吐倫球堅蚧生物學特性研究
2013-06-11 10:29:14任金龍張魯豫賈曉江
環境昆蟲學報 2013年6期
范 毅,趙 莉*,任金龍,張魯豫,賈曉江
(1.新疆農業大學農學院,烏魯木齊 830052;2.新疆輪臺縣森林病蟲害防治檢疫站,新疆輪臺 841600)
吐倫球堅蚧Rhodococcus turanicus Arch.又名中亞朝球蠟蚧,屬于半翅目 Hemiptera,蚧科Coccidae,褐球蚧屬Rhodococcus。主要危害杏、李、桃、蘋果、梨等,是林果業重要害蟲,分布于俄羅斯,新疆(湯祊德,1991;王子清,2001;梁愛萍,2005)。新疆是杏的起源中心之一(羅新書 等,1998)。輪臺白杏Armeniaca vulgaris cv.Luntaibaixing 被譽為“杏中極品”是原產于新疆輪臺縣境內的一種古老的地方優良杏品種,栽培歷史達1000 多年,并已獲準國家原產地域產品保護。目前輪臺縣杏樹面積超過16000 hm2,其中輪臺白杏占60% 以上,并且已逐步進入盛果期(程衛東等,2006)。近年來由于吐倫球堅蚧的蔓延,使得輪臺縣約80%的杏園都被其危害,尤其是老杏園被害率達100%(張滋林等,2010)。該蟲成蟲與若蟲不僅以口針刺吸汁液危害,造成樹勢衰弱甚至整株死亡,而且大量排出蜜露誘發煤污病從而阻礙寄主植物的光合作用,嚴重影響了杏的產量與品質。Babayan(1970)對吐倫球堅蚧的形態特征進行了描述;朱曉鋒等(2009)對吐倫球堅蚧若蟲在田間的空間分布進行了初步研究;楊森等(2008)探索了草皮逍遙蛛對其的捕食作用;范毅等(2013)對其的藥劑防治方法及防治時期進行了系統的研究。2009年至2012年,在前人的研究基礎上,通過田間調查和室內觀察相結合的方法,對吐倫球堅蚧生物學特性進行了系統的研究,以期為該害蟲的綜合防控提供理論依據。
1 材料與方法
1.1 樣地概況
樣地選擇在輪臺縣周邊,位于東經84°14'37.13″~84°16'1.58″,北緯41°44'40.14″~41°47'52.66″。所選樣地內杏樹品種均為小白杏Armeniaca vulgaris cv.Luntaibaixing,樹齡為8~12年,樹高4~6 m,間作冬小麥。
1.2 室外調查
2009~2012年,選擇4 塊樣地,每樣地隨機確定5株杏樹,被選樹按東、南、西、北各固定一枝2~4年生20 cm 枝條或5片葉片,每7 d 調查1次(除12月到翌年2月),觀察吐倫球堅蚧的生活史、發生規律及杏樹的物候期。10月~11月,調查1~6年生(鑒別以芽齡痕為準)10 cm枝條上的若蟲數量,來年3月查看各枝條上若蟲的越冬死亡率。
當吐倫球堅蚧雌雄形態初顯差異時,每塊樣地隨機選擇20 枝枝條,每枝枝條保留15~25頭未交尾的雌蟲并用15 cm×35 cm 的紗網籠罩,待雌成蟲產卵末期將籠罩的枝條采回實驗室,觀察其產卵量及卵孵化率。
雌蟲體型漸膨大,初排蜜露時,選擇同一齡級桃,梨,杏混交林為試驗地,桃、梨、杏各取3株,每株樹固定2 枝2~4年生(保留15頭雌成蟲)枝條,將10 cm×30 cm 濾紙編號稱重后固定在選中枝條下3~5 cm 處,用40 cm×100 cm 紗網籠罩,每日12 時收集,直至雌蟲不再排出蜜露。
1.3 室內測定
1.3.1 成蟲習性的研究
3月上中旬,從田間采回大批(100~150 枝)帶越冬若蟲的枝條置于室內(溫度22℃~25℃,濕度25%~35%),隨機取60 枝用12 cm×20 cm的紗網分別籠罩,每日記錄雄蟲羽化數與雌蟲膨大數,統計雌雄性比。
將20頭雄蛹放入指形管(直徑1 cm),用脫脂棉塞住管口,每2 h 觀察雄蟲的羽化規律。再將初羽化的雄蟲移入內有3~5頭未交尾雌蟲的培養皿中,觀察其交尾行為。每處理重復3次。
當雌成蟲開始孕卵時,每3 天從田間采回一批(300~400頭)雌成蟲,并隨機對其中100頭進行剖查、統計產卵量,直至雌蟲不再產卵,從調查當日起逐日記錄當地的氣溫。
雌成蟲產卵末期,用游標卡尺和電子天平(1/10000)隨機對60頭雌成蟲的體長、體重及其產卵量進行測量與統計。
1.3.2 卵發育與溫度的關系
將同日產出的卵分別放置在5個不同溫度梯度(16℃、20℃、24℃、28℃、32℃)的恒溫條件下(人工氣候箱PQX-280B-22H)和5 處溫度差異大的變溫條件下(分別用溫濕度記錄儀EL-USB-2 記錄變溫條件下的溫、濕度值),逐日分0∶00、8∶00、16∶00 檢查卵孵化量,各處理重復5次。
1.3.3 若蟲習性研究
若蟲初孵時,每日10∶00、18∶00 檢查涌出介殼(雌蟲體)的若蟲數量,直至3 d 未有若蟲爬出,掀開介殼清點未孵化卵量,求出孵化率,試驗重復15次。
在一間僅北面有自然光,東、南、西以窗簾遮光的室內,將100頭雌成蟲(每頭雌成蟲體內約含1500頭初孵若蟲與卵)置于地面中心點,72 h后測量各方位若蟲的爬行距離,試驗重復3次。
2011年6月~9月,從每塊樣地隨機取100片樹葉帶回室內,觀察并統計每片樹葉正面和背面吐倫球堅蚧若蟲總數,并同時記錄在葉片上可觀察到的、若蟲自然死亡數、被捕食數及寄生數。
1.4 數據分析(牟吉元,1997)
吐倫球堅蚧卵的發育起點溫度(C)和有效積溫(K)的計算公式分別為:
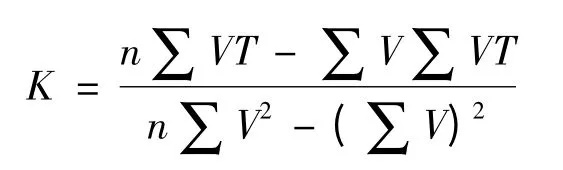
發育起點溫度(C)和有效積溫(K)的標準誤分別為Sc、Sk。
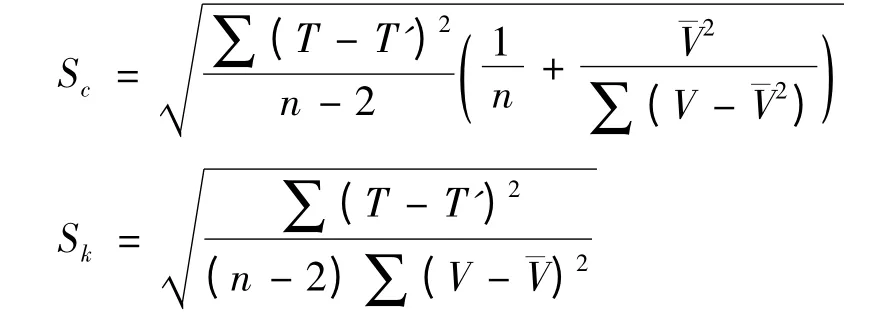
試驗數據均采用Excel 2003、SPSS 18.0 軟件進行分析。
2 結果與分析
2.1 生活史
通過2009年~2012年連續4年在杏園的系統調查,吐倫球堅蚧在新疆輪臺縣1年發生1代(表1),以2齡若蟲在枝條上越冬。3月中、下旬,當旬平均溫度為7.1±1.4℃時,越冬2齡若蟲開始化蛹,此時正逢杏樹現蕾。從4月開始,當旬平均溫度達到16.2±2.6℃時,雄蟲開始羽化,此時杏花凋謝,杏樹坐果。當旬平均溫度達到21±0.8℃時,雌蟲開始產卵,杏樹果實逐漸膨大。從5月開始,旬平均溫度為20.9±0.3℃,杏樹果實開始著色并逐漸成熟,此時卵開始孵化。6月~8月間,月平均溫度均在25℃以上,在此期間若蟲固著于葉片取食,直到10月,當旬平均溫度為8.8±1.1℃,杏樹進入落葉期時,若蟲由葉片向枝條轉移開始越冬。

表1 吐倫球堅蚧年生活史Table1 Life history of Rhodococcus turanicus
2.2 發生規律
吐倫球堅蚧在田間的發生規律見圖1。越冬若蟲自3月開始發育,體背分泌白色蠟絲并逐漸將身體包裹形成蠟殼。隨進一步的發育,3月中下旬,越冬若蟲蠟殼內蛻皮后進入蛹期,此時蠟殼與蟲體分離,雌雄逐漸分化,雌若蟲由橢圓形發育為扁圓狀,蠟殼與蟲體緊貼不易分離。4月中旬,雄蟲大量羽化,雌成蟲體型迅速膨大,并由臀裂基部排出大量蜜露。雌雄交尾后1~2 d,雄成蟲大量死亡,雌成蟲體型膨大為近球形,體背逐漸硬化,并分泌白色蠟粉。在4月下旬,雌成蟲開始產卵。5月中旬雌成蟲停止產卵,同時雌成蟲大量死亡。5月下旬若蟲孵化,初孵若蟲體小,橙紅色,藏于介殼內不立即爬出。由于雌成蟲死亡,介殼失水,臀裂處開裂,大量初孵若蟲涌出介殼,6月上旬為涌散高峰(圖2),初孵若蟲經1~3 d 爬行至葉片、嫩枝、果實等處開始固定取食,固定后4~7 d 體色由橙紅漸變為淺黃色。8月下旬一齡若蟲蛻皮進入2齡,若蟲蛻皮后體型略微增長,體表分泌一層透明蠟質薄膜。10月下旬二齡若蟲由葉片向枝條轉移,11月中下旬完成轉移,開始越冬(圖3)。
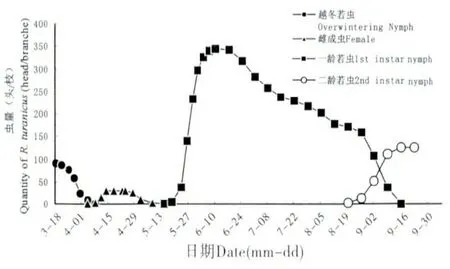
圖1 吐倫球堅蚧田間消長規律Fig.1 Population dynamics of Rhodococcus turanicus in field
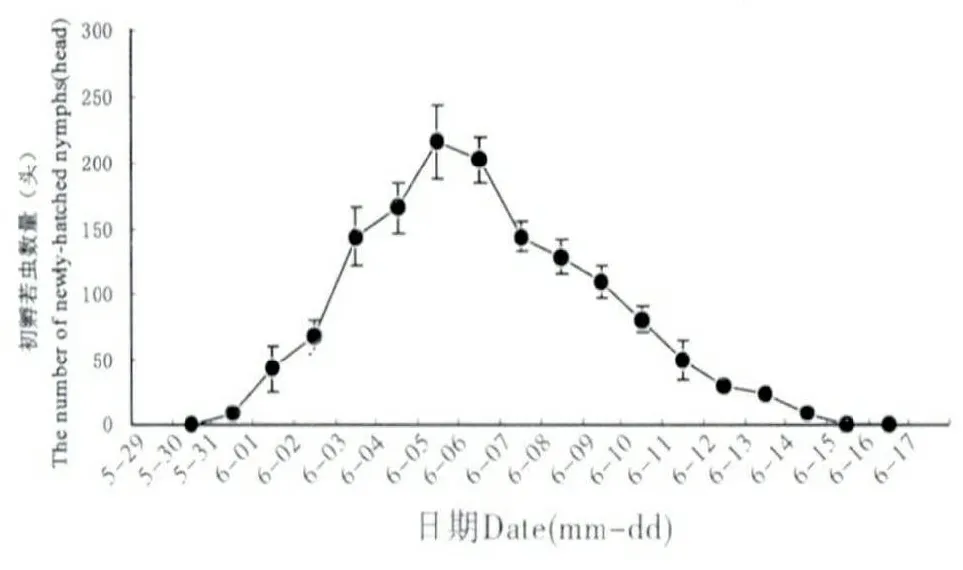
圖2 初孵若蟲逐日涌散量Fig.2 Daily incubation amount of nymph
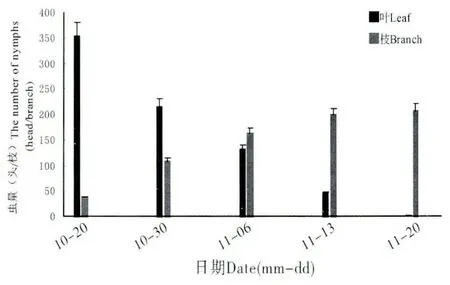
圖3 不同時期越冬若蟲在葉片和枝條的數量Fig.3 The number of the overwintering nymph in the leaves and branches in different periods
2.3 吐倫球堅蚧生活習性的研究
2.3.1 成蟲
在室內(溫度22℃~25℃,濕度25%~35%),雄成蟲羽化時,蟲體蠕動,頭胸部出現明顯的蛻裂線,蟲體依靠頭、胸、腹部的伸縮及附肢的活動逐漸將蛻裂線掙開,頭胸部最先脫出,脫出后的足將蛻皮向后蹬,同時腹部繼續伸縮活動,直至體軀全部脫出。脫出后雄成蟲繼續停在蠟質蛹殼內18~24 h,待腹末白色蠟質尾須長出后,再退出蠟質蛹殼,整個羽化過程地進行需時24~28 h,羽化時間集中在12∶00~14∶00時。
初羽化的雄成蟲十分活躍,爬動或飛翔,一般會沿枝條尋找合適的雌成蟲,發現雌成蟲后,雄成蟲通常先圍繞雌成蟲爬行,隨后雄成蟲將觸角向前伸展碰觸雌蟲并迅速爬向雌成蟲,同時雄成蟲將翅、觸角及腹末的一對白色長尾須豎起,腹部彎曲觸探雌成蟲臀裂處進行交尾,交尾時間一般持續1~2 分鐘。雄成蟲可多次交配,一般2~4次,最多6次,交尾后的雄蟲只能存活1~3 h,未交尾的雄蟲可存活2~3 d。雌成蟲只接受與一頭雄蟲交尾,未交尾的雌成蟲不產卵。雌雄性比約為1∶2(表2)。
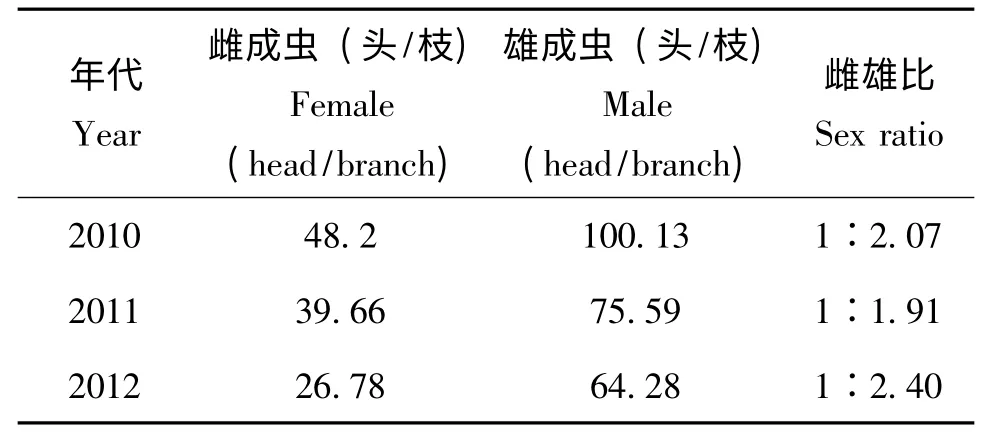
表2 吐倫球堅蚧雌雄性比Table 2 Sex ratio of Rhodococcus turanicus
雌成蟲排蜜初期,體型逐漸膨大,當雌成蟲體長增至3~4 mm 時,其每日排蜜量劇增,4月中旬為排蜜高峰期,此時正值雌成蟲孕卵期,整個排蜜期為22~24 d(圖4)。杏樹、桃樹、梨樹上的單雌累計排蜜量分別為35.95 mg、35.05 mg、29.68 mg,梨樹上的單雌累計排蜜量與其在杏樹和桃樹上的累計排蜜量差異顯著(圖5)。
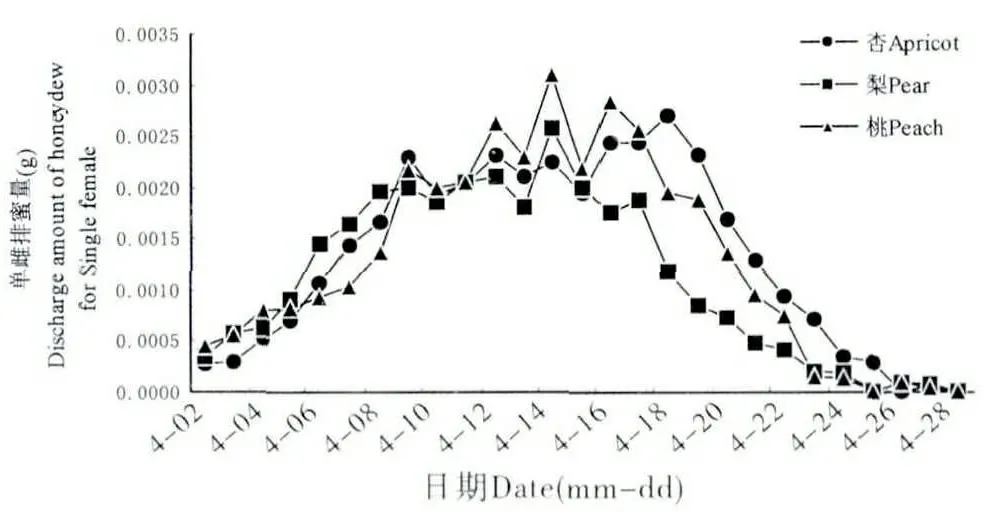
圖4 吐倫球堅蚧在不同寄主上的逐日排蜜量Fig.4 Daily honeydew excretion of Rhodococcus turanicus on different host
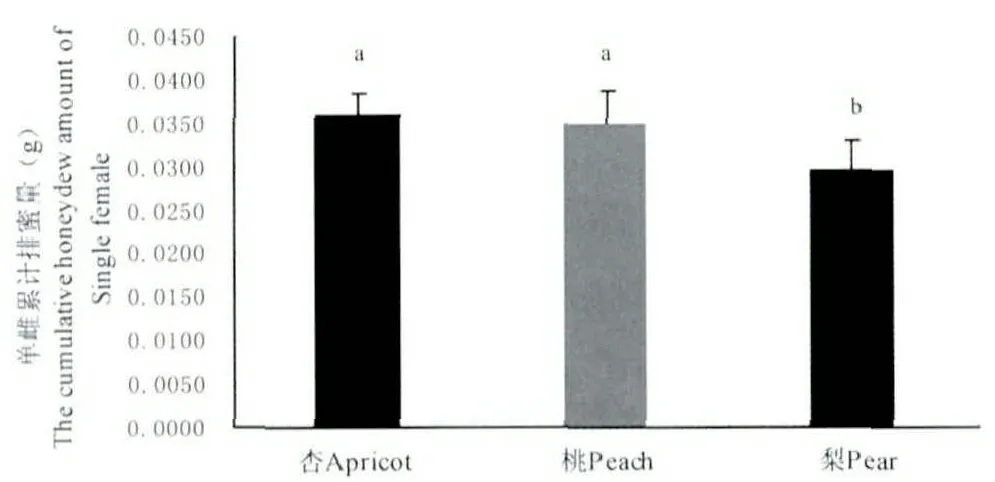
圖5 不同寄主上單雌累計排蜜量Fig.5 The cumulative honeydew amount of Single female on different host
2009~2011年,每年自4月開始當旬平均溫度達21.43±0.7℃時,為雌成蟲產卵高峰期。雌成蟲平均產卵量為1437.82 粒,單雌最大產卵量可達3500 粒(圖6)。
2010年,卵近孵化期,實測60頭雌成蟲的產卵量、自身體長和體重,以產卵量為因變量Y,以體長X1和體重X2為自變量進行逐步回歸分析(選入變量α≤0.01,剔除變量α≥0.05)(唐守正等,2009),所得回歸方程如下:
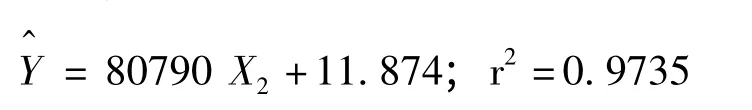
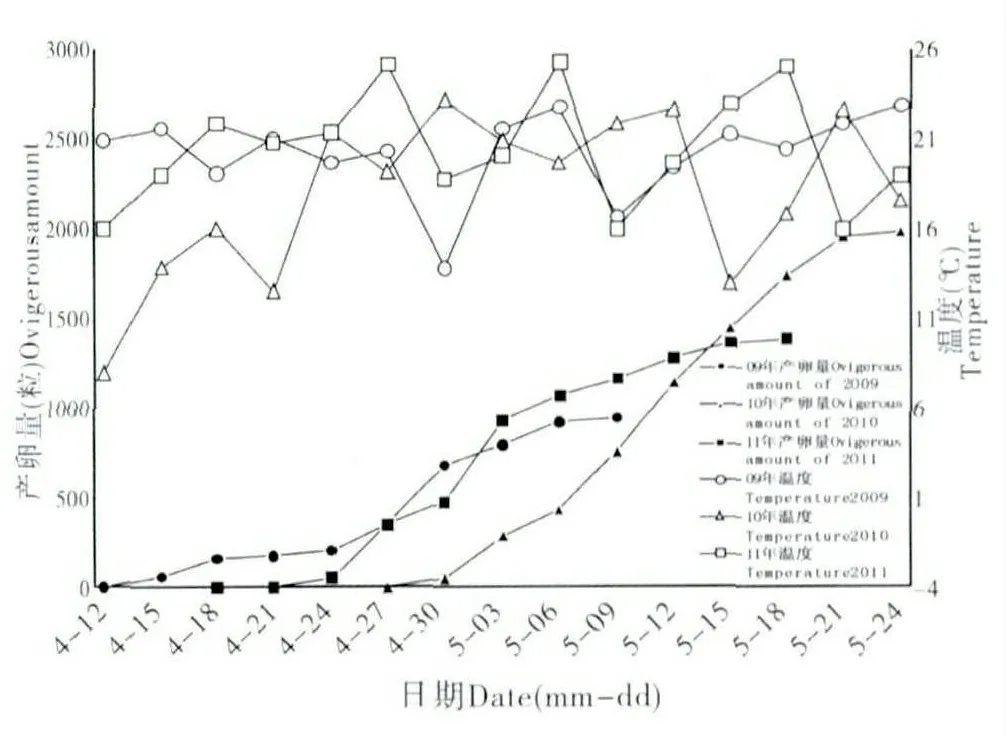
圖6 雌成蟲的產卵情況Fig.6 Spawning condition of female

圖7 殘差散點圖Fig.7 Residuals scatter diagram
2011年,在卵孵化期,以同樣的方法對24頭雌成蟲的產卵量及體重進行測量,并將體重代入方程求得產卵量預測值,見表3。用24 組數據對體重與產卵量的預測模型(回歸方程)進行驗證,采用置信橢圓檢驗出F=3.03<F0.05(2,22)=3.44,說明產卵量的預測值和實測值之間差異不顯著。因此,產卵量與體重的預測模型是適用的。
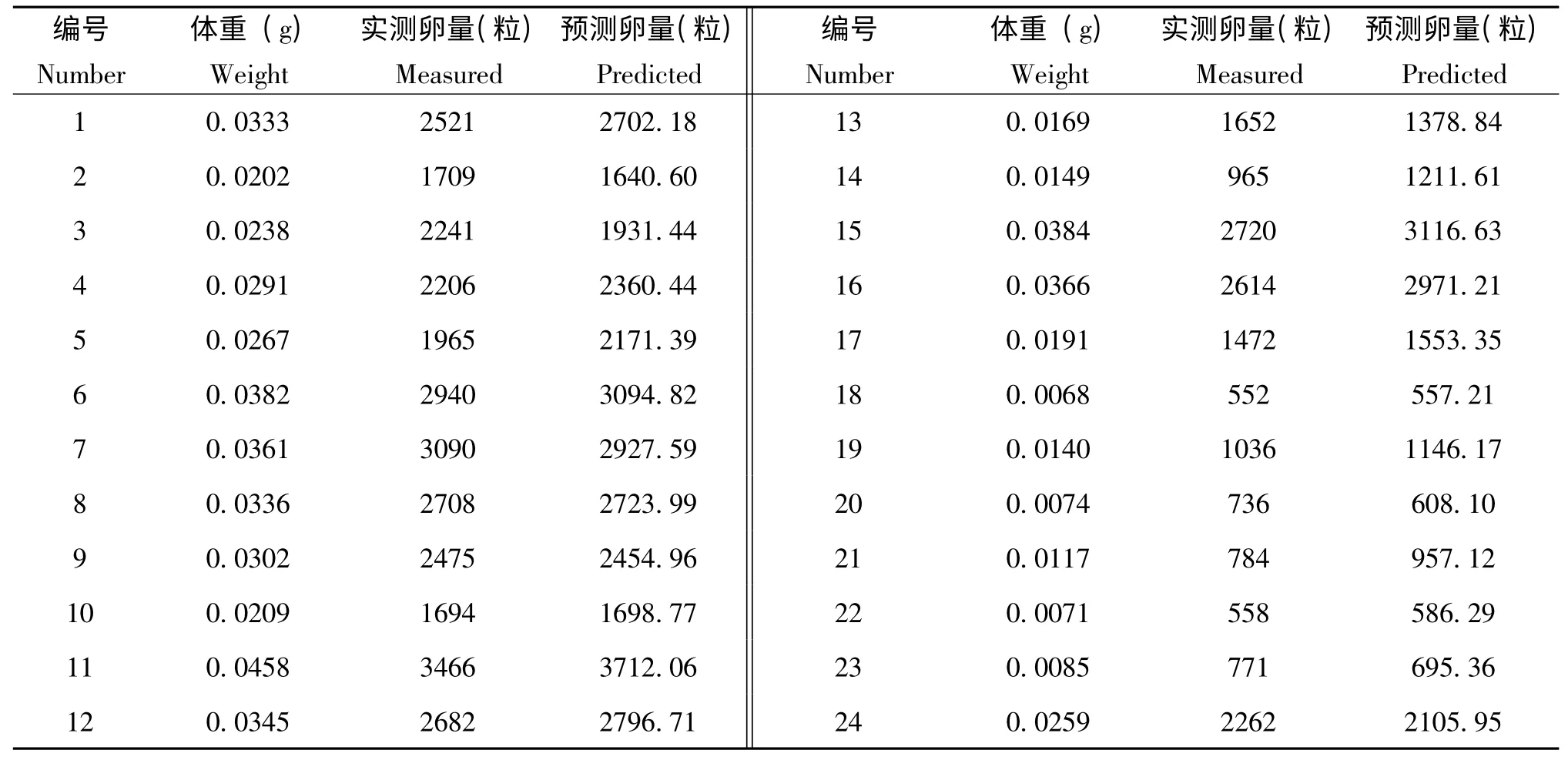
表3 產卵量的實測值及預測值Table3 Measured and predicted of ovigerous amount
2.3.2 卵
將卵置于恒溫與變溫條件下觀察其發育速率(圖8)。結果表明,卵的發育速率均隨溫度的升高而加快,經回歸分析,兩種條件下卵的發育速率與溫度呈直線關系,且相關性較高。根據公式將實測試驗數據計算得出恒溫與變溫條件下卵的發育起點溫度、有效積溫及表達式(表4)。
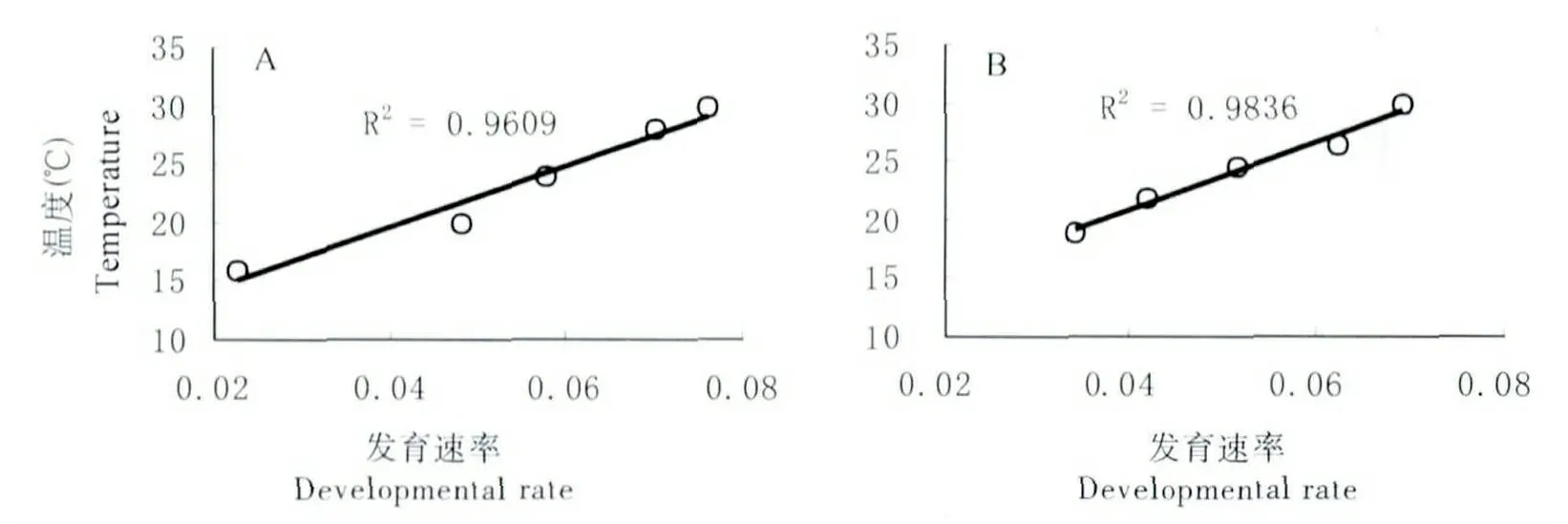
圖8 卵在恒溫(A)與變溫(B)條件下發育速率Fig.8 Developmental condition of eggs in constant temperature(A)and variable temperature(B)

表4 卵發育起點溫度及有效積溫Tabble4 Developmental threshold temperature and effective accumulated temperature of eggs
2.3.3 若蟲
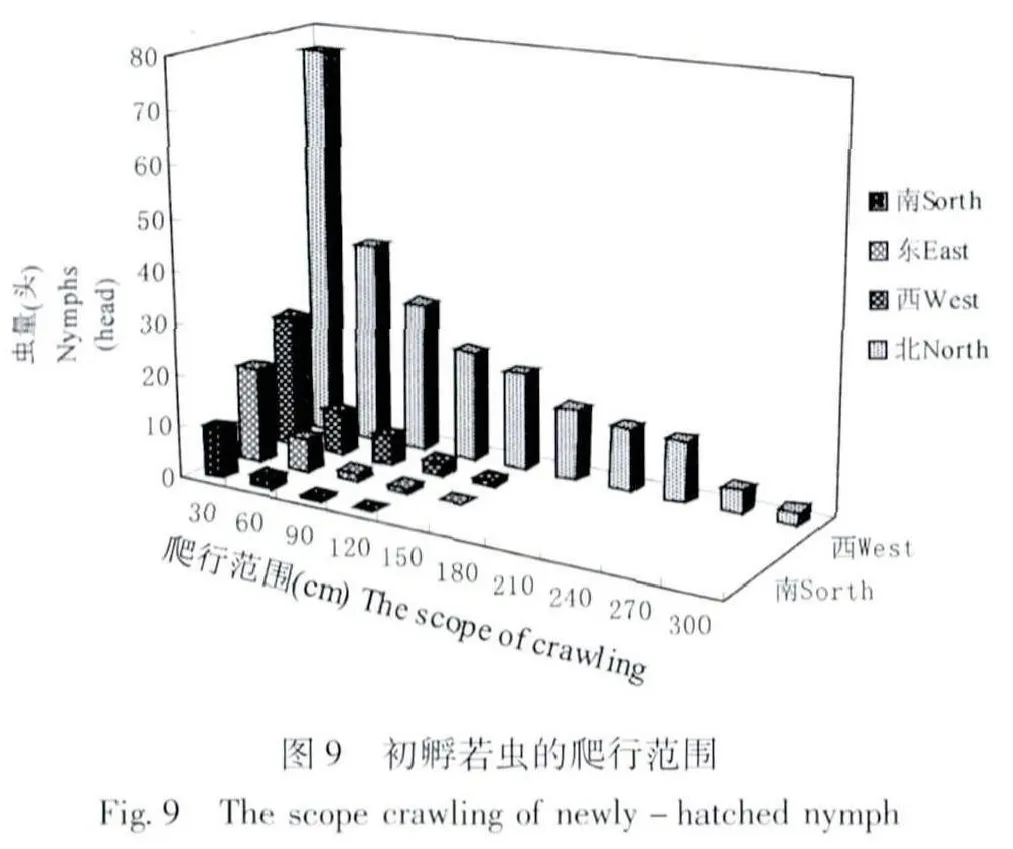
在室內(溫度22℃~25℃,濕度25%~35%),雌成蟲介殼內的若蟲孵化率為91.3%。初孵若蟲爬行范圍主要在距中心點60 cm 范圍內。在北面光線充足的條件下,初孵若蟲的爬行范圍可增至300 cm 處(圖9)。據實測初孵若蟲最大有效爬行距離達463 cm。
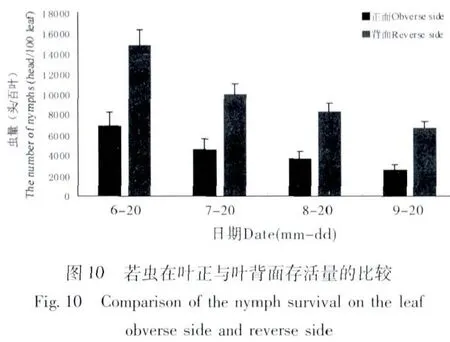
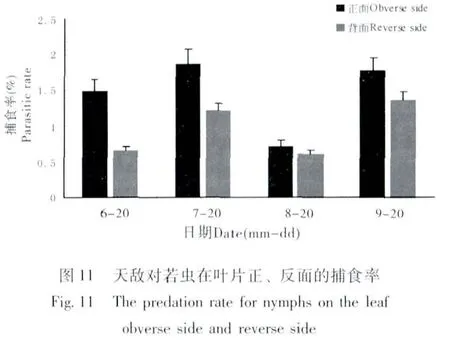
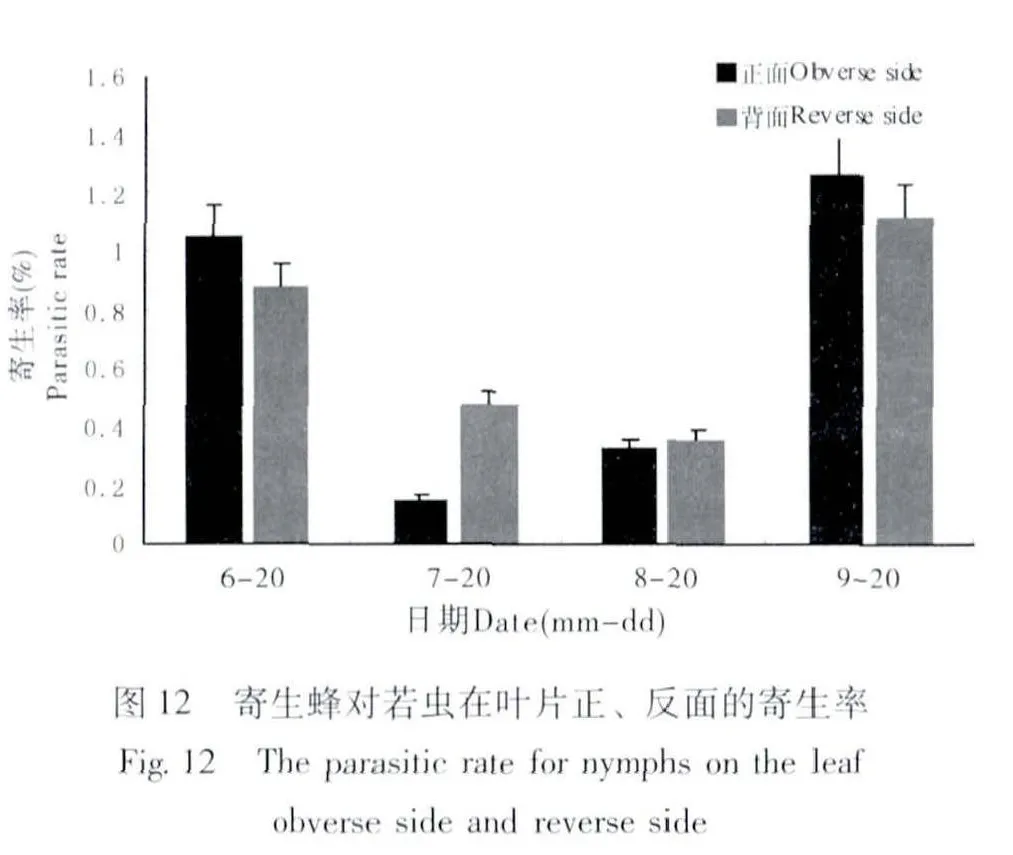
若蟲在葉片正面與背面取食的選擇性有很大差異,大多數若蟲選擇在葉片背面固定取食。若蟲在葉片正面與背面的存活數量見圖10,6月~9月葉片背面的若蟲數量均高于葉片正面,而若蟲總體數量為下降趨勢,7月和9月天敵對若蟲的捕食率相對較高,對葉片正面若蟲的捕食率高于葉片背面(圖11)。6月和9月寄生蜂對葉片正面若蟲的寄生率高于葉片背面(圖12)。
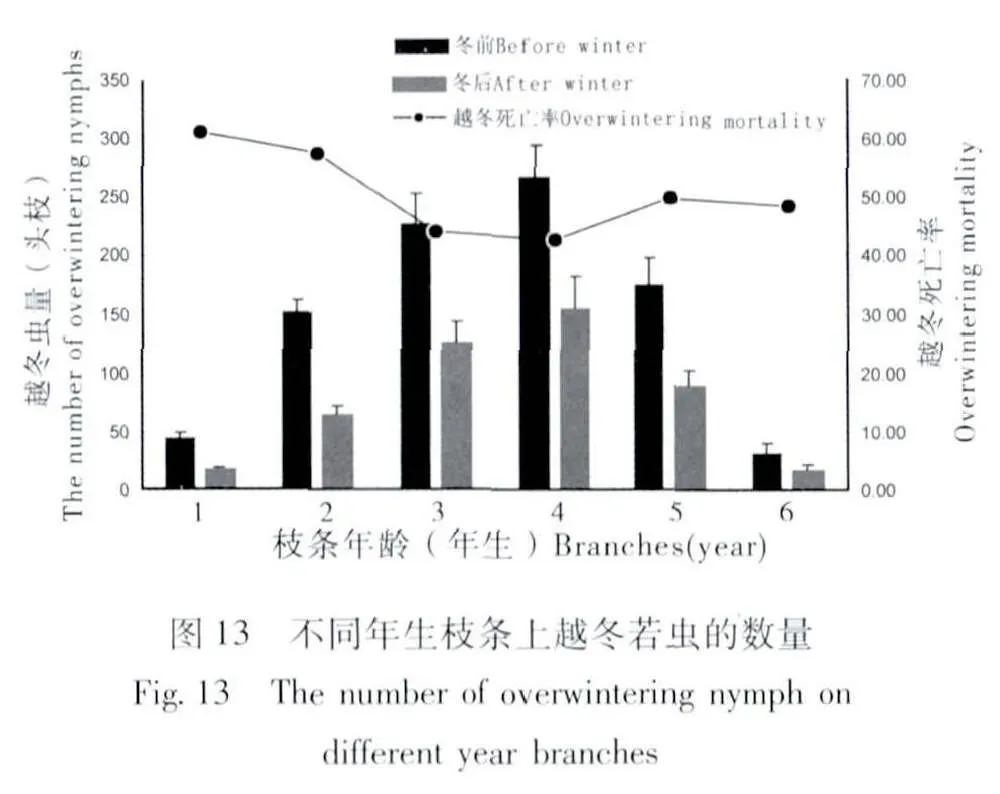
經田間調查,二齡若蟲在杏樹1~6年生枝條上越冬,主要聚集在3~4年生枝條上越冬。若蟲越冬死亡率可達46.37%,不同年生枝條上若蟲的越冬死亡率均有不同,若蟲在1年~2年生枝條上的越冬死亡率較高,為61.24%和57.52%;3~4年生枝條上的越冬死亡率相對較低,為44.23%和42.57%;5年~6年生枝條上若蟲越冬死亡率為49.75%和48.47%(圖13)。
3 結論與討論
2009~2012年,通過在室內和田間對吐倫球堅蚧生物學特性的系統研究。在新疆輪臺縣吐倫球堅蚧1年發生1代,以2齡若蟲固著于杏樹1年~6年生枝條上越冬,其中3~4年生枝條上越冬若蟲數量最多。吐倫球堅蚧的發育與杏樹物候及當地氣溫關系緊密,在田間的消長規律中以若蟲期時間最長、數量最大。吐倫球堅蚧雄蟲羽化集中在12∶00~14∶00,雌雄性比約為1∶2,雄蟲的存活時間很短,與雌成蟲交尾后即刻死亡,雌雄營兩性生殖。吐倫球堅蚧雌成蟲發育時期取食量在劇增,并大量排出蜜露,4月中旬為排蜜盛期,單雌最大排蜜量為35.95 mg。雌成蟲繁殖力強,平均產卵量為1437.82 粒,最高產卵量可達3500 粒,自4月開始當旬平均溫度達21.43±0.7℃時,為產卵高峰期,產卵量與體重相關性顯著,預測模型為^Y=80790 X2+11.874。卵的發育起點溫度為9.15±0.1593℃;有效積溫為290.17±4.876 DD。卵在雌成蟲體(介殼)的保護下孵化率達90%以上,初孵若蟲待介殼失水出現裂縫時涌散而出,6月上旬為涌散期。越夏若蟲主要選擇在葉片背面固定取食,這不僅可以減少天氣因素的影響,還可以降低天敵的捕食率和寄生率。越冬若蟲主要選擇3年、4年生枝條的背面固定越冬,可降低其越冬死亡率。
有研究表明與吐倫球堅蚧同一屬的沙里院褐球蚧Rhodococcus sariuoni Borchsenius,可行孤雌生殖或兩性生殖,在發生密度小時,主要以孤雌生殖,雄蟲很少;大發生時,雄蟲數量劇增,主要行兩性生殖(謝映平等,2006)。而本研究通過對吐倫球堅蚧發生密度不同的樣地進行調查,均未見孤雌生殖的現象。
關于蚧科昆蟲產卵量與體長、體重關系的研究報道較多(吳林等,1999;丁玉洲等,2001;唐燕平,2002),均以體長、體重分別于產卵量建立關系式。本研究采用逐步回歸的方法對吐倫球堅蚧產卵量與體長、體重進行分析得出,產卵量與體重極相關。通過殘差分析和置信橢圓檢驗預測模型^Y=80790 X2+11.874 是可行的,所以在實際生產中,可根據雌成蟲體重預測產卵量。但自然環境下導致模型的預測產生差異的因素很多,所以預測模型的驗證與調整仍需要進一步的研究。
References)
Wang ZQ,2001.Fauna Sinica(Insecta Vol.22).Beijing:Science Press.385-387.[王子清,2001.中國動物志 昆蟲綱 第二十二卷.北京:科學出版社.385-387]
Liang AP,2005.A proposal to stop using the insect order name“Homoptera”.Chinese Bulletin of Entomology,42(3):332-336.[梁愛萍,2005.關于停止使用“同翅目Homoptera”目名的建議.昆蟲知識,42(3):332-336]
Tang FD,1991.Coccidea of China.Taiyuan:Shanxi University United Press.323-325.[湯祊德,1991.中國蚧科.山西:山西高校聯合出版社.323-325]
Luo XS,Chen XS,Fan CL,Yu XZ,1989.Research progress of apricot.Deciduous Fruits,(4):9-13.[羅新書,陳學森,范春林,于希志,1990.杏樹研究進展.落葉果樹,(4):9-13]
Cheng WD,Lv GH,Li L,Liang X,2006.Integrated utilization and industrialization of Xinjiang little white apricot.Transactions of the Chinese Society of Agricultural Engineering,22(9):241-244.[程衛東,呂國華,李琳,梁霞,2006.新疆小白杏資源的綜合利用及其產業化.農業工程學報,22(9):241-244]
Zhang ZL,Zhao L,Zhang LY,Li JR,Li BJ,2010.Study on the efficacy trials of several pesticides controlling Rhodococcus turanicus Arch.nymph.Journal of Xinjiang Agricultural University,33(4):326-328.[張滋林,趙莉,張魯豫,李軍如,李炳均,2010.幾種藥劑防治吐倫球堅蚧若蟲的藥效研究.新疆農業大學學報,33(4):326-328]
Babayan GA,1970.New data on the morphological and biological characteristics of the Turanian scale Rhodococcus turanicus Arch.(Homoptera,Coccoidea) in the Armenian SSR.Entomologicheskoe Obozrenie,49(4):729-735.
Zhu XF,Abudukeyoumu T,Xu BQ,Tuerxun N,Feng Y,Yang S,Study on the spatial distribution pattern and sampling technique of Rhodococcus turanicus Arch.in apricot orchard.Journal of Environmental Entomology,31(2):102-106.[朱曉鋒,阿布都克尤木,徐兵強,吐爾遜那依,馮洋,楊森,2009.杏樹吐倫球堅蚧空間分布型及抽樣技術研究.環境昆蟲學報,31(2):102-106]
Yang S,Tuerxun N,Abdukeyimu,Xu BQ,2008.Preliminary study on predation of Philodromus cespitum(Walckener) etc.to Rhodococcus turanicus Arch.Acta Arachnologica Sinica,17(1):43-45.[楊森,吐爾遜娜依,阿不都克尤木,徐兵強,2008.草皮逍遙蛛等天敵對吐倫球堅蚧捕食作用的初步研究.蛛形學報,17(1):43-45]
Fan Y,Zhao L,Zhang LY,Ren JL,Aimaijiang 2013.Control effect and bioassay of several insecticides on Rhodococcus turanicus.Plant Protection,39(1):171-177.[范毅,趙莉,張魯豫,任金龍,艾買江,2013.幾種藥劑對吐倫球堅蚧的毒力及田間防效.植物保護,39(1):171-177]
Mu JY,Xu HF,Li HG,1997.Insect Ecology and Pest Forecast.Beijing:China Agricultural Science and Technology.75-113.[牟吉元,徐洪富,李火茍,1997.昆蟲生態與害蟲預測預報.北京:中國農業科技社.75-113]
Kang SZ,Lang KJ,Li HK,2009.Calculation of Statistical and Mathematical Model.Beijing:Science Press.75-94.[唐守正,郎奎建,李海奎,2009.統計與生物數學模型計算.北京:科學出版社.75-94]
Xie YP,Xue JL,Zheng LY,2006.Wax Secretions of Soft Scale Insect Their Ultrastructure ﹠ Chemical Composition.Beijing:China Forestry Publishing House.74-75.[謝映平,薛皎亮,鄭樂怡,2006.蚧科昆蟲的蠟泌物超微結構和和化學成分.北京:中國林業出版社.74-75]
Wu L,Yang CC,Hong GP,Fan XL,Ren JM,1999.Study on forecast of ovipositing amount and occurrence time of Ceroplastes ceriferus.Forest Research,12(3):321-324.[吳林,楊春材,洪國朋,范賢龍,任建敏,1999.角蠟蚧產卵量與卵期預測預報的研究.林業科學研究,12(3):321-324]
Ding YZ,Fang GF,Wang CQ,Fang TQ,Wang CH,2001.Study on the relations between Kernesnawai Kuwana Form indicatrixs and occurrence amount.Journal of Anhui Agricultural University,28(1):77-81.[丁玉洲,方國飛,王成慶,方泰全,汪成海,2001.栗絳蚧形態生物指標與發生量關系研究.安徽農業大學學報,28(1):77-81]
Tang YP,Liu Y,2002.Forecast of the oviposition amount and period of Eulecanium kuwanai.Forest Pest and Disease,12(2):9-12.[唐艷平,劉遠,2002.皺大球蚧產卵與卵期預測預報的研究.中國森林病蟲,12(2):9-12]
