Reservoir Ecosystems Proceedings of a Regional Work Shop on MAB project 5.Warsaw Facultas Wien,1980:152-188.
[4] Nyholm N,Sorensen P E,Olrik K,et al.Restoration of lake nakskov indrefjord denmark,using algal ponds to remove nutrients from inflowing river water[J].Prog wat Technol.,1978,10(6):881-892.
[5] 常會慶,王世華,徐曉峰,等.兩種水生植物對重富營養化水體修復效果研究[J].水土保持研究,2012,19(5):161-165.
[6] 黃玉潔,張銀龍,李海東,等.太湖人工恢復濕地區植物群落建植對沉積物中氮、磷空間分布的影響[J].水土保持研究,2011,18(5):262-265.
[7] Budd R,O′geen A,Goh K S,et al.Removal mechanisms and fate of insecticides in constructed wetlands[J].Chemosphere 2011,83(11):1581-1587.
[8] 南楠,張波,李海東,等.洪澤湖濕地主要植物群落的水質凈化能力研究[J].水土保持研究,2011,18(1):228-231.
[9] Gumbricht T.Nutrient removal processes in freshwater submersed macrophyte system[J].Ecological Engineering,1993,2(1):1-30.
[10] Paul L.Nutrient elimination in pre-dams:results of long term studies[J].Hydrobiologia,2003,504(1/3):289-295.
[11] 段偉,劉昌明,黃炳彬.官廳水庫入庫口復合濕地系統對入庫水質的凈化[J].北京師范大學學報:自然科學版,2009,45(5/6):595-601.
[12] 田猛,張永春.用于控制太湖流域農村面源污染的透水壩技術試驗研究[J].環境科學學報,2006,26(10):1665-1670.
[13] Carr G M.Macrophyte growth and sediment phosphorus and nitrogen in a Canadian prairie river[J].Freshwater Biology,1998,39(3):525-536.
[14] Fernández-Aláez M,Fernández-Aláez C,Bécares E.Nutrient content in macrophytes in Spanish shallow lakes[J].Hydrobiologia,1999,408:317-326.
[15] 閻自申.前置庫在滇池流域運用研究[J].云南環境科學,1996,15(6):33-35.
[16] Mu?oz A R,Trevisan M,Capri E.Sorption and photo degradation of chlorpyrifos on riparian and aquatic macrophytes[J].J Environ Sci Health Part B,2008,44(1):7-12.
[17] Paul L,Putz K.Suspended matter elimination in a predam with discharge dependent storage level regulation[J].Limnologica,2008,38(3/4):388-399.
[18] Salvia-Castellvi M,Dohet A,Vander Borght P,et al.Control of the eutrophication of the reservoir of Eschsur-S?re(Luxembourg):evaluation of the phosphorus removal by predams[J].Hydrobiologia,2001,459(1/3):61-71.
[19] 國家環保局,《水和廢水監測分析方法》編委會.水和廢水監測分析方法[M].4版.北京:中國環境科學出版社,2002:16-30.
猜你喜歡
環境(2023年5期)2023-06-30 01:20:01
小讀者(2021年2期)2021-03-29 05:03:48
華人時刊(2019年13期)2019-11-17 14:59:54
當代水產(2019年1期)2019-05-16 02:42:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
