貯藏溫度對山竹果皮木質化相關酶活性和果肉腐爛的影響
2013-03-06 02:32:11于立梅劉朝霞陳海光
食品科學 2013年18期
楊 敏,于立梅,*,劉朝霞,陳海光
(1.仲愷農業工程學院輕工食品學院,廣東 廣州 510225;2.廣東檢驗檢疫技術中心,廣東 廣州 510623)
貯藏溫度對山竹果皮木質化相關酶活性和果肉腐爛的影響
楊 敏1,于立梅1,*,劉朝霞2,陳海光1
(1.仲愷農業工程學院輕工食品學院,廣東 廣州 510225;2.廣東檢驗檢疫技術中心,廣東 廣州 510623)
以海南山竹果為原料,研究不同貯藏溫度(30、4℃)對采后山竹果皮木質化相關酶活性和果肉腐爛的影響,探討山竹果皮木質化效應。結果表明,不同貯藏溫度對山竹果皮木質化效應程度不同,低溫處理初期有冷害現象,導致酶活性和木質素含量增強,但隨著貯藏期延長至末期,常溫處理的木質素含量比同低溫處理的木質素含量增加14.3%。常溫貯藏山竹果實PAL、POD、PPO及CAD活性比低溫增加顯著。常溫處理和低溫處理的山竹PAL活性峰值分別為170.40U/(g·h)和180.12U/(g·h),在貯藏后期(6~15d),各組處理的活性均呈現持續下降的趨勢。低溫處理的腐爛指數低于常溫處理,15d常溫處理的腐爛率增加了78.89%,低溫處理的腐爛率增加了32.22%,可能是低溫處理延緩果殼木質化相關酶活性,減少細胞代謝物質含量的積累,保持較低的呼吸強度,從而可以抑制腐爛。
山竹果;木質化;酶活性; 貯藏溫度
山竹(Garcinia mangostana L.)為藤黃科(Guttiferae)山竹子屬植物,又稱倒捻子、鳳果或莽吉柿,原產于馬來西亞群島,現分布于印度尼西亞、菲律賓、緬甸、馬來西亞、越南、泰國、錫蘭、中國等地。山竹味偏酸,補益作用較強,具有“熱帶果后”之稱。果實可食部分占29 %~45%,果皮表皮紫褐色,其余部分淺白色,厚度為0.8~1.1cm,占單果鮮質量的52%~68%[1-2]。由于山竹生產的季節性較強,采收后極易出現果皮木質化、褐變和腐爛變質,影響著山竹的食用品質和商品價值。木質化的出現與果殼組織細胞壁中木質素的沉積有關,而木質素的生成由植物細胞中的一些酶來調控,如多酚氧化酶(PPO)、苯丙氨酸解氨酶(PAL)、肉桂醇脫氫酶(CAD)、過氧化物酶(POD)及其同工酶等。在植物體內,PAL是在木質化進程中將L-苯丙氨酸轉化成反式肉桂酸的關鍵酶,而過氧化物酶參與將羥基肉桂醇聚合成木質素[3]。對于采后山竹果實木質化及硬度的增加是否與相關酶的活性變化相關,果皮木質化對果肉的影響均未見報道。因此本實驗以海南山竹果實為材料,主要探討常溫(30℃)和低溫(4℃)處理對山竹果殼木質化相關酶活性和果肉腐爛的影響,初步探討山竹采后果品木質化對果肉的影響,為山竹果殼木質化形成機理和果肉保鮮提供理論依據。
1 材料與方法
1.1 材料與試劑
實驗原料山竹采自海南五指山市毛道鄉山竹種植園。果實于正常食用成熟度采收,采后當天運回學院實驗室。
PVP(聚乙烯吡咯烷酮) 張家港科悅精細化工有限公司;NADP(輔酶Ⅱ) 美國Sanland 公司;磷酸氫二鈉、磷酸二氫鈉、反式肉桂酸(cinnanic acid) 美國Alfa Aesar公司;β-巰基乙醇、硼砂、硼酸、鹽酸、L-苯丙氨酸、三羥甲基氨基甲烷(Tris)等均為國產分析純試劑。
1.2 儀器與設備
DU-730型紫外-可見分光光度計 日本島津分析儀器廠;恒溫水浴鍋 廣東環凱微生物科技有限公司;電熱恒溫培養箱 上海索譜儀器有限公司;HY-Z型調速振蕩器 常州國華電器有限公司;高速離心機 長沙湘儀離心機儀器有限公司;KQ5200超聲波清洗器 昆山市超聲儀器有限公司。
1.3 方法
選取無機械損傷、大小均勻、成熟度一致的山竹果實,分裝于0.02mm聚乙烯袋,用橡皮筋繞兩道扎袋口而不密封,分別于30℃(常溫)和4℃(低溫)貯藏。每份35個,重復5次。每3d每組隨機取1份,將果肉和果皮分離,果皮粉碎后進行相關指標的測定。
1.3.1 木質素含量的測定
參照朱海英等[4]的方法。
1.3.2 粗纖維含量的測定
參考黃萍等[5]的方法。
1.3.3 PPO活性的測定
參照Galeazzi[6]和Zauberman[7]等的方法。樣品液制備:取山竹果殼4.0g,加入20mL 0.1mol/L pH6.8預冷的磷酸鈉緩沖液,在冰浴條件下研磨,勻漿后將勻漿液全部轉移入離心管中。在12000r/min、4℃條件下離心20min,收集上清液即為PPO提取液,低溫保存備用。
PPO活性的測定:在6.7mL的反應體系中,加入4.4mL 0.1mol/L pH6.8的磷酸緩沖液,2.0mL 0.1mol/L的鄰苯二酚(0.1mol/L pH6.8的磷酸緩沖液溶解)和0.3mL酶提取液,立即混合后在420nm波長處比色,然后迅速放在37℃的水浴中反應,酶提取液加入后開始計時,測定在420nm波長處反應11min吸光度的變化,空白為0.1mol/L的磷酸緩沖液(pH6.8)。以每克樣品(鮮質量)每分鐘吸光度變化0.001為一個酶活力單位(U),酶活性以U/g表示。
1.3.4 PAL活性的測定
參照Koukol[8]和王敬文[9]等的方法,稍有變動。樣品液制備:稱取1.0g山竹果殼,置于研缽中,加入20mL 0.lmol/L pH8.8預冷的硼酸-硼砂提取緩沖液(含40g/L PVP、5mmol/L β-巰基乙醇),在冰浴條件下研磨成勻漿。將勻漿液全部轉入到離心管中,于4℃條件下,12000r/min離心20min,收集上清液,即為粗酶提取液,低溫保存備用。
PAL活性的測定:l.0mL 0.05mol/L L-苯丙氨酸,2.9mL 0.1mol/L pH8.8硼酸緩沖液,0.1mL樣品液。以煮沸5min的失活酶樣品液作為對照,以加0.1mL的硼酸緩沖液為空白對照。混勻后,將其置于37℃水浴中保溫60min。保溫結束后,立即向反應管中分別加入0.1mL 6.0mol/L的HCl終止反應,如有沉淀應離心除去。用空白對照調零,分別測定樣品管和對照管中溶液在波長290nm處的吸光度(A1和A0),以每小時每克樣品(鮮質量)酶促反應體系吸光度A290變化0.01為1個PAL活性單位(U),酶活性以U/g表示。
1.3.5 CAD活性的測定
參照Goffner等[10]的方法。樣品液制備:取山竹果殼3.0g,加入15mL 0.1mol/L pH8.8預冷的Tris-HCl提取緩沖液(含40g/L PVP、5mmol/L β-巰基乙醇),在冰浴條件下研磨,勻漿后在12000r/min、4℃條件下離心20min,收集上清液即為樣品液,低溫保存備用。
CAD活性的測定:酶液反應體系為:2.8mL 0.1mol/L pH8.8 Tris-HCl緩沖液、1mL 2mmol/L的NADP、1mL 1mmol/L的反式肉桂酸、0.1mL樣品液,以煮沸5min的樣品液作對照,以加0.1mL緩沖液為空白對照;將其置于37℃保溫30min。結束保溫后,立即向反應管中分別加入0.1mL 6.0mol/L的HCl終止反應,如有沉淀應離心除去。用空白對照調零,分別測定樣品管和對照管中溶液在波長340nm處的吸光度(A1和A0),以每小時每克樣品(鮮質量)酶促反應體系吸光度A340變化0.001為1個CAD活性單位(U),酶活性以U/g表示。
1.3.6 POD活性的測定
參照Hammerschmidt[11]、Kar[12]和Srivastava[13]等的方法。樣品液制備:取山竹果殼4.0g,加入20mL 0.1mol/L pH6.8預冷的磷酸鈉提取緩沖液(含40g/L PVP、5mmol/L β-巰基乙醇),在冰浴條件下研磨,勻漿后以12000r/min、4℃條件下離心20min,收集上清液即為樣品液,低溫保存備用。
POD活性的測定:6.2mL反應液中含有0.3mL 4.0%愈創木酚、0.3mL 0.46%過氧化氫、5.0mL磷酸鈉緩沖液(pH6.8)和0.6mL酶液,從加酶液后1min開始記錄每分鐘反應體系在470nm波長處的吸光度,連續測定6min,以每克樣品(鮮質量)每分鐘吸光度變化0.001為1個POD活性單位(U),酶活性以U/g表示。
1.3.7 腐爛指數的測定
參考芮懷瑾等[14]的方法,將果實按腐爛面積大小分為4級。0級,無腐爛;1級,腐爛面積小于果實面積的10%;2級,腐爛面積占果實面積的10%~30%;3級,腐爛面積大于果實面積30%。按下式計算腐爛指數:腐爛指數/% =(∑(級別×該級果數)/總果數)×100
1.3.8 數據分析方法
每個實驗測定重復3次,獲得平均值和標準偏差。應用SPSS 19.0軟件對所有試驗最終數據進行方差分析,采用Duncan’s新復極差法進行方差分析,檢驗差異顯著性(P<0.05表示差異顯著,P<0.01表示差異極顯著)。
2 結果與分析
2.1 不同溫度貯藏山竹果殼木質素含量的變化
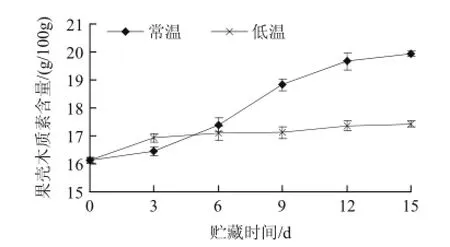
圖1 不同溫度貯藏山竹果殼木質素的變化Fig.1 Changes of lignin content during storage of mangosteen pericarp at different temperatures
木質素作為植物次生代謝產物,是構成細胞壁次生結構的主要成分。山竹果殼木質素含量的上升是衡量山竹果實貯藏期木質化敗壞的最重要的生理指標之一。如圖1 所示,在整個貯藏期,山竹果殼木質素含量隨著貯藏期的延長均呈上升趨勢。在0~3d,低溫處理的木質素含量高于常溫處理,可能是冷害作用加速了木質化,持續6d左右,但差異不顯著。在6~15d貯藏期,低溫處理的木質素含量始終低于常溫處理,0d時,兩種處理組的木質素含量均為16.13g/100g,當貯藏15d后,常溫處理的木質素含量比同期低溫處理的木質素含量增加14.3%。經差異顯著性檢驗分析,在貯藏第9~15天時,常溫與低溫處理的木質素含量之間的差異性顯著(P<0.05)。上述結果表明,低溫貯藏在一定程度上能抑制山竹果殼木質素的形成,延緩果殼的木質化。徐俐等[15]的研究表明,竹筍低溫貯藏時的呼吸強度明顯低于常溫貯藏,進一步說明了室溫條件下鮮竹筍的呼吸作用比低溫條件下旺盛,與本實驗結果類似。
2.2 不同溫度貯藏山竹果殼粗纖維含量的變化
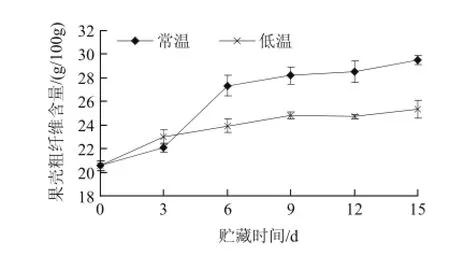
圖2 不同溫度貯藏山竹果殼粗纖維含量的變化Fig.2 Changes of coarse fiber content during storage of mangosteen pericarp at different temperatures
如圖2所示,山竹果殼粗纖維含量隨著貯藏期的延長均呈上升趨勢。在貯藏0~3d時,低溫處理的粗纖維含量高于常溫,貯藏3~6d,常溫處理的粗纖維急速增加,其中在第9天時達到了穩定值,常溫和低溫處理的果殼粗纖維含量分別為29.50g/100g和25.37g/100g,常溫處理比低溫處理的含量高4.13%。經差異顯著性檢驗分析,6d后常溫處理與低溫之間差異性顯著(P<0.05)。上述結果表明,常溫貯藏條件下的山竹果殼粗纖維生成速率較低溫處理快,低溫貯藏在一定程度上能抑制山竹果殼粗纖維的形成,延緩果殼的衰老。經相關性分析,山竹果殼粗纖維含量與木質素含量相關系數為0.851,表明山竹果殼木質化除了表現為木質素含量的增加,也伴隨著粗纖維含量的增加。劉尊英等[16]研究結果表明,降低貯藏溫度能顯著抑制采后豌豆苗纖維化速率,并有效抑制可溶性糖和蛋白質等營養成分的下降及因葉綠素降解而引起的黃化。
2.3 不同溫度貯藏山竹果殼PPO活性的變化
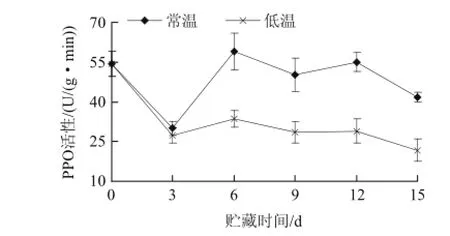
圖3 不同溫度貯藏山竹果殼PPO活性的變化Fig.3 Changes of PPO activity during different temperature storage of mangosteen pericarp at different temperatures
從圖3可知,山竹果殼在貯藏期內其多酚氧化酶活性均呈現比較一致的變化,山竹果殼的PPO活性在0~3d內迅速下降,而后3~6d內又呈現上升趨勢。6~9d緩慢下降,其中12d活性稍高,差異不顯著,總體趨勢下降,經差異顯著性檢驗分析,第3天后,各處理間差異顯著(P<0.05)。原因可能為酶和底物接觸,又有氧氣存在,就會發生木質化。伴隨著木質化的進行,山竹果殼表面變得致密堅硬,果殼木質層的不斷加厚也使得果實內外環境被逐漸阻隔開來。導致缺少作為反應條件之一的氧氣,PPO活性逐漸下降。其次由于PPO作為植物氧化酶系中得一種內源呼吸酶,其活性與新陳代謝密切相關:隨著貯藏時間推移,山竹代謝速率的下降導致PPO活性隨之降低。以上結果表明:低溫處理能較有效降低山竹果殼中PPO活性。吳振先等[17]報道,荔枝PPO活性在果實貯藏過程中也呈現出先降低后升高,最后又降低的變化趨勢。2.4 不同溫度貯藏山竹果殼PAL活性的變化
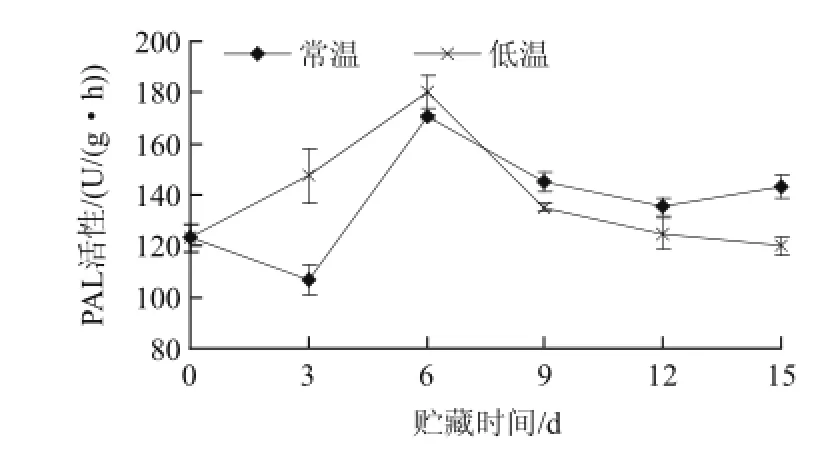
圖4 不同溫度貯藏山竹果殼PAL活性的變化Fig.4 Changes of PAL activity during storage of mangosteen pericarp at different temperatures
苯丙氨酸解氨酶(PAL)是連接初級代謝和苯丙烷類代謝、催化苯丙烷類代謝第一步反應的酶,也是木質化沉積最重要的酶類。從圖4可知,在0~3d內,常溫處理的PAL活性稍下降,隨后在3~6d內上升至峰值,而低溫處理的PAL活性在0~6d急劇上升,可能是低溫冷害誘導PAL活性升高,常溫處理和低溫各組處理的峰值分別為170.40U/(g·h)和180.12U/(g·h),在貯藏后期(6~15d),各組處理的活性均呈現持續下降的趨勢。經差異顯著性檢驗分析,貯藏第3d時低溫處理和常溫處理的差異性顯著(P<0.05),低溫處理的山竹果殼PAL活性在初期明顯高于常溫處理的山竹果殼,低溫冷害作用加速了木質化沉積。鄭永華等[18]研究認為低溫誘導枇杷果實PAL活性上升,低溫條件下果實PAL活性一直處于較高水平,促進果實木質素的合成,導致冷害癥狀的出現。
2.5 不同溫度貯藏山竹果殼CAD活性的變化
肉桂醇脫氫酶(CAD)是植物木質素合成途徑中重要的限速酶,也是木質素單體合成最后階段的一種酶,從圖5得知,兩種處理的山竹果的CAD活性呈現不同的變化。在0~3d內,低溫和常溫各組CAD活性都出現上升趨勢,低溫顯著高于常溫,可能是低溫誘導CAD活性。在3~6d內,低溫活性下降,常溫處理的CAD活性增長非常迅速,常溫從375U/(g·h)增長到579.67U/(g·h)、低溫3d達到峰值為446U/(g·h)。6~15d 低溫處理D的CAD活性變化甚微。9~15d,常溫處理的CAD活性呈緩慢增長趨勢。經差異顯著性檢驗分析,不同的貯藏期,兩種處理的CAD活性均差異顯著(P<0.05)。經相關性分析,山竹果殼的CAD活性與木質素含量的相關系數為0.397,表明CAD能促進山竹果殼木質素含量的增加,并在山竹果殼木質化進程中起到了極其重要的作用。吳錦程等[19]研究得出,枇杷果實貯藏過程中溫度對CAD活性影響較大,12℃條件下貯藏的果肉CAD活性顯著低于4℃貯果的活性水平,低溫誘導了CAD活性的較大增加,從而促進4℃條件下果肉木質素的生成。本實驗結果表明不同生理狀態下不同實驗材料CAD的生理功能有所差別。
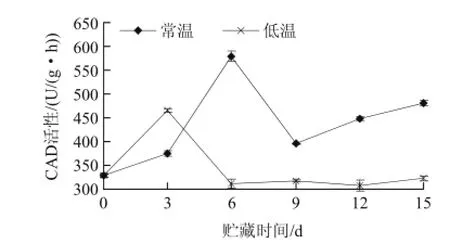
圖5 不同溫度貯藏山竹果殼CAD活性的變化Fig.5 Changes of CAD activity during storage of mangosteen pericarp at different temperatures
2.6 不同溫度貯藏山竹果殼POD活性的變化
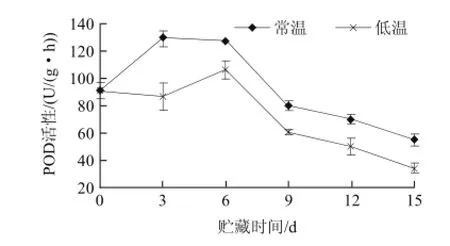
圖6 不同溫度貯藏山竹果殼POD活性的變化Fig.6 Changes of POD activity during storage of mangosteen pericarp at different temperatures
過氧化物酶(POD)是木質素生物合成中最后一步的關鍵酶,在H2O2參與下催化木質素單體聚合成木質素,參與木質素在細胞壁的聚合,提高組織木質化程度。如圖6所示,兩種處理的樣品POD活性呈現相似的變化趨勢,貯藏期間的變化為0~6d POD增加,可以認為是采后逆境的脅迫所致,誘發細胞產生來抵御活性氧的傷害。隨著木質化進程,第6~15天POD活性下降。通過木質素相關性分析,0~6d,POD活性與組織木質素含量呈顯著正相關。許曉春等[20]發現,荔枝果皮POD活性貯藏初期有一定程度的增加,隨著果皮褐變的開始,活性減小, 席玙芳等[21]在相關研究中發現POD活性與組織木質素含量呈顯著正相關。
2.7 不同貯藏溫度對山竹果肉腐爛指數的變化
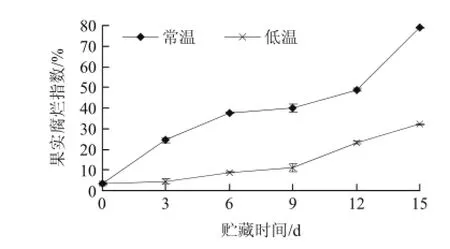
圖7 不同貯藏溫度山竹果肉腐爛指數的變化Fig.7 Changes of decay index during storage of mangosteen pulp at different temperatures
果蔬采后腐爛是一個全球性的問題,在世界范圍內約有25%的果蔬產品因腐爛變質而不能利用,在山竹果為期15d的貯藏過程中,山竹果肉的色澤呈現乳白色→白色、黃斑點→大面積的黃色的變化,果肉也不同程度地發生自溶現象。采后腐爛一個主要原因是隨呼吸作用增強而造成乙烯濃度過高。山竹果肉產生自溶現象,果肉產生自溶現象是由于本身降解酶的作用,破壞了果肉表面的防腐膜,使高糖果汁外溢,引起各種微生物的滋生而加劇了整個果肉的敗解。果肉自溶現象除了與果肉本身降解酶和微生物侵染有關外,還因為果肉中果膠含量及存在方式發生了改變和淀粉酶降解淀粉。由圖7可以看出,不同處理的山竹果腐爛指數均隨著貯藏時間的延長而逐漸增大,但低溫處理的腐爛指數低于常溫處理,15d常溫處理的腐爛率增加了78.89%,低溫處理的腐爛率增加了32.22%,可能是低溫處理延緩果殼木質化相關酶活性,減少細胞代謝物質含量的積累,保持較低的呼吸強度,從而可以抑制腐爛,其中以低溫處理效果最佳。
3 結 論
果蔬組織的木質化由一系列相關酶類控制。多酚氧化酶(PPO)、苯丙氨酸解氨酶(PAL)、肉桂醇脫氫酶(CAD)和過氧化物酶(POD)都參與酚類物質代謝,是木質化過程中的關鍵酶類。PPO通過參與酚類物質(如綠原酸、香豆素等)的氧化過程,而促進木質素的形成;PAL是木質素生物合成中的限速酶,能催化苯丙氨酸轉化為肉桂酸;CAD是苯丙烷類代謝中催化木質素前體香豆醛向香豆醇轉化,它是木質素代謝途徑中的一個重要環節;而POD則在木質素生物合成的最后一步中通過催化H2O2分解而使木質素單體發生聚合反應形成木質素。本實驗中,隨著貯藏期的延長,山竹果實的硬度逐漸上升,果實中PAL、POD、PPO及CAD活性表現為不同時期的增加趨勢,同時伴隨著木質素含量的增加。在促進山竹果殼木質化過程中,CAD起到主導作用,PPO與PAL也能促進木質素的形成積累,這3種酶與木質素含量的相關性均表現為正相關。低溫處理雖在貯藏初期有一定的冷害失調,但在整個貯藏期低溫能顯著抑制木質化相關酶活性的上升,抑制木質素含量的增加,因此能較好地保持山竹果實的新鮮度,延緩出現木質化現象和果肉腐爛。
[1] 楊連珍. 山竹子[J]. 熱帶農業科學, 2002, 8(4): 60-66.
[2] 劉全儒, 于明. 幾種熱帶果實的食用部[J]. 生物學通報, 2003, 38(1): 10-13.
[3] 潘瑞熾. 植物生理學[M]. 4版. 北京: 高等教育出版社, 2001: 133-135.
[4] 朱海英, 李人圭, 王隆華, 等. 絲瓜果實發育中木質素代謝及有關導管分化的生理生化研究[J]. 華東師范大學學報, 1997(1): 87-94.
[5] 黃萍, 鐘新民, 李必元, 等. 雪菜中粗纖維含量的測定[J]. 吉林農業科學, 2011(1): 45-47.
[6] GALEAZZI M A M, SGARBIERI V, COSTANTINIDES S M. Isolation, purification and physiocochemical characterization of polyphenol oxidase from dwarf variety of banana (Musa carendishii) [J]. Journal of Food Science, 1981, 46(1): 150-155.
[7] ZAUBERMAN G, FUCHS Y, ROT I, et al. Chilling injury, peroxidase, and cellulase activities in the peel of mango fruit at low temperature[J]. Hort Science, 1988, 23(4): 732-733.
[8] KOUKOL J, CONN E E. The metabolism of aromatic and properties of the Phenylalanine deaminase of Hordeum vulgare[J]. J Biol Chem, 1961, 236(10): 2692-2698.
[9] 王敬文, 薛應龍. 植物苯丙氨酸解氨酶的研究[J]. 植物生理學報, 1981, 7(4): 373-380.
[10] GOFFNER D, JOFFROY I, GRIMA P J, et al. Purification and characterization of isoforms of cinnamyl alcohol denydrogenase from Eucalyptus xylem[J]. Planta. 1992, 188(1): 48-53.
[11] HAMMERSCHMIDT R, KUC J. Lignification as a mechanism for induced systemic resistance in cucumber[J]. Physiological and Plant Pathology, 1982, 20(1): 61-71.
[12] KAR M, SMITH E H. Calalase. Peroxidase and polyphenol oxidase actibitiesduring rice leaf senescence[J]. Plant Physioligy, 1976, 57: 315-319.
[13] SRIVASTAVA M K, DWIVED U N. Delayed ripening of banana fruit by salicylic acid[J]. Plant Sci, 2000, 158(1/2): 87-96.
[14] 芮懷瑾, 汪開拓. 熱處理對冷藏枇杷木質化及相關酶活性的影響[J].農業工程學報, 2009, 25(7): 294-298.
[15] 徐俐, 陸加貴, 劉萬軍. 不同保鮮劑對竹筍纖維化及保鮮效果的影響[J]. 貴州大學學報: 農業與生物科學版, 2002, 20(2): 110-114.
[16] 劉尊英, 姜微波, 趙玉梅, 等. 溫度、赤霉素和乙烯處理對采后豌豆苗纖維合成及品質變化的影響[J]. 中國農業大學學報, 2003, 8(1): 75-77.
[17] 吳振先, 許曉春, 陳維信, 等. 荔枝果皮褐變機理研究進展[J]. 保鮮與加工, 2004, 4(1): 8-10.
[18] 鄭永華, 李三玉, 席玙芳. 枇杷冷藏過程中果肉木質化與細胞壁物質變化的關系[J]. 植物生理學報, 2000, 26(4): 306-310.
[19] 吳錦程, 唐朝暉, 陳群, 等. 不同貯藏溫度對枇杷果肉木質化及相關酶活性的影響[J]. 武漢植物學研究, 2006, 24(3): 235-236.
[20] 許曉春, 吳振先, 陳維信, 等. 荔枝果皮褐變機理研究進展[J]. 保鮮與加工, 2004, 4(1): 8-10.
[21] 席嶼芳, 羅自生, 程度, 等. 竹筍采后木質化與多酚氧化酶、過氧化酶和苯丙氨酸解氨酶活性的關系[J]. 植物生理學通訊, 2001, 37(4): 294-295.
Effect of Storage Temperature on Enzyme Activities Associated with Peel Lignification and Pulp Decay of Mangosteen
YANG min1,YU Li-mei1,*,LIU Zhao-xia2,CHEN Hai-guang1
(1. College of Light Industry and Food Technology, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China;2. Guangdong Inspection and Quarantine Technology Center, Guangzhou 510623, China)
Mangosteen harvested from Hainan province was stored at 30 or 4 ℃ to investigate the effect of storage temperature on enzyme activities associated with peel lignification and pulp decay of mangosteen during postharvest storage. Results showed the degree of peel lignifications varied depending on the storage temperature. Chilling injury occurred at the early period of cold storage, leading to increased enzyme activities and lignin content, but lignin content was increased by 14.3% at the later stage of storage at 30 ℃ than storage at 4 ℃. In addition, the activities of PAL, POD, PPO and CAD in mangosteen stored at 30 ℃ were significantly higher than those observed at 4 ℃. The peak PAL activity at 30 ℃ and 4 ℃was 170.40 U/(g?h) and 180.12 U/(g?h), respectively. A continuous decrease in PAL activity was observed at the later stage of storage, regardless of storage temperature. The decay index of mangosteen after 15 days of storage at 30 ℃ was 78.89% compared to 32.22% at 4 ℃. This result may be related to inhibited enzyme activities involved in peel lignifications at low temperature, reduced accumulation of cellular metabolites and low respiration intensity, therefore inhibiting pulp decay.
mangosteen fruit;lignification;enzymes activity;storage temperature
TS255.36
A
1002-6630(2013)18-0307-05
10.7506/spkx1002-6630-201318063
2012-07-03
廣東省自然科學基金項目(10451022501005679)
楊敏(1967—),女,副教授,碩士,研究方向為食品貯藏與加工。E-mail:yangmgz-001@163.com
*通信作者,于立梅(1973—),女,副教授,博士,研究方向為食品貯藏與加工。E-mail:biyingwang2003@163.com
