低植酸玉米育種的研究概況
2012-12-24 12:51:00徐秀紅郭國錦呂桂華盧華兵
浙江農業科學 2012年10期
徐秀紅,郭國錦,呂桂華,盧華兵
(浙江省東陽玉米研究所,浙江 東陽 322100)
植酸,又名肌醇六磷酸,是成熟作物種子中含磷最豐富的化合物,占種子干重的1%以上,總磷含量的65% ~85%[1]。是種子萌發和幼苗生長所需肌醇和磷酸等營養成分的貯藏庫,是玉米籽粒中廣泛存在的一種有機酸 (肌醇六磷酸)。植酸可與Zn2+、Fe3+、Ca2+等金屬陽離子螯合形成植酸鹽,難以被人和非反芻動物 (豬、雞、魚等)消化利用,降低了磷元素以及與之結合的微量金屬元素、蛋白質和淀粉的生物有效性,尤其在發展中國家植酸是造成微量營養元素缺乏的主要原因,被認為是一種抗營養因子[2]。鰲合形成的植酸鹽因為不能被吸收利用,隨著糞便排出體外,進入水體后造成水體富營養化,繼而引起水體環境污染。另外,土壤中因為缺乏分解微生物,難以分解這些植酸鹽,即使畜禽糞便作為有機肥還田,也難以被農作物吸收利用;大量磷肥的使用,也造成了不可再生磷資源的浪費,人類將會面臨磷礦短缺的局面。
培育低植酸作物,能夠降低植酸抗營養性,提高微量元素營養成分的生物有效性,可減少磷對環境的污染和動物飼料中磷元素的添加,這些已成為育種學家、營養學家和環境學家關注的焦點之一[3]。因此,作物低植酸種質的創新和品種選育近年來已成為育種家們研究的熱點,并先后在玉米、大麥、水稻、小麥、大豆等作物中創造出低植酸突變體。本文綜述了迄今為止玉米低植酸突變種質創新、突變體植酸含量變化、農藝性狀及種子特性、突變的遺傳學研究方面的研究進展,并對低植酸玉米的研究前景進行展望。
1 低植酸玉米突變體的選育
低植酸作物的選育開始于1992年,主要是應用化學誘變劑 (疊氮化鈉,EMS)或γ-射線誘變。低植酸玉米最早是Raboy等[4]在1996年采用EMS化學誘變劑對玉米花粉進行誘變,育成了2份玉米低植酸突變體lpa1-1和lpa2-1,植酸含量分別下降66%和30%。Pilu等[5]通過 EMS誘導玉米花粉,育成了一份低植酸玉米突變體lpa241,其植酸下降90%。隨后,Shi等[6-8]利用轉座子插入和基因沉默的方式育成了3份玉米低植酸突變體lpa2、lpa3和lpa1,植酸分別下降30%、50%和93%。在國內,低植酸玉米研究稍晚,2006年浙江大學核農所王雪艷等[9]開始了玉米低植酸突變體的初步篩選,獲得了6個低植酸突變體,植酸下降幅度在43.94%~79.09%。中國農科院于2007年9月10日宣布了轉植酸酶基因玉米的問世,并在2009年獲得了農業部頒發的轉基因生物安全證書,成為我國首例獲得安全證書的糧食作物。之后,劉欣芳等[10-11]也開展對轉植酸酶基因玉米的獲得及其后代的初步鑒定。
目前,選育低植酸玉米的主要方法是:用EMS處理玉米花粉或者γ-射線處理干種子,M1代自交獲得M2代種子,混收;M2按單株種植,套袋授粉自交,按株收獲種子,形成株系。每株系取成熟的8粒M3種子,按Chen等測定無機磷 (Pi)含量的方法進行無機磷檢測,如果檢測到1粒以上呈高無機磷 (HIP)種子的單株,此單株將被認為是可能的突變株,8粒全部呈HIP的單株視為可能的純合低植酸突變體,繼續種植檢測,若還為純合,則可確認為低植酸突變體。研究表明,低植酸突變體的植酸含量下降都伴隨著無機磷含量的成倍增加[8],且正常玉米中無機磷的含量很低,在0.3~0.5 mg·g-1,因此,篩選低植酸突變體可以通過種子的無機磷檢測來間接選擇[12]。
2 低植酸玉米突變體的植酸含量及其他成分含量的變化
玉米種子中植酸含量在8.3~22.2 mg·g-1,占種子干重的0.69%~1.14%,占種子全磷量的75%~80%,植酸主要的沉積部位是盾片層(胚),占90%,只有10%在糊粉層[13-14]。玉米種子中植酸的含量一般受品種、種子成熟度、栽培地點及土壤氣候條件等影響,主要由品種的遺傳特性和環境條件決定的[12]。
有關研究表明,玉米低植酸突變體植酸含量的降幅達30% ~93%。最早Raboy等發現的lpa1-1和lpa2-1 2個突變體植酸的降幅分別為66%和50%,lpa1-1在植酸下降的同時,其無機磷含量同等比例升高,但肌醇和低價肌醇磷酸鹽中間物沒有積累;lpa2-1植酸含量下降伴隨無機磷的升高,五價或更低價的肌醇磷酸鹽含量上升,野生型中卻未積累相關的成分。與野生型相比,以上2種低植酸突變體的總磷含量都保持不變[2,4]。Pilu 等[5]發現的lpa241突變體,其與lpa1-1等位,但植酸的降幅為90%。Shi等[7]發現一個低植酸突變體,與lpa1和lpa2不等位,命名為lpa3,其植酸的降幅是50%,其植酸含量下降的同時,無機磷和肌醇含量顯著增加,未積累低價肌醇磷酸鹽中間物,總磷含量也保持不變。Shi等發現的另外2個突變體,按照其他成分含量的變化,分別歸類于lpa2和lpa1。
迄今為止,發現的玉米低植酸突變體多數總磷含量基本不變,在正常的環境條件下均能自然發芽,是非致死型突變體,但Larson等[5,15]在低植酸玉米研究中發現了一類突變體,其在正常的栽培環境條件下純合的突變體種子不能發芽,低植酸性狀只能通過雜合的株加以保留;Pilu等[5]在玉米低植酸的研究中共篩選到29份低植酸突變體,發現有28份是致死型的,其中18份可能由于胚特異性致死,另外的10份可能由于胚乳引起。
3 玉米低植酸突變體的農藝性狀及種子特性變化
植酸在種子的萌發和植物的生長發育過程中起著重要作用,因此,研究所發現的絕大多數玉米低植酸突變體隨著植酸含量的下降,一些農藝性狀和種子的品質特性也出現了變化,與對照野生型相比,突變體的發芽率和成苗率降低,種子的干重下降,繼而產量下降,抗逆性變差,生育期延遲等。
Raboy等[4]在研究低植酸玉米時發現,與對照相比,lpa1-1雜合型的發芽、莖葉長度、種子成熟時的含水量以及花期等農藝性狀都沒有顯著的差異,但其產量比對照減產8% ~23%,lpa2-1減產4%~16%。大部分低植酸突變體純合狀態下種子具有活力,包括一些植酸含量幾乎接近零的突變體,但其產量性狀都會變差,植酸含量下降小于75%時,與野生型相比,產量損失一般在5%~15%;當植酸含量下降幅度巨大,達到90%~98%時,突變體的生長發育就會受到嚴重的影響,甚至會死亡,這些都將導致產量嚴重下降[2]。Pilu等[5]在研究低植酸玉米突變體lpa241時發現,與對照相比,突變體的發芽率下降了30%,但其生長發育卻沒有受到顯著影響。
此外,還有一些研究表明,轉基因技術育成若干玉米低植酸突變體,其在農藝性狀和種子的品質特性上與親本沒有顯著的區別。運用反向遺傳學方法,利用Mu轉座子插入和球蛋白的胚特異性啟動子構建的基因沉默載體,Shi等[12]獲得玉米低植酸突變體lpa3、lpa2和lpa1,突變并沒有影響一般田間條件下的種子發芽率,種子干重也沒有顯著下降。中國農科院范云六等[11]自主研發的轉植酸酶基因玉米,是從曲霉屬的一種真菌中分離出能產生植酸酶的基因,并把它插入玉米基因組中而獲得的,這種轉植酸酶基因玉米在種子發芽率、生長速度和產量上都沒有受到影響。
4 玉米低植酸突變的遺傳學研究
4.1 遺傳模式
研究證實,目前所獲得的玉米低植酸突變均為隱性突變,共出現3類突變體,分別受1對非等位隱性基因控制,如純合的lpa1/lpa1和lpa2/lpa2雜交,后代植酸水平正常,說明這2個突變位點互不等位;而Shi等獲得的低植酸突變體與lpa1和lpa2雜交后,發現其與這2個突變位點互不等位,定為lpa3。這些突變體的雜合型后代分離比例基本符合,野生型∶雜合型∶低植酸基因型1∶2∶1,野生型∶低植酸型表現型為3∶1的遺傳模式。
研究中還發現,有些分離后代中純合單株的比例低于理論值,即在后代的分離群體中純合的低植酸植株較少,這可能與其發芽力下降有關系。
4.2 基因定位及克隆
在玉米中,已發現3類純合低植酸突變體lpa1、lpa2和lpa3,且都位于1S染色體上,其中lpa1-1突變位點與RFLP分子標記umc157相連鎖,遺傳距離為7.7 cM[4];lpa2-1突變位點則與RFLP分子標記umc167相連鎖,遺傳距離為10 cM[14]。另一個玉米低植酸突變體 lpa241,與lpa1等位,與RFLP分子標記umc1222相距大約9.2 cM[5]。
在種子的發育過程中,植酸合成途徑中任何一步的酶發生突變或植酸的運輸途徑受阻,就會造成植酸的合成或積累受抑制,引起種子植酸含量的下降,產生低植酸突變體[15]。一個突變體植酸含量的下降,可能是植酸合成的2個底物——肌醇和無機磷的合成或供應受阻導致,也可能是2個底物轉化成磷酸鹽的過程受阻,還有可能是植酸的運輸或者其他基因調控引起。因此,了解植酸的代謝途徑(圖1),對研究低植酸突變體非常重要,圖1中與植酸代謝途徑相關的基因已經相繼被克隆出來[16]。
迄今為止,作物中發現的低植酸突變基因有很多,大豆中發現的突變基因主要是肌醇磷酸合成酶(MIPS)基因[17-18]和抗藥相關蛋白 (MRP)基因[19],擬南芥中發現的主要是肌醇磷酸激酶(AtIPK1和AtIPK2β)基因[20],水稻中發現的有肌醇激酶 (MIK)基因[21]和 MRP 基因[22],大麥中發現的有硫轉運蛋白 (HvST)基因[23]。在玉米中目前發現的3個低植酸突變的基因已經被克隆出來,玉米 lpa1突變是由于1個 MRP基因(ZmMRP4)突變導致,它編碼一個MRP-ABC(ATP結合盒)轉運蛋白,構建基因沉默載體使其沉默后,產生了植酸含量下降無機磷升高的玉米低植酸突變體lpa1;玉米lpa2突變是由于一個肌醇磷酸激酶 (ZmIpk)基因序列的重排導致,在ZmIpk基因的編碼序列核苷酸158位置上,DNA序列發生由C到T突變,在開放閱讀框的N-末端產生一個終止密碼子,僅有34個氨基酸,蛋白翻譯終止,促使肌醇磷酸激酶活性下降,多磷酸肌醇的合成減少,突變體的植酸含量下降約30%,無機磷含量增加約3倍,肌醇和肌醇磷酸鹽IP3、IP4和IP5得到積累;玉米lpa3突變則是由于MIK基因發生突變引起,它使催化肌醇和磷酸生成肌醇-3-磷酸的步驟受抑,最終導致植酸含量的下降[12]。
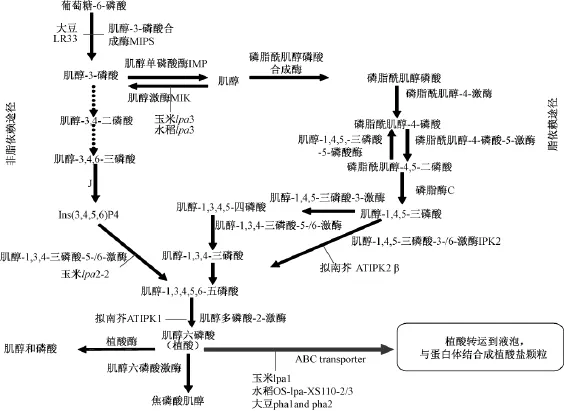
圖1 植物體內植酸的代謝途徑及引起植酸含量下降的已報道突變基因
5 展望
玉米是糧食、飼料和經濟兼用型作物,玉米總產量的75%用于飼料加工,是飼料原料總量的60%。低植酸玉米直接作為飼料,可提高動物對微量金屬元素的吸收利用,提高其生物有效性,有效改善微量金屬元素缺乏癥,同時可減少其糞便中的磷含量,提高磷的利用率,減少環境中的磷污染,達到綠色環保的目的;或以低植酸玉米為原料生產飼料,在生產過程中可不加植酸酶或降低其使用量,從而降低生產成本,增加畜禽對飼料中的鈣、鐵、鋅等金屬微量營養元素的吸收和利用,提高其生物有效性。因此,低植酸玉米的研究應用前景廣闊。
研究表明,低植酸突變會使農藝性狀變差,種子的品質和特性一定程度上變劣,這說明植酸含量的下降與產量的下降呈正相關[24]。產量性狀的下降表明,直接利用低植酸突變體育種存在一定難度,但對產量影響較小的突變也可用于育種,選擇不同的低植酸突變基因可減少農藝性狀變差和種子品質變劣[25]。另外,還可以通過多次回交、雜交等改良低植酸品種的產量性狀。此外,研究結果表明,大多數轉基因玉米其農藝性狀和種子特性沒有受到影響,這也給低植酸玉米育種帶來新手段。隨著低植酸玉米育種技術的不斷成熟,具有營養和環境雙重功能的低植酸玉米雜交種的選育將為改善動物和人類的營養、減少環境污染等作出巨大貢獻。
[1] Raboy V.Accumulation and storage of phosphate and minerals[G] //Larkins B A,Vasil I K.Cellular and Molecular Biology of Plant Seed Development. Dordrecht,the Netherlands:Kluwer Academic Publishers,1997:441-477.
[2] Raboy V.Seeds for a better future:“low phytate”grains help to overcome malnutrition and reduce pollution [J].Trends in Plant Sci,2001,6(10):458-462.
[3] Lott J N A,Ockenden I,Raboy V,et al.Phytic acid and phosphorus in crop seeds and fruits:a global estimate [J].Seed Sci Res,2000,10:11-33.
[4] Raboy V,Gerbasi P F,Young K A,et al.Origin and seed phenotype of maize low phytic acid 1-1 and low phtic acid 2-1 [J].Plant Physiol,2000,124:355-368.
[5] Pilu R,Panzeri D,Gavazzi G,et al.Phenotypic,genetic and molecular characterization of a maize low phytic acid mutant(lpa 241)[J].Theor Appl Genet,2003,107:980-987.
[6] Shi J R,Wang H Y,Wu Y S,et al.The maize low-phytic acid mutant lpa2 is caused by mutation in an inositol phosphate kinase gene [J].Plant Physiol,2003,131:507-515.
[7] Shi J R,Wang H Y,Hazebroek J,et al.The maize lowphytic acid 3 encodes a myo-insitol kinase that plays a role in phytic acid biosynthesis in developing seeds [J].T Plant J,2005,42:708-719.
[8] Shi J R,Wang H Y,Schellin K,et al.Embryo-specific silencing of a transporter reduces phytic acid content of maize and soybean seeds [J]. NatBiotechnol, 2007, 25:930-937.
[9] 王雪艷,王忠華,梅淑芳,等.高無機磷低植酸玉米突變體篩選初報[J].核農學報,2006,20(1):15-18.
[10] 劉欣芳,高曉蓉,蘇喬,等.轉植酸酶基因玉米的獲得及其后代的初步鑒定 [J].玉米科學,2008,16(1):15-19.
[11] 張琪,陳茹梅,楊文竹,等.組成型表達轉植酸酶基因(phyA2)玉米的獲得[J].農業生物技術學報,2010,18(4):623-639.
[12] 徐秀紅.一個植酸代謝相關水稻基因的定位、克隆與特性研究[D].杭州:浙江大學,2009.
[13] Raboy V.Low-phytic-acid grains [J].Food Nutr Bull,2000,21:423-427.
[14] Larson S R,Raboy V.Linkage mapping of maize and barley myo-inositol1 - phosphate synthase DNA sequences:correspondence with a low phytic acid mutation [J].Theor Appl Genet,1999,99:27-36.
[15] RaboyV.The ABCsoflow-phytate crops [J].Nat Biotechnol,2007,25:874-5.
[16] Raboy V.Forward Genetics Studies of Seed Phytic Acid [J].Israel Journal of Plant Sciences,2008,55:171-181.
[17] Hitz W D,Carlson T J,Kerr P S,et al.Biochemical and molecular characterization of a mutation that confers a decreased raffinosaccharide and phytic acid phenotype on soybean seeds[J].Plant Physiol,2002,128:650 –660.
[18] Yuan F J,Zhao H J,Ren X L,et al.Generation and characterization of two novel low phytate mutations in soybean(Glycine max L.Merr) [J].Theor Appl Genet,2007,115:945-957.
[19] Gillman J D,Pantalone V R,Bilyeu K.The low phytic acid phenotype in soybean line CX1834 is due to mutations in two homologs of the maize low phytic acid gene [J].Plant Genome,2009,2:179-190.
[20] Stevenson-Paulik J, BastidasR J, Chiou S T, etal.Generation ofphytate-free seeds in Arabidopsis through disruption of inositol polyphosphate kinases[J].Proc Natl Acad Sci USA,2005,102:12612–12617.
[21] Kim S I,Andaya C B,Newman J W,et al.Isolation and characterization of a low phytic acid rice mutant reveals a mutation in the rice orthologue of maize MIK [J].Theor Appl Genet,2008,117:1291-1301.
[22] Xu X H,Zhao H J,Liu Q L,et al.Mutations of the multidrug resistance-associated protein ABC transporter gene 5 result in reduction of phytic acid in rice seeds[J].Theor Appl Genet,2009,119:75-83.
[23] Ye H X,Zhang Z Q,Broughton S,et al.A nonsense mutation in a putative sulphate transporter gene results in low phytic acid in barley [J].Funct Integr Genomics,2011,11:103-110.
[24] Zhao H J,Liu Q L,HW F U,et al.Effect of non-lethal low phytic acid mutations on grain yield and seed viability in rice[J].Field Crops Res,2008,108:206-211.
[25] Yuan F J,Zhu D H,Deng B,et al.Effects of two low phytic acid mutations on seed quality and nutritional traits in soybean[J].Agricultural and Food Chemistry,2009,57(9):3632-3638.
