青蒿琥酯對肺泡上皮細胞TGF-β1/MAPK通路的影響*
2013-03-04 00:39:18王昌明黎紅秀
重慶醫學 2013年14期
關鍵詞:肺纖維化
陳 娟,王昌明,周 燕,黎紅秀
(桂林醫學院附屬醫院呼吸內科,廣西桂林541001)
特發性肺纖維化(idiopathic pulmonary fibrosis,IPF)是一組多種病因引起的以氣道受損、炎癥因子大量分泌、間充質細胞增生和細胞外基質異常沉積為特點的漸進性疾病,患者確診后平均生存期僅3~5年[1]。IPF發病機制不明,近年來國內外研究表明肺泡上皮細胞-間質細胞轉分化(epithelial-mesenchymal transition,EMT)是肺纖維的重要機制之一,細胞因子轉化生長因子-β1(transforming growth factor beta1,TGF-β1)可通過活化p38絲裂原活化蛋白激酶類(p38mitogen-activated protein kinases,p38MAPK)誘導 EMT 過程[2]。目前,對IPF尚缺乏有效的治療方法,臨床上常用糖皮質激素和免疫抑制劑,但效果并不滿意。本課題組前期研究表明,青蒿素衍生物青蒿琥酯除抗瘧疾外,還具有抗纖維化作用[3-4]。本研究用青蒿琥酯為工具藥,研究在其干預下對TGF-β1誘導的EMT過程中的影響及其可能機制。
1 材料與方法
1.1 材料 永生化大鼠肺泡Ⅱ型上皮細胞(rat lung epithelial-t-antigen negative,RLE-6TN)株購自中南大學湘雅實驗中心細胞庫。TGF-β1購自Peprotech公司(美國);四甲基偶氮唑鹽(MTT)粉劑購自sigma公司(美國);Total RNA提取試劑、逆轉錄試劑盒購自TaKaRa寶生物工程大連有限公司;引物由Invitrogen公司設計合成;組織細胞裂解液(tissue and cell lysis solution,WIP)、BCA蛋白濃度測定試劑盒購自北京博奧森生物技術有限公司;p38MAPK兔抗人一抗購自Santa cruz(美國);α-平滑肌肌動蛋白(α-smooth muscle actin,α-SMA)兔抗人單克隆抗體購自ABBIOTEC公司(美國);V-波形蛋白(vimentin,Vim)兔抗人一抗購自Bioworld Technology;辣根酶標記山羊抗兔IgG購自中杉金橋;胎牛血清購自GIBCO公司;1640培養基購自HyClone公司。
1.2 方法
1.2.1 細胞培養 RLE-6TN細胞在37℃、5%CO2、飽和濕度的條件下用含10%胎牛血清的1640培養基培養。穩定傳代后用含1%胎牛血清的培養液培養細胞同步化24h,根據相關文獻及預實驗結果,誘導劑TGF-β1按說明書配置好母液后稀釋于含1%胎牛血清的培養液中,終濃度為3ng/mL,青蒿琥酯用NaHCO3溶解后再調整到相應終濃度作用于細胞。
1.2.2 細胞分組 MTT粉劑用PBS配成5mg/mL的存儲液,過濾除菌,4℃保存。將細胞分為6組,1個對照組、5個實驗組(其中1個誘導組,4個藥物干預組)。計數細胞,3 000個/孔細胞種植96孔板,設5個復孔,在1%FBS(對照組)、TGF-β13ng/mL(TGF-β1組)、TGF-β13ng/mL+青蒿琥酯1 mg/L(TGF-β1聯合1組)、TGF-β13ng/mL+青蒿琥酯2mg/L(TGF-β1聯合2組)、TGF-β13ng/mL+ 青蒿琥酯 4mg/L(TGF-β1聯合3組)、TGF-β13ng/mL+青蒿琥酯8mg/L(TGF-β1聯合4組)作用細胞24h后加入MTT溶液至終濃度0.5mg/mL,繼續培養4h,吸干凈 MTT溶液,加入二甲基亞砜(dimethyl sulfoxide,DMSO)150μL,避光平搖10min,用酶標儀測定490nm處的吸光度值(A)。根據抑制率=﹝(對照組OD490值-實驗組OD490值)/對照組OD490值﹞×100%計算細胞抑制率。用寇式改良法計算IC50,根據IC50選擇藥物干預濃度進行以下實驗。
1.2.3 倒置顯微鏡觀察 當細胞生長融合至60%~80%時,棄培養液,換含1%胎牛血清的1640培養液靜止24h,換新鮮靜止液,于對照組、TGF-β1組及 TGF-β1聯合1、2、3、4組培養液中分別加入1%FBS、TGF-β13ng/mL及TGF-β13ng/mL+青蒿琥酯1、2、4、8mg/L作用細胞24h后,使用倒置相差顯微鏡觀察細胞形態學的改變并拍照。
1.2.4 Western blot檢測上皮及間質細胞標志物的表達 各組細胞生長、培養同1.2.3,培養細胞24h后用WIP裂解液(1 mL)+蛋白酶抑制劑(10μL)+磷酸酶抑制劑(10μL)混合,每皿(d=10cm)加入 WIP裂解液混合液200μL,冰上裂解45 min,用細胞刮刮下后離心取上清液,BCA蛋白測定試劑盒做標準曲線測定蛋白濃度后,取30μg樣品蛋白進行10%的十二烷基磺酸鈉-聚丙烯酰胺(SDS-PAGE)凝膠變性電泳,然后轉膜至聚偏二氟乙烯(PVDF)膜,加入一抗,37℃孵育2h,Tris-Buffered-Saline with Tween-20(TBST)洗滌后加辣根酶標記二抗,37℃孵育2h,TBST洗滌后加適量ECL發光液(ECL luminous liquid)暗室發光,曝光后在凝膠成像系統下掃描并獲取密度值。
1.3 統計學處理 采用SPSS17.0統計軟件進行分析,各實驗均重復3次。實驗數據以±s表示,差異顯著性分析采用One-way ANOVA分析,組間采用方差分析比較,以P<0.05為差異有統計學意義。
2 結 果
2.1 青蒿琥酯對TGF-β1誘導的RLE-6TN細胞增殖的影響TGF-β1作用于RLE-6TN 24h后,細胞較對照組增殖明顯(P<0.05);青蒿琥酯不同濃度作用于 TGF-β1誘導的 RLE-6TN 24h后,細胞增殖受到抑制(表1)。根據抑制率用寇式改良法計算IC50,24hIC50=8.86mg/L。
2.2 青蒿琥酯對TGF-β1誘導的RLE-6TN細胞形態的影響與對照組比較,加入TGF-β1后,RLE-6TN由鋪路石狀上皮形態變成梭形、紡錘形,而且其細胞間隙略變大,細胞和細胞之間的連接變得松散。在TGF-β1誘導后再加入青蒿琥酯,即TGF-β1聯合1、2、3、4組,與 TGF-β1組比較,細胞的梭形結構減少,部分恢復鋪路石狀上皮形態,但細胞之間連接仍松散,見圖1。
2.3 青蒿琥酯對TGF-β1誘導的RLE-6TN細胞p38MAPK及間質細胞標志性蛋白表達的影響 細胞培養24h后,TGF-β1誘導RLE-6TN表達p38MAPK、α-SMA、Vim均較對照組明顯增加(P<0.05),青蒿琥酯作用于 TGF-β1誘導的 RLE-6TN后,p38MAPK、α-SMA、Vim表達均較 TGF-β1組明顯減少(P<0.01),且各組間減少呈劑量依賴性(P<0.05)。內參為甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH),各組蛋白表達的電泳圖,見圖2。

圖1 倒置顯微鏡下各組細胞形態(×100)
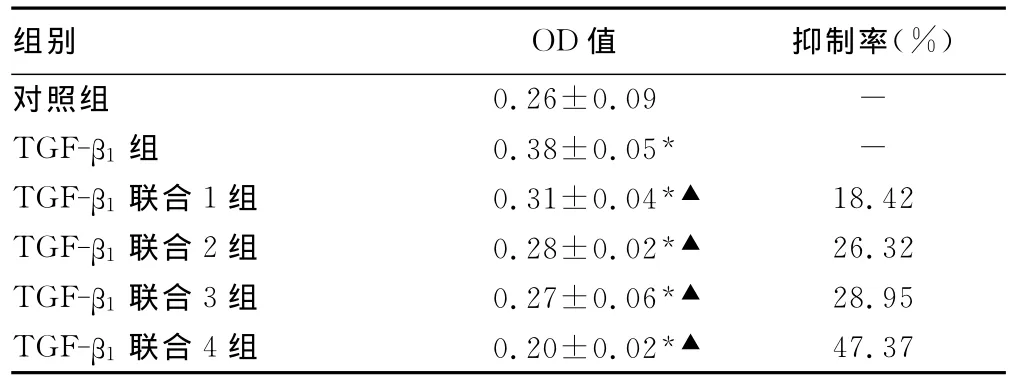
表1 各組細胞24h后增殖抑制率比較
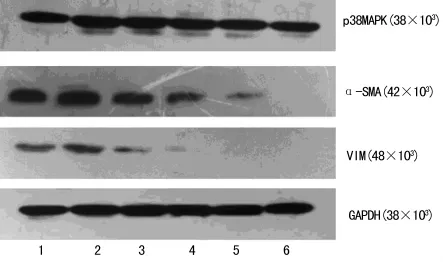
圖2 各組蛋白表達的電泳圖
3 討 論
肺間質纖維化(Pulmonary fibrosis,PF)是一組多種病因引起的以氣道、肺泡結構排列紊亂,膠原大量沉積為特點的漸進性疾病。臨床表現為刺激性干咳、無痰、進行性的呼吸困難等,最終可以引起呼吸功能衰竭而死亡。目前肺纖維化的病因及其發病機制尚未完全闡明,但都有其共同規律,肺部受感染或損傷后,損傷的肺上皮細胞和血管內皮細胞產生大量的細胞因子,刺激炎癥細胞的聚集、活化,分泌膠原等細胞外基質,沉積在肺間質,導致肺纖維化[5]。
TGF-β1是一種多效性的細胞因子,是目前肺纖維化的研究熱點和重要藥物作用靶點。TGF-β1可由淋巴細胞、單核細胞、上皮細胞和成纖維細胞等多種細胞產生,通過自分泌和旁分泌的方式調節細胞的增殖、分化、遷移、黏附、調節細胞外基質的代謝,參與肺胚胎發育、組織損傷和修復[6]。
Furukawa等[7]研究表明,TGF-β1誘導MAPK磷酸化活性變化,通過MAPK信號轉導途徑調節人肺上皮細胞向肌纖維母細胞轉化,產生大量的細胞外基質,導致肺間質纖維化。肺泡上皮細胞(alveolar epithelial cell,AEC)包括Ⅰ和Ⅱ型,肺泡Ⅱ型上皮細胞(typeⅡalveolar epithelial cell or typeⅡpneumocyte,ATⅡ)是一種多功能細胞,對維持肺泡結構和功能有重要意義。它是Ⅰ、Ⅱ型上皮細胞的祖細胞[8-9],在正常細胞更新和損傷修復過程中,ATⅡ可以分化為Ⅰ型細胞,也可通過有絲分裂補充自身的數量。此外,ATⅡ具有合成和分泌肺表面活性物質,維持肺泡內外液體平衡,免疫調節等作用[10]。
絲氨酸/蘇氨酸蛋白激酶(MAPKs)信號通路是真核細胞轉導細胞外信號到細胞內引起細胞反應的四大信號系統之一,其參與了細胞生長、發育及細胞間功能同步等多種生理功能,目前已知MAPKs由4個主要成員構成,即細胞外信號調節激酶 (ERK1/ERK2)、應激活化蛋白激酶 (SAOK/JNK)、p38MAPK和ERK5。其中p38MAPK與肺纖維化過程密切相關[11]。TGF-β1誘導活化TGF-β1受體復合物,級聯活 化p38MAPK,p38MAPK把信號傳遞到細胞核內,活化EMT相關轉錄因子Snail家族蛋白中的Snail1和Slug表達,從而引起細胞形態及分泌功能的改變。Snail家族蛋白通過鋅指結構域及N-末端結構域起著轉錄抑制子的作用,Snail1在EMT過程中的作用早已有報道[12]。
張新志等[13]用抗纖靈沖劑作用于SD大鼠后,發現抗纖靈沖劑可通過調節TGF-β1/p38MAPK信號轉導通路,發揮抗腎小管間質纖維化效應。Matsuoka等[14]研究也證實,用p38MAPK拮抗劑干預由博萊霉素誘導的肺纖維化動物模型后纖維化程度明顯減輕,以上資料均顯示p38MAPK的活化與纖維化的發生、發展密切相關。
Willis等[15]研究表明Vim是一種中間絲蛋白,是間質細胞的典型標記物,α-SMA是一種肌動蛋白,幾乎所有的真核細胞都表達肌動蛋白,常用于標記間質細胞。TGF-β1誘導的EMT早期變化就是細胞質內細胞骨架進行重排,表達新的表型蛋白如α-SMA,細胞質內肌絲也從上皮型的角蛋白(cytokeratin,CK)轉變為間質細胞的Vim。細胞黏附連接主要成分E-鈣黏蛋白的下調或丟失,緊密連接成分的下調,以及間充質細胞特異性的蛋白標記,如纖維連接蛋白、Vim和N-鈣黏蛋白的表達上調已成為人們判斷EMT發生的重要的分子指標[16]。
青蒿琥酯是抗瘧疾的有效單體,近年來研究發現青蒿琥酯除了有很好的抗瘧作用外,還具有抗炎、抗腫瘤、抗纖維化等作用[17]。本課題前期研究表明青蒿琥酯可以抑制人胚肺成纖維細胞增殖、促進其凋亡[3-4],且呈劑量依賴性。以上的研究結果為青蒿素防治肺纖維化提供了理論依據。但是目前國內外關于青蒿琥酯防治肺纖維化的研究報道較少,主要是關于其對成纖維細胞增殖的抑制作用,而對青蒿琥酯調控TGF-β1誘導的EMT的作用機制研究少見報道。
本研究穩定傳代ATⅡRLE-6TN后,予TGF-β13ng/mL誘導ATⅡ轉分化為間質細胞,結果表明,TGF-β1作用于RLE-6TN 24h后,p38MAPK、α-SMA、Vim蛋白均表達增加(P<0.05)。可認為TGF-β1成功誘導RLE-6TN轉分化為間質細胞,其可能機制為活化p38MAPK蛋白。給予不同劑量的青蒿琥酯作用于TGF-β1誘導后的RLE-6TN 24h后,信號蛋白p38MAPK及間質細胞標志物α-SMA、Vim蛋白均較TGF-β1組表達明顯減少(P<0.01),且呈濃度依賴性。可認為青蒿琥酯對TGF-β1誘導的轉分化過程有抑制作用,且隨著青蒿琥酯濃度的增加,這種抑制作用逐漸增強。
綜上所述,TGF-β1能誘導EMT過程。青蒿琥酯具有抗纖維化作用,其可能機制為抑制p38MAPK的活性從而抑制EMT過程。
[1]陳恒屹,崔社懷.特發性肺間質纖維化發病機制及治療進展[J].中華肺部疾病雜志:電子版,2009,2(1):58-62.
[2]李飛鳳,周建華,胡永斌,等.p38MAPK信號通路調控二氧化硅誘導的人支氣管上皮細胞上皮-間質轉型[J].工業衛生與職業病,2011,37(2):69-73.
[3]王昌明,張孝飛,譚寧,等.青蒿琥酯對人胚肺成纖維細胞系HFL-I細胞增殖,凋亡的影響及機制[J].山東醫藥,2009,50(3):33-35.
[4]王昌明,張孝飛,黃嵐珍,等.青蒿琥酯誘導人胚肺成纖維細胞凋亡的分子機制研究[J].時珍國醫國藥,2010,21(11):2837-2838.
[5]孔琪,秦川.肺纖維化的發病機制及關鍵靶點[J].國外醫學:呼吸系統分冊,2005,25(5):331-333,336.
[6]石永利,孟凡青.TGF-β1誘導的上皮-間葉轉變與肺纖維化[J].臨床與實驗病理學雜志,2011,27(5):527-529.
[7]Furukawa F,Matsuzaki K,Mori S,et al.p38MAPK mediates fibrogenic signal through Smad3phosphorylation in rat myofibroblasts[J].Hepatology,2003,38(4):879-889.
[8]Emura M.Stem cells of the respiratory epithelium and their in vitro cultivation[J].In Vitro Cell Dev Biol,1997,33(1):3-14.
[9]Uhal BD.Cell cycle kinetics in the alveolar epithelium[J].Am J Physiol,1997,272(6Pt 1):L1031-1045.
[10]Pison U,Wright JR,Hawgood S.Specific binding of surfactant apoprotein SP-A to rat alveolar macrophages[J].Am J Physiol,1992,262(4Pt 1):L412-417.
[11]黃政坤,包建東,戴軍,等.p38MAPK抑制劑對急性肺損傷大鼠肺組織核因子-κB表達的影響[J].臨床麻醉學雜志,2007,23(4):323-325.
[12]Nieto MA.The snail superfamily of zinc-finger transcription factors[J].Nat Rev Mol Cell Biol,2002,3(3):155-166.
[13]張新志,黃迪,吳鋒,等.TGF-β1/p38MAPK 通路對腎間質纖維化影響及抗纖靈沖劑干預機制的實驗研究[J].中華中醫藥雜志,2011,26(2):245-248.
[14]Matsuoka H,Arai T,Mori M,et al.A p38MAPK inhibitor,FR-167653,ameliorates murine bleomycin-induced pulmonary fibrosis[J].Am J Physiol Lung Cell Mol Physiol,2002,283(1):L103-112.
[15]Willis BC,dubois RM,Borok Z.Epithelial origin of myofibroblasts during fibrosis in the lung[J].Proc Am Thorac Soc,2006,3(4):377-382.
[16]宋建國,王冬梅.上皮細胞向間質細胞轉變的調控及其與細胞凋亡的關系[J].生命科學,2010,22(11):1147-1153.
[17]楊耀芳.青蒿素及衍生物的藥理作用和臨床應用[J].中國臨床藥學雜志,2003,12(4):253-254.
猜你喜歡
中老年保健(2022年2期)2022-11-25 23:46:31
昆明醫科大學學報(2022年4期)2022-05-23 13:04:50
昆明醫科大學學報(2021年4期)2021-07-23 01:21:34
國際呼吸雜志(2019年21期)2019-11-25 09:52:20
國際呼吸雜志(2019年20期)2019-11-23 08:46:14
國際呼吸雜志(2019年8期)2019-04-29 09:15:22
國際呼吸雜志(2019年3期)2019-03-01 05:39:12
中國繼續醫學教育(2015年3期)2016-01-06 01:36:31
吉林大學學報(醫學版)(2015年3期)2015-12-17 07:47:38
天津醫科大學學報(2015年3期)2015-06-05 12:21:49
