植物葉片水穩定同位素研究進展
2013-08-02 07:03:12余武生萬詩敏
生態學報 2013年4期
羅 倫,余武生 ,萬詩敏,周 平
(1.中國科學院青藏高原環境變化與地表過程重點實驗室,中國科學院青藏高原研究所,北京 100101;2.中國科學院研究生院,北京100049;3.天津城市建設學院,基礎學科部,天津 300384;4.長春氣象探測中心,長春 130062)
植物葉片不僅是聯系分子生態學和宏觀生態學的重要橋梁,同時也是植物光合作用和呼吸作用進行物質和能量交換的重要場所。因此,植物葉片水穩定同位素變化可以直接溝通植物葉片內部與外界的物質和能量聯系[1-2]。尤其是,通過研究植物葉片水穩定同位素可以反映植物生長周圍的氣象與生態信息,如溫度[3]、相對濕度[4]、氣壓[5]和蒸騰速率[6]以及氣孔導度等[7]。早在 1965 年,Gonfiantini等[8]就發現一些植物葉片水相對于植物水源會發生δ18O富集。隨后,植物葉片水δD的富集現象也被發現[9]。進一步研究證實,不同物種植物葉片水相對于植物水源(土壤水或莖水)均會發生穩定同位素分餾[10-11]。特別是,一些學者將研究開放水體蒸發過程時的Craig-Gordon模型進行修正并應用到植物葉片水上[12-14],使得植物葉片水穩定同位素研究得到進一步拓展。而在模型的發展與應用過程中,又促進了對植物葉片水穩定同位素分餾機制及其與生態因子的相互關系的了解[1,15]。
植物葉片水作為參與水循環的一個重要環節,了解葉片水穩定同位素與大氣水汽、降水、土壤水等不同水體中穩定同位素的相互關系,有助于揭示其在局地水體穩定同位素循環中的分配與貢獻。而在實際研究中,大氣水汽[16]、降水[17]、地表徑流[18]、湖水與土壤水[19]等不同圈層的多相態水體穩定同位素研究相對成熟,對于植物圈的葉片水穩定同位素與水循環相結合的研究卻相對較少,直到最近才逐漸受到科學家們的青睞。如,Lai等[20]曾在美國華盛頓州的南部森林建立了包含大氣水汽、降水、土壤水、植物葉片水、莖水以及在穩態和非穩態環境下經過蒸騰而進入空氣中的水汽等不同水體中穩定同位素水文循環。
在國內,穩定同位素生態學研究起步較晚[21-22],且大多數是關于確定植物水源[23-24]、水分利用[25]和生態效應[26]以及區分生態系統蒸散組分[27]等方面的研究,較少涉及到植物葉片水穩定同位素研究。近年來,該方面的研究逐漸得到重視:溫學發等[28]介紹了葉片水δ18O的富集和大氣水汽δ18O的原位觀測技術;Zhang等[29]用穩定同位素技術區分生態系統中的蒸散發;特別是張世春揭示了冬小麥葉片水穩定同位素的日變化過程,發現葉片水穩定氫、氧同位素均表現為在6:00最低,12:00—15:00區間最高的變化趨勢[30]。
隨著國內植物葉片水穩定同位素研究逐漸展開,相關成果將逐漸得以報道。根據目前國內外研究進展,本文詳細介紹了植物葉片水穩定氫、氧同位素在植物體中的分餾過程及形式,分析了葉片水穩定同位素組成與氣象和生態因子的關系,闡述了葉片水穩定同位素相關模型,并討論了這些模型可能存在的問題以及未來植物葉片水穩定同位素的研究趨勢。
1 植物葉片水穩定同位素分餾過程及形式
植物葉片水穩定同位素組成一般用δL或ΔL表示:
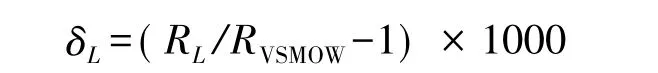
式中,RL為葉片水中穩定同位素比值(18O/16O或D/H);RVSMOW為“標準平均大洋水”穩定同位素比值。
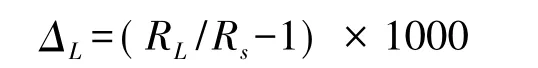
式中,ΔL表示葉片水相對于植物水源的穩定同位素組成;Rs為植物水源(莖水或土壤水)穩定同位素比值。
植物葉片水分來源于植物根系吸收土壤中的水,然后經過莖中導管傳輸到葉柄,最后進入植物葉片中。除泌鹽類植物外[31],一般情況下,水分在到達葉片之前均不會發生穩定同位素分餾。當水分到達葉片時,由于分餾效應,植物葉片水穩定同位素在葉片上的分布并不均勻。研究發現,從葉片基部(葉基)沿葉脈向葉尖δD值明顯升高,而且葉肉中的δD比葉脈、葉柄以及植物莖中的δD值均高[32]。δ18O同樣也存在類似的趨勢,由葉基向葉尖,葉脈向葉片邊緣δ18O值逐漸升高[33]。在自然條件下,植物葉片水穩定同位素會發生熱力學平衡分餾、動力學分餾和植物生化分餾3種形式的穩定同位素分餾。正是由于上述諸種分餾效應的存在導致葉片水穩定同位素在葉片各部位分布不均。
當水分到達葉片時,葉片水會發生蒸發,此時植物葉片水由液態變為氣態,在此相變過程中會發生熱力學平衡分餾效應。根據Majoube[34]的研究,與溫度相關的平衡分餾系數可表示為:
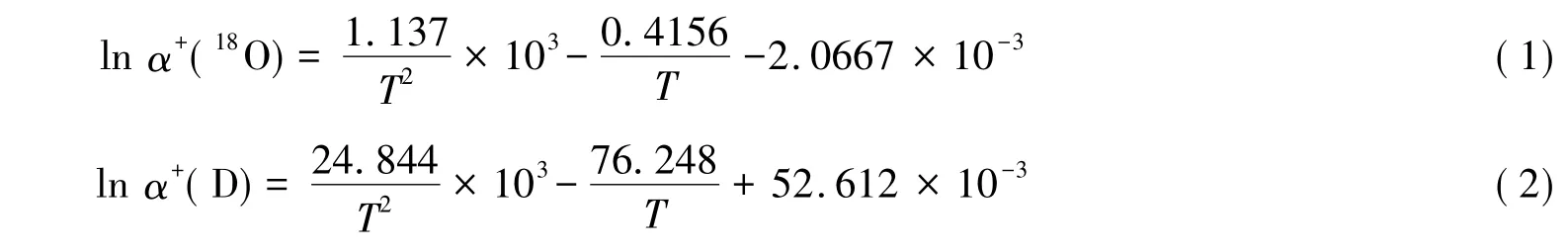
式中,α+表示平衡分餾系數,其與平衡分餾效應ε+的關系為:ε+=1000(α+-1)≈1000 lnα+(‰)(α+趨近于1時);T表示葉片的開爾文絕對溫度。
植物葉片水除了會發生熱力學平衡分餾外,水分子在穿過氣孔進入大氣過程中,需要克服一定的阻力,而輕、重同位素的擴散速率又不同,因此會發生動力學分餾。在此過程中,動力分餾效應(εk)與水分子擴散過程中受到的葉片氣孔阻力(rs)和邊界層阻力(rb)之間存在如下關系[35-37]:
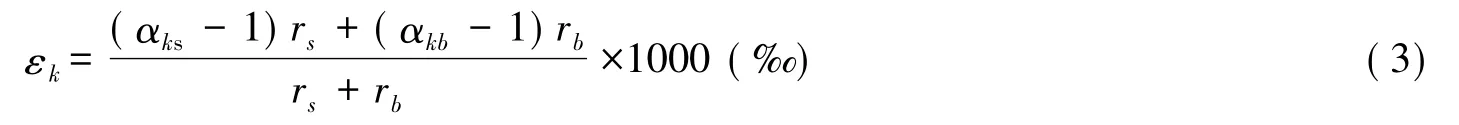
式中,αks表示水分子穿過氣孔時的動力分餾系數,αkb表示水分子穿過葉片邊界層時的動力分餾系數,它們之間的關系為 αkb=αks2/3[10]。Merlivat[36]計算出 αks是 1.025(H/D) 和 1.0285(16O/18O),此時相應的 αkb為1.017(H/D)和 1.0189(16O/18O)[13-14]。2003 年 Cappa 等[37]將 αks進一步修正為 1.016(H/D)和 1.032(16O/18O)。
另外,植物進行生理活動時,也會發生分餾。比如,植物進行光合作用時,空氣中的CO2作為重要反應物,通過氣孔進入葉片的CO2只有部分被固定:在與水合成葡萄糖的過程,羰基氧與H2O分子中氧原子發生交換,使得δ18O發生分餾[38];而未被吸收的部分CO2與葉片水在碳酸酐酶(CA)催化下,經過穩定同位素平衡反應后返回大氣,此時是以C18O16O的形式存在于大氣中[39-40],同樣發生δ18O分餾。而光合作用過程中攝取的氫參與合成有機物會發生一系列生物化學反應,這些生化反應會引起δD發生明顯的負同位素分餾效應[41]。另外,在一些脂類物質的合成中,也會發生生化分餾[3]。雖然植物葉片水存在生化分餾效應,但是與熱力學平衡分餾效應和動力學分餾效應相比,其分餾過程復雜。故本文未重點涉及生化分餾效應。
2 植物葉片水穩定同位素與氣象、生態因子的關系
2.1 氣象因子
在植物葉片水穩定同位素的平衡分餾過程中,其分餾程度受制于溫度的影響:隨著溫度升高,平衡分餾效應降低。目前關于葉片水δ18O和δD與溫度關系的研究結論不盡相同。Dongmann等[10]研究山毛櫸(Fagus silvatica)和橡木(Quercus robur)葉片水δ18O發現,其與溫度之間存在很好的正相關性。Zhou等[3]選取3種C3植物和3種C4植物分別在20℃和30℃時進行對比研究,結果顯示,在30℃時所有植物葉片水δD的值均比在20℃時的值大,即高溫有助于葉片水δD的富集。但是,Cooper和DeNiro[11]在加州大學洛杉磯分校校園(UCLA)和棕櫚沙漠鎮的峽谷研究中心(Deep Canyon)兩個地區研究5種C3植物和4種CAM植物葉片水δ18O和δD與溫度的關系時,發現僅在Deep Canyon干旱地區僅兩種C3植物葉片水的δ18O與溫度呈正相關性,Deep Canyon干旱地區的其它植物和UCLA地區所有植物的葉片水δ18O和δD與溫度之間均無統計意義。這說明,不同地區不同植物種類的葉片水穩定氫、氧同位素對溫度因子的響應并不一致。
相對濕度(h)是影響植物葉片水穩定同位素富集的一個重要因子。相對濕度對葉片水穩定同位素組成的影響是由大氣水汽壓(ea)和葉內細胞間水汽壓(ei)共同決定的,其關系為h=ea/ei[1]。當濕度增加,葉內外水汽壓差變小,植物蒸騰降低,葉片水富集作用減小[42],即相對濕度與葉片水穩定同位素之間存在負相關性。比如,干季比濕季植物葉片水更富集[4,43];白天比夜間植物葉片水更富集[44]。此外,研究發現相對濕度越高,相對濕度與葉片水穩定同位素之間的相關性更為顯著[45-46]。這可能與葉片自身空間異質性有關:相對濕度越大,異質性越小;相對濕度越小,異質性越大[46]。有研究證實,相對濕度越低,葉尖與葉基兩處的Δ18O差異越大[47-48]。同樣地,?antr?ucˇek等[32]發現ΔD也表現出類似的現象。
氣壓的變化導致植物蒸騰速率的改變,因而影響動力分餾過程,進而也影響葉片水的富集。不過,很少直接探討大氣氣壓對植物葉片水穩定同位素富集的影響,更多的是利用葉-氣水汽壓差(leaf-to-air vapor pressure difference,葉內水汽壓與大氣水汽壓的差值)或空氣水汽壓虧缺(atmospheric vapor pressure deficit,空氣飽和水汽壓與實際水汽壓的差值)研究其對葉片水穩定同位素富集的影響。如Ripullone等[7]對棉花葉片水研究發現,在氣溫為29℃時,葉片水Δ18O與葉-氣水汽壓差存在正相關;當溫度為20℃時,葉片水Δ18O與葉-氣水汽壓差的相關性增強。另外,Li等[5]通過對落葉松的研究證實了葉片水Δ18O與空氣水汽壓虧缺之間存在顯著的正相關。
風對蒸騰作用的影響取決于風速。微風能把氣孔外面的水蒸氣吹走,促使植物進一步蒸騰,但是強風可能引起氣孔關閉,植物蒸騰變慢[49]。因而,風速對植物葉片水穩定同位素富集的影響較為復雜,導致二者之間的相關性難以顯著[10]。不過,在森林水汽研究中,冠層上端的水汽δ18O與風速之間存在一定的聯系[20]。由于水汽δ18O與植物水源δ18O存在分餾[50],而水源δ18O又是葉片水δ18O的初始來源,故風速通過影響水汽δ18O而間接影響葉片水穩定同位素組成。
2.2 生態因子
植物在蒸騰過程中,水汽會逃逸出葉片,從而導致葉片水相對富集,相對而言,蒸騰出去的水汽δT相對貧化。蒸騰作用的大小受外界環境和內部阻力共同影響。植物葉片水穩定同位素組成與蒸騰速率(E)之間存在顯著的正相關。如,Sheshshayee等[6]在研究向日葵葉片水Δ18O時就發現這種相關性。另外,葉片各部位蒸騰速率的不均一性,導致葉片各部位富集程度不同。如,?antr?ucˇek等[32]發現雪花桉(Eucalyptus pauciflora)葉片無論在干旱或濕潤環境下,葉基的蒸騰速率小于葉尖的,因而葉基的ΔD值小于葉尖的。
氣孔導度gs通過影響植物葉片的蒸騰,進而對動力分餾過程產生一定的影響。如果其它因子的影響不變,氣孔導度gs變大時,則氣孔阻力rs減小,植物蒸騰速率增大,動力分餾效應相應變化。Sheshshayee等[6]指出轉基因豇豆葉片水Δ18O與gs之間呈正相關。而Ripullone等[7]發現,在溫度為20℃時,棉花葉片水Δ18O與gs二者之間存在顯著的負相關,而當溫度為29℃時,這種相關性稍微變弱。這種關于氣孔導度對葉片水穩定同位素的不同影響的報道,可能與植物種類的不同有關。
此外,葉片長度[51]、水分脅迫[52]、露水[53]等其它生態因子以及光合作用[39-41]等植物生理過程都會影響植物葉片水穩定同位素組成。
3 植物葉片水穩定氫、氧同位素關系及意義
早在1961年,Craig[54]發現全球大氣降水中δ18O和δD有很好的線性關系:δD=8δ18O+10。植物果汁中δ18O和δD也存在很好的線性關系,但是其斜率卻比當地大氣降水線的斜率低。比如,Bong等[55]研究西紅柿的果汁中δ18O和δD,發現其線性方程的斜率僅為2.52。Allison等[56]發現植物葉片上每一個位置的δ18O和δD之間也存在一定的線性關系,并將其稱為葉片水蒸騰線,定義為:
δD=mδ18O+b (4)式中,m的數學意義表示回歸線的斜率,b表示回歸線的截距。
葉片水蒸騰線的斜率m顯著低于當地降水線的斜率,但稍微高于蒸發點水汽δD與δ18O回歸線的斜率。比如Cernusak等[57]擬合了狹葉羽扇豆(Lupinus angustifolius)葉片水 δD與 δ18O之間的回歸方程為δD=3.1δ18O-6.4‰,而當地降水線方程為 δD=7.1δ18O+11.1‰;Kim 和 Lee[2]的研究表明棉花葉片水蒸騰線的斜率為3.0,蒸發點水汽δD與δ18O回歸線的斜率為2.6。目前研究結果中,葉片水蒸騰線的斜率m均較小。Cooper和DeNiro[11]研究發現9種植物葉片水蒸騰線的斜率從1.5到3.21不等,斜率的波動受當地氣溫和濕度的共同影響,而且斜率較小的植物葉片水穩定同位素與當地的氣溫和相對濕度的相關性更顯著。Yakir等[58]也發現葉片水蒸騰線的斜率m可能受相對濕度影響:在較濕潤(平均相對濕度59%)條件下,棉花葉片水蒸騰線的斜率為2.4;在較干燥條件(平均相對濕度45%)下,棉花葉片水蒸騰線的斜率為2.0。如果在干旱或半干旱條件下,斜率m會更低。如Gat等[59]發現在西亞干旱區正午時,多種植物葉片水蒸騰線的平均斜率在2.1到2.8之間,而在半干旱條件下,松針葉片水蒸騰線的斜率僅為1.3[56]。
在相同條件下,H182O相對于H162O發生的動力分餾效應大于DH16O相對于H162O的[17,60],故葉片水蒸騰線中斜率m越低,說明在分餾過程中動力分餾比平衡分餾的作用更大[60]。因此葉片水蒸騰線的斜率的大小可以指示葉片水穩定同位素動力分餾的強弱程度[2]。
4 葉片水穩定同位素主要模型
4.1 修正的Craig-Gordon穩態模型
由于直接測定植物葉片水穩定同位素相對復雜且費用較高,不能滿足大量實驗的數據要求。因此,通過一些比較容易測定的參數建立模型來確定植物葉片水穩定同位素值顯得很有必要。早期,生態學家將Craig-Gordon模型進行修正應用于植物蒸騰過程穩定同位素模擬。該模型基于假設植物蒸騰δT與植物莖水δs相等。但是通過比較模擬預測值與實際測定值發現,二者之間存在一定的差異。一般情況下,修正的Craig-Gordon穩態模型預測值比實際值高[13-14,32]。但是也有研究發現,修正的Craig-Gordon穩態模型預測葉片水穩定同位素的平均值比實際觀測值的平均值偏低[33,51]。其原因可能為:(1)該模型不能反映葉片水穩定同位素組成的葉片尺度空間的變化;(2)Craig-Gordon方程的穩態假設只對某一特定時段(比如,中午)或比較長的時間尺度(數天到數周)才有效[61]。
4.2 String-of-lakes模型
為了更好地模擬葉片水穩定同位素組成的葉片尺度的空間變化,string-of-lakes模型得以應運而生。該模型將葉片視為一個湖泊,葉基相當于湖泊的底部,葉尖相當于湖面。在湖水穩定同位素研究中,表層湖水由于蒸發導致穩定同位素富集,底層湖水蒸發小,則不容易富集。因而,從表層到底層,穩定同位素富集程度逐漸降低,形成一個梯度。同理,葉基到葉尖也會由于富集程度不同,產生一個梯度[62]。根據string-of-lakes模型,可把葉片從基部到葉尖等分成n段,每一部分的葉片水δ值是不同的,各段之間是呈梯度變化,然后分別求出各段的葉片水 δ值,第 n段葉片水 δn值可表示如下[51,62]:

式中,δa表示大氣水汽穩定同位素組成;δn表示液態水進入第n段的蒸發面(比如氣孔)時的δ值;F+和E分別代表流入和蒸發流出某一段的通量;h表示葉片周圍的相對濕度;ε=ε++(1-h)εk,ε+為液-氣平衡分餾效應,εk為動力分餾效應。在該模型中,先求解出各段的δ值,然后再對整個葉片求平均[51]。
4.3 Péclet效應的穩態模型——Farquhar& Lloyd模型
為了提高模型模擬的精確性,Farquhar和Lloyd[63]引入Péclet效應對Craig-Gordon模型進行修正。Péclet效應是指水在葉片中從葉脈到葉片邊緣蒸發點遷移過程中,由于輕水較重水容易蒸發,故在蒸發點重水富集,從而導致重同位素又會從葉片邊緣蒸發點逆向擴散到葉脈[52]。其原理先通過Craig-Gordon模型計算出蒸發點水的穩定同位素組成Δes(因其是不可測定的),可以近似表示為[35,63]:

式中,Δes、Δv分別表示葉片蒸發點水和大氣水汽相對于植物水源的穩定同位素組成。ea、ei分別表示大氣水汽壓和葉內細胞間水汽壓。在確定了 Δes后,可通過如下Péclet效應方程確定整個葉片水穩定同位素組成[63]:

式中,ΔLs為Péclet效應模型模擬的葉片水相對于植物水源的穩定同位素組成;Δes可以通過方程(6)得到;y為 Péclet數,其表達式為[63]:

式中,E表示蒸騰速率(mol·m-2·s-1);L表示葉脈到葉片邊緣蒸發點的有效長度(m);C表示水的摩爾濃度(5.56×104mol/m3);Dw表示是示蹤物(H182O或HDO)在水中的擴散率。
Péclet效應在Craig-Gordon模型上的應用,提高了模型模擬的精度[44,64]。但在野外條件下模擬效果并不理想[65],這是由于葉片各個部位的蒸發強度不同,葉片水運動的有效距離很難精確,而Péclet效應也僅考慮了橫向(葉脈到蒸發點)一個方向上的變化。于是,有學者提出通過增加Péclet數以進一步校正Craig-Gordon模型,即混合路徑 Péclet效應模型,公式如下[47]:

式中,Δx、Δrv和Δm分別表示經過Péclet效應校正后的葉片主脈、細葉脈以及葉肉等各組分水的Δ值;φx、φrv和φm表示對應的組分水在葉片水中的比重(φx+φrv+φm=1);ΔLs表示混合路徑模型的最后模擬值。一般情況下,由于細葉脈的水所占比例少,可以忽略中間項。而Cuntz等[61]認為,葉脈(主脈)一般在葉片的中央,而葉片的向上、下表面都有氣孔,都會導致葉片水的蒸發富集,且富集程度不同,故在穩態條件下需對葉片上、下表面分別進行Péclet效應校正。ΔLs可以表示為:
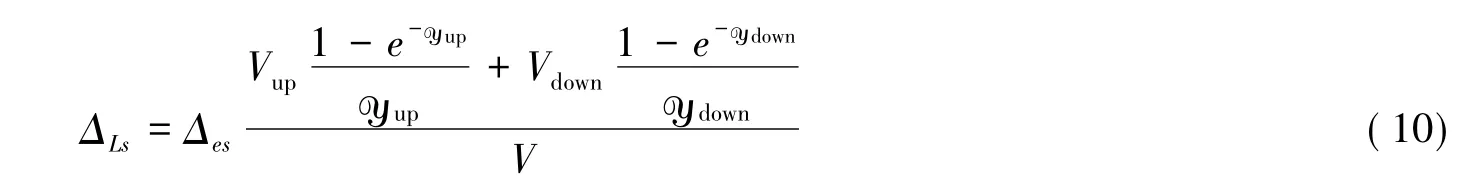
式中,V為整個葉片的含水量(mol/m2),Vup和Vdown分別表示葉上、下表面含水量(mol/m2),yup和ydown分別表示葉片上、下表面的Péclet數。
4.4 非穩態效應的模型——Dongmann模型
上述模型的建立均是在穩態條件下建立的,因此可以稱為穩態模型。然而研究發現植物葉片水穩定同位素只有在正午時分才能接近穩態。因此,要得到理想的模擬效果,模型建立的時間因素不能忽略。代表性的模型為1974年Dongmann等[10]提出的一種簡單的非穩態模型,該模型可以定量研究葉片水穩定同位素的分餾與富集。Cernusak等[57]人進一步將其完善如下:

式中,ΔLn表示非穩態模型計算出的葉片水相對于水源的穩定同位素組成;W表示為葉片水濃度(mol/m2);wi表示細胞間水蒸氣的摩爾分數(mol/mol);g表示氣孔導度和邊界層導度之和(mol·m-2·s-1);αk為動力分餾系數;t表示發生穩定同位素分餾的時間(s)。
4.5 Péclet效應的非穩態模型——Farquhar& Cernusak模型
但Dongmann模型沒有考慮Péclet效應。為了進一步提高模擬的精度,2005年Farquhar和Cernusak開發了一種同時考慮Péclet效應和非穩態效應的模型,其表示如下[65]:
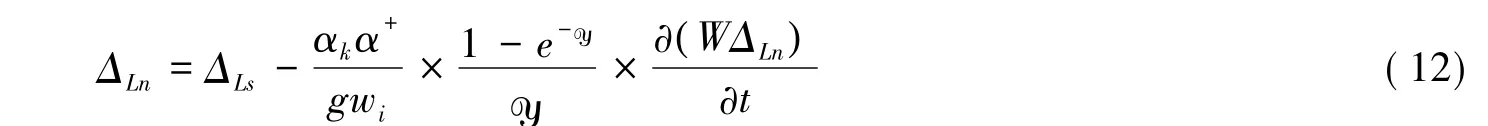
不過Cernusak等發現由于夜晚植物蒸騰速率小,y也就很小,因此在夜晚可以忽略Péclet效應[57]。故式(12)在夜間可以表示為方程(11)。
4.6 二維模型
近年來,一些學者開始嘗試從二維的角度構建相關模型。2007年Ogée等[64]從葉片解剖學的空間模式角度探討了穩定同位素在單子葉植物葉片的葉脈和葉肉中擴散的混合路徑,提出了二維模型的概念。該模型很好地解釋了C3和C4植物葉片水穩定同位素富集的差異是由于葉肉的曲折度引起,而不是簡單地取決于葉脈與蒸發點之間的直線距離[64]。之后Shu等[66]對二維模型進行了部分修正并應用于美國北部的松針的研究,該模型重要的部分在于引進了蒸騰速率的累積函數(f),精確地模擬了松針從葉基到葉尖δ18O的富集情況。不過,由于該模型可能未充分考慮植物所有生物過程,所以對葉片水δD的模擬效果仍不理想。
5 問題與展望
盡管植物葉片水穩定同位素研究取得了長足的進展,但該領域的研究還存在很多問題,主要表現在以下幾個方面:
在穩定同位素分餾與估算方面,目前葉片水穩定同位素平衡分餾的計算,主要是借鑒于其它水體穩定同位素平衡分餾機理。但是,植物葉片水穩定同位素分餾有其特殊性。比如,在植物葉片水中存在生化分餾,而其它水體幾乎不存在這種分餾形式,而且目前尚不清楚植物葉片水中的生化分餾會導致多大程度的重同位素富集或虧損。另外,不同種類的植物葉片水穩定同位素分餾也較其它水體復雜。如,在計算植物葉片水動力分餾時,只考慮氣孔阻力與邊界層阻力,這意味著植物葉片的蒸騰,全部通過氣孔進行。但是一些喜陰或喜濕的植物,角質層蒸騰是重要的部分,因此對于此種類的植物,需要考慮角質層阻力對穩定同位素分餾的影響。
在氣象與生態因子方面,相對濕度、蒸騰速率、葉片內外水汽壓差和水汽壓虧缺等因子均與葉片水穩定同位素之間存在顯著的正相關性。而溫度與部分植物的葉片水穩定同位素之間存在一定的正相關,但在某些植物葉片水中二者的相關性并不明顯[11]。另外,氣孔導度與葉片水Δ18O的關系也不確定,這可能與植物種類有關,即不同的植物,氣孔導度對葉片水Δ18O的影響可能不同。如Kodama等[67]研究發現單子葉植物黑小麥(Triticum×Secale)葉片水Δ18O主要受葉肉導度而不是氣孔導度的影響。Ferrio等[15]認為葡萄(Vitis vinifera)葉片水Δ18O受氣孔導度和水力導度共同作用。另外,一些環境因子(如風速、光照等)會改變蒸騰速率,但目前這些因子對葉片水穩定同位素的影響尚未發現統計學上的意義。
在穩定氫、氧同位素關系方面,很多報道已證實葉片水穩定氫、氧同位素之間存在線性相關,且葉片水蒸騰線的斜率指示葉片水穩定同位素的動力分餾[2]。而關于截距b的實際意義,鮮有報道。另外,在降水中過量氘d(d=δD-8δ18O)[17]是指示水汽來源的一項重要指標,而在植物葉片水穩定同位素研究中很少涉及過量氘及其意義的研究。
在模型方面,穩態模型中的修正的 Craig-Gordon模型[13-14]、string-of-lakes模型[62]、非穩態模型中的Dongmann模型[10]以及Cernusak等[57]人改進的非穩態模型均未考慮Péclet效應。穩態模型中的Farquhar&Lloyd 模型[63]、非穩態的Farquhar& Cernusak 模型[65]以及二維模型[66]均考慮了 Péclet效應。研究表明Péclet效應與 Craig-Gordon方程的耦合,雖然能夠提高穩態模型模擬的精度[44,64],但其模擬的效果仍然不及Farquhar&Cernusak模型的理想[1]。string-of-lakes模型考慮了葉片水δ值從葉基到葉尖縱向的不同,故其對平行脈葉片的水穩定同位素模擬具有一定的優勢,而對網狀脈葉片的模擬效果較差[32-33,51]。二維模型能夠提高δ18O的模擬精度[66],但該模型對δD的應用有待進一步完善。
因此,結合國內外的研究現狀和存在的問題,需要從以下幾個方面進行改進和拓展:
(1)加強植物葉片水穩定同位素在生理過程中變化的研究,明確不同生理過程中生化分餾效應對重同位素富集或虧損的影響。
(2)由于葉片水穩定同位素組成與溫度、風速、氣孔導度等氣象、生態因子之間的關系仍然不夠明確。因此,需要深入研究葉片水穩定同位素組成與這些要素之間的聯系,進一步揭示葉片水穩定同位素富集的內在機制。
(3)加強葉片水中的δ18O和δD之間的線性關系研究,在已有的研究基礎上,充分挖掘擬合曲線斜率m和截距b所代表的實際意義,并進一步揭示植物葉片水蒸騰線的斜率m遠低于大氣降水線的斜率的原因。另外,需要加強葉片水過量氘的實際意義的研究。
(4)當前模型的開發大多是在室內通過控制某些氣象和生態因子模擬而成,因此需要通過野外實際測量,判定相關模型對不同氣候區不同類型的植物的適用性,特別是在極端環境下(如,青藏高原等)的應用情況,并進一步完善相關模型。
(5)加強區域穩定同位素循環研究,揭示植物葉片水與當地大氣降水、地表徑流、土壤水、地下水以及大氣水汽等不同水體穩定同位素的內在聯系,從而構建區域穩定同位素的循環模式。
最后值得一提的是,隨著技術的發展,近年來出現了穩定同位素激光分析儀(IRIS),這種新的同位素分析儀具有成本低、體積小、操作簡單以及可實現原位測量等優點。但是穩定同位素激光分析儀在測量植物水或土壤水時,由于水樣中有機污染物的存在會引起較大的誤差[68]。為了減小這種誤差,廠商發布了一種數據軟件,而實際分析發現,該數據軟件只能進行數據質量控制,無法消除有機物的干擾[69-70]。于是,最近又推出了一種微型高溫熱解模塊技術,期望通過對有機物燃燒以消除有機物的存在對測量結果的影響。新技術的不斷突破,對植物葉片水穩定同位素研究帶來了測量的便捷和精度的提高,將推動植物葉片水穩定同位素研究的進一步發展。
[1]Kahmen A,Simonin K,Tu K,Goldsmith G R,Dawson T E.The influence of species and growing conditions on the 18-O enrichment of leaf water and its impact on‘effective path length’.New Phytologist,2009,184(3):619-630.
[2]Kim K,Lee X.Isotopic enrichment of liquid water during evaporation from water surfaces.Journal of Hydrology,2011,399(3/4):364-375.
[3]Zhou Y,Grice K,Chikaraishi Y,Stuart-Williams H,Farquhar G D,Ohkouchi N.Temperature effect on leaf water deuterium enrichment and isotopic fractionation during leaf lipid biosynthesis:Results from controlled growth of C3and C4land plants.Phytochemistry,2011,72(2/3):207-213.
[4]Cernusak L A,Mejia-Chang M,Winter K,Griffiths H.Oxygen isotope composition of CAM and C3Clusia species:non-steady-state dynamics control leaf water18O enrichment in succulent leaves.Plant,Cell and Environment,2008,31(11):1644-1662.
[5]Li S-G,Tsujimura M,Sugimoto A,Sasaki L,Yamanaka T,Davaa G,Oyunbaatar D,Sugita M.Seasonal variation in oxygen isotope composition of waters for a montane larch forest in Mongolia.Trees-Structure and Function,2006,20(1):122-130.
[6]Sheshshayee M S,Bindumadhava H,Ramesh R,Prasad T G,Lakshminarayana M R,Udayakumar M.Oxygen isotope enrichment(Δ18O)as a measure of time-averaged transpiration rate.Journal of Experimental Botany,2005,56(422):3033-3039.
[7]Ripullone F,Matsuo N,Stuart-Williams H,Wong S C,Borghetti M,Tani M,Farquhar G.Environmental effects on oxygen isotope enrichment of leaf water in cotton leaves.Plant Physiology,2008,146(2):729-736.
[8]Gonfiantini R,Gratziu S,Tongiorgi E.Oxygen isotopic composition of water in leaves//Isotopes and Radiation in Soil-Plant Nutrition Studies.Vienna:Proceedings of a Symposium,International Atomic Energy Agency,1965:405-410.
[9]Wershaw R L,Friedman I,Heller S J,Frank P A.Hydrogen isotopic fractionation of water passing through trees//Hobson G D,Speers G C,eds.Advances in Organic Geochemistry.Oxford:Pergamon Press,1970:55-67.
[10]Dongmann G,Nürnberg H W,F?rstel H,Wagener K.On the enrichment of H182O in the leaves of transpiring plants.Radiation and Environmental Biophysics,1974,11(1):41-52.
[11]Cooper L W,DeNiro M J.Covariance of oxygen and hydrogen isotopic compositions in plant water:species effects.Ecology,1989,70(6):1619-1628.
[12]Craig H,Gordon L.Deuterium and oxygen-18 variations in the ocean and the marine atmosphere//Tongiorgi E,ed.Stable Isotopes in Oceanographic Studies and Paleotemperatures.Spoleto,1965:9-130.
[13]Flanagan L B,Ehleringer J R.Stable isotope composition of stem and leaf water:Applications to the study of plant water use.Functional Ecology,1991,5(2):270-277.
[14]Flanagan L B,Comstock J P,Ehleringer J R.Comparison of modeled and observed environmental influences on the stable oxygen and hydrogen isotope composition of leaf water in Phaseolus vulgaris L.Plant Physiology,1991,96(2):588-596.
[15]Ferrio J P,Pou A,Florez-Sarasa I,Cessler A,Kodama N,Flexas J,Ribas-Carbó M.The Péclet effect on leaf water enrichment correlates with leafhydraulic conductance and mesophyll conductance for CO2.Plant,Cell and Environment,2012,35(3):611-625.
[16]Wen X F,Zhang S C,Sun X M,Yu G R,Lee X.Water vapor and precipitation isotope ratios in Beijing,China.Journal of Geophysical Research,2010,115:D01103,doi:10.1029/2009JD012408.
[17]Dansgaard W.Stable isotopes in precipitation.Tellus,1964,16(4):436-468.
[18]Meredith K T,Hollins S E,Hughes C E,Cendón D I,Hankin S,Stone D J M.Temporal variation in stable isotopes(18O and2H)and major ion concentrations within the Daring River between Bourke and Wilcannia due to variable flows,saline groundwater influx and evaporation.Journal of Hydrology,2009,378(3/4):313-324.
[19]Gibson J J,Birks S J,Edwards T W D.Global prediction of δAand δ2H-δ18O evaporation slopes for lakes and soil water accounting for seasonality.Global Biogeochemical Cycles,2008,22:GB2031,doi:10.1029/2007GB002997.
[20]Lai C T,Ehleringer J R,Bond B J,Paw U K T.Contributions of evaporation,isotopic non-steady state transpiration and atmospheric mixing on the δ18O of water vapour in Pacific Northwest coniferous forests.Plant,Cell and Environment,2006,29(1):77-94.
[21]Wu S H,Pan T,Dai E F.The progress and prospect of stable isotopes in plants.Progress in Geography,2006,25(3):1-11.
[22]Lin G H.Stable isotope ecology:a new branch of ecology resulted from technology advances.Chinese Journal of Plant Ecology,2010,34(2):119-122.
[23]Duan D Y,Ouyang H,Song M H,Hu Q W.Water sources of dominant species in three alpine ecosystems on the Tibetan Plateau,China.Journal of Integrative Plant Biology,2008,50(3):257-264.
[24]Zhou C X,Sun Z Y,Yu S W.Using D and18O stable isotopes to determine the water sources of sand dune plants in Linze,middle reaches of Heihe River.Geological Science and Technology Information,2011,30(5):103-109.
[25]Sun S J,Meng P,Zhang J S,Huang H,Wan X C.Deuterium isotope variation and water use in an agroforestry system in the rocky mountainous area of North China.Acta Ecologica Sinica,2010,30(14):3717-3726.
[26]Sun S,Huang J,Han X,Lin G.Comparisons in water relations of plants between newly formed riparian and non-riparian habitats along the bank of Three Gorges Reservoir,China.Trees-Structure and Function,2008,22(5):717-728.
[27]Yuan G F,Zhang N,Sun X M,Wen X F,Zhang S C.Partitioning wheat field evapotranspiration using Keeling Plot method and continuous atmospheric vapor δ18O data.Chinese Journal of Plant Ecology,2010,34(2):170-178.
[28]Wen X F,Zhang S C,Sun X M,Yu G R.Recent advances in H182O enrichment in leaf water.Chinese Journal of Plant Ecology,2008,32(4):961-966.
[29]Zhang S C,Wen X F,Wang J L,Yu G R,Sun X M.The use of stable isotopes to partition evapotranspiration fluxes into evaporation and transpiration.Acta Ecologica Sinica,2010,30(4):201-209.
[30]Zhang S C.Continuous Measurements of Atmospheric Water Vapor δ18O and δD for Hydrological and Ecological Applications [D].Beijing:Graduate University of Chinese Academy of Sciences(Institute of Geographic Sciences and Natural Resources Research),2010.
[31]Lin G H,Sternberg S L.Hydrogen isotopic fractionation by plant roots during water uptake in coastal wetland plants//Ehleringer J R,Hall A E,Farquhar G D,eds.Stable Isotopes and Plant Carbon-Water Relations.San Diego:Academic Press,1993:497-510.
[32]?antr?ucˇek J,Kveˇtonˇek J,?etlík J,Bulícˇková L.Spatial variation of deuterium enrichment in bulk water of snowgum leaves.Plant Physiology,2007,143(1):88-97.
[33]Gan K S,Wong S C,Yong J W H,Farquhar G D.18O spatial patterns of vein xylem water,leaf water,and dry matter in cotton leaves.Plant Physiology,2002,130(2):1008-1021.
[34]Majoube M.Fractionnement en oxygène 18 et en deutérium entre l'eau et sa vapeur.Journal de Chimie Physique et de Physico-Chimie Biologique,1971,10:1423-1436.
[35]Farquhar G D,Hubick K T,Condon A G,Richards R A.Carbon isotope fractionation and plant water-use efficiency//Rundel P W,Ehleringer J R,Nagy K A,eds.Stable Isotopes in Ecological Research.New York Springer,1988,68:21-40.
[36]Merlivat L.Molecular diffusivities of H216O,HD16O and H218O in gases.Journal of Physical Chemistry,1978,69(6):2864-2871.
[37]Cappa C D,Hendricks M B,DePaolo D J,Cohen R C.Isotopic fractionation of water during evaporation.Journal of Geophysical Research,2003,108(D16):4525-4534.
[38]Guy R D,Fogel M L,Berry J A.Photosynthetic fractionation of the stable isotopes of oxygen and carbon.Plant Physiology,1993,101(1):37-47.
[39]Gillon J S,Yakir D.Naturally low carbonic anhydrase activity in C4and C3plants limits discrimination against C18OO during photosynthesis.Plant,Cell and Environment,2000,23(9):903-915.
[40]Gillon J S,Yakir D.Influence of carbonic anhydrase activity in terrestrial vegetation on the18O content of atmospheric CO2.Science,2001,291(5513):2584-2587.
[41]Yakir D,DeNiro M J.Oxygen and hydrogen isotope fractionation during cellulose metabolism in Lemna gibba L.Plant Physiology,1990,93(1):325-332.
[42]Butt S,Ali M,Fazil M,Latif Z.Seasonal variations in the isotopic composition of leaf and stem water from an arid region of Southeast Asia.Hydrological Sciences Journal,2010,55(5):844-848.
[43]Lai C T,Ometto J P H B,Berry J A,Martinelli L A,Domingues T F,Ehleringer J R.Life form-specific variations in leaf water oxygen-18 enrichment in Amazonian vegetation.Oecologia,2008,157(2):197-210.
[44]Snyder K A,Monnar R,Poulson S R,Hartsough P,Biondi F.Diurnal variations of needle water isotopic ratios in two pine species.Trees-Structure and Function,2010,24(3):585-595.
[45]Kahmen A,Simonin K,Tu K P,Merchant A,Callister A,Siegwolf R,Dawson T E,Arndt S K.Effects of environmental parameters,leaf physiological properties and leaf water relations on leaf water δ18O enrichment in different Eucalyptus species.Plant,Cell and Environment,2008,31(6):738-751.
[46]Helliker B R.On the controls of leaf-water oxygen isotope ratios in the atmospheric crassulacean acid metabolism epiphyte Tillandsia usneoides.Plant Physiology,2011,155(4):2096-2107.
[47]Farquhar G D,Gan K S.On the progressive enrichment of the oxygen isotopic composition of water along a leaf.Plant,Cell and Environment,2003,26(6):801-819.
[48]Helliker B R,Ehleringer J R.Differential18O enrichment of leaf cellulose in C3versus C4grasses.Functional Plant Biology,2002,29(4):435-442.
[49]Li Q,Tan X L.Research advances in arid plant evapotranspiration.Gansu Agricultural Science and Technology,2006,12(10):18-20.
[50]Song X,Barbour M M,Saurer M,Helliker B R.Examining the large-scale convergence of photosynthesis-weighted tree leaf temperatures through stable oxygen isotope analysis of multiple data sets.New Phytologist,2011,192(4):912-924.
[51]Helliker B R,Ehleringer J R.Establishing a grassland signature in veins:18O in the leaf water of C3and C4grasses.Proceedings of the National Academy of Sciences of the United States of America,2000,97(14):7894-7898.
[52]Welp L R,Lee X,Kim K,Griffis T J,Billmark K A,Baker J M. δ18O of water vapour,evapotranspiration and the sites of leaf water evaporation in a soybean canopy.Plant,Cell and Environment,2008,31(9):1214-1228.
[53]Kim K,Lee X.Transition of stable isotope ratios of leaf water under simulated dew formation.Plant,Cell and Environment,2011,34(10):1790-1801.
[54]Craig H.Isotopic variations in meteoric water.Science,1961,133(3465):1702-1703.
[55]Bong Y S,Lee K S,Shin W J,Ryu J S.Comparison of the oxygen and hydrogen isotopes in the juices of fast-growing vegetables and slow-growing fruits.Rapid Communications in Mass Spectrometry,2008,22(18):2809-2812.
[56]Allison G B,Gat J R,Leaney F W.The relationship between deuterium and oxygen-18 delta values in leaf water.Chemical Geology:Isotope Geoscience Section,1985,58(1/2):145-156.
[57]Cernusak L A,Pate J S,Farquhar G D.Diurnal variation in the stable isotope composition of water and dry matter in fruiting Lupinus angustifolius under field conditions.Plant,Cell and Environment,2002,25(7):893-907.
[58]Yakir D,DeNiro M J,Gat J R.Natural deuterium and oxygen-18 enrichment in leaf water of cotton plants grown under wet and dry conditions:evidence for water compartmentation and its dynamics.Plant,Cell and Environment,1990,13(1):49-56.
[59]Gat J R,Yakir D,Goodfriend G,Fritz P,Trimborn P,Lipp J,Gev I,Adar E,Waisel Y.Stable isotope composition of water in desert plants.Plant and Soil,2007,298(1/2):31-45.
[60]Cooper L W,DeNiro M J,Keeley J E.The relationship between stable oxygen and hydrogen isotope ratios of water in astomatal plants//Taylor H P,O'Neil J R,Kaplan I R,eds.Stable Isotope Geochemistry:A Tribute to Sam Epstein.San Antonio:The Geochemical Society,Special Publication,1991:247-255.
[61]Cuntz M,Ogée J,Farquhar G D,Peylin P,Cernusak L A.Modelling advection and diffusion of water isotopologues in leaves.Plant,Cell and Environment,2007,30(8):892-909.
[62]Gat J R,Bowser C.The heavy isotope enrichment of water in coupled evaporative systems//Taylor H P,O'Neil J R,Kaplan I R,eds.Stable Isotope Geochemistry:A Tribute to Samuel Epstein.San Antonio:The Geochemical Society,Special Publication,1991:159-168.
[63]Farquhar G D,Lloyd J.Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere//Ehleringer J R,Hall A E,Farquhar G D,eds.Stable Isotopes and Plant Carbon-Water Relations.San Diego:Academic Press,1993:47-70.
[64]Ogée J,Cuntz M,Peylin P,Bariac T.Non-steady-state,non-uniform transpiration rate and leaf anatomy effects on the progressive stable isotope enrichment of leaf water along monocot leaves.Plant,Cell and Environment,2007,30(4):367-387.
[65]Farquhar G D,Cernusak L A.On the isotopic composition of leaf water in the non-steady state.Functional Plant Biology,2005,32(4):293-303.
[66]Shu Y,Feng X,Posmentier E S,Sonder L J,Faiia A M,Yakir D.Isotopic studies of leaf water.Part 1:A physically based two-dimensional model for pine needles.Geochimica et Cosmochimica Acta,2008,72(21):5175-5188.
[67]Kodama N,Cousins A,Tu K P,Barbour M M.Spatial variation in photosynthetic CO2,carbon and oxygen isotope discrimination along leaves of the monocot triticale(Triticum × Secale)relates to mesophyll conductance and the Péclet effect.Plant,Cell and Environment,2011,34(9):1548-1562.
[68]West A G,Goldsmith G R,Brooks P D,Dawson T E.Discrepancies between isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters.Rapid Communications in Mass Spectrometry,2010,24(14):1948-1954.
[69]West A G,Goldsmith G R,Matimati I,Dawson T E.Spectral analysis software improves confidence in plant and soil water stable isotope analyses performed by isotope ratio infrared spectroscopy(IRIS).Rapid Communications in Mass Spectrometry,2011,25(16):2268-2274.
[70]Zhao L,Xiao H,Zhou J,Wang L,Cheng G,Zhou M,Yin L,McCabe M F.Detailed assessment of isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters.Rapid Communications in Mass Spectrometry,2011,25(20):3071-3082.
參考文獻:
[21]吳紹洪,潘韜,戴爾阜.植物穩定同位素研究進展與展望.地理科學進展,2006,25(3):1-11.
[22]林光輝.穩定同位素生態學:先進技術推動的生態學新分支.植物生態學報,2010,34(2):119-122.
[24]周辰昕,孫自永,余紹文.黑河中游臨澤地區沙丘植物水分來源的D、18O同位素示蹤.地質科技情報,2011,30(5):103-109.
[25]孫守家,孟平,張勁松,黃輝,萬賢崇.華北石質山區核桃-綠豆復合系統氘同位素變化及其水分利用.生態學報,2010,30(14):3717-3726.
[27]袁國富,張娜,孫曉敏,溫學發,張世春.利用原位連續測定水汽δ18O值和Keeling Plot方法區分麥田蒸散組分.植物生態學報,2010,34(2):170-178.
[28]溫學發,張世春,孫曉敏,于貴瑞.葉片水H182O富集的研究進展.植物生態學報,2008,32(4):961-964.
[30]張世春.大氣水汽δ18O和δD原位連續觀測及其應用研究[D].北京:中國科學院研究生院(地理科學與資源研究所),2010.
[49]李倩,譚雪蓮.旱地植物蒸騰作用研究進展.甘肅農業科技,2006,12(10):18-20.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
中學物理·高中(2016年12期)2017-04-22 11:53:03
爆笑show(2016年7期)2017-02-09 09:36:13
光學精密工程(2016年6期)2016-11-07 09:07:19
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
