ATP 競爭性GSK-3 抑制劑對人食管鱗癌細胞多藥耐藥的影響*
2012-12-23 04:08:00甘思遠鐘雪云謝思明
中國病理生理雜志 2012年7期
關鍵詞:檢測
甘思遠, 鐘雪云, 謝思明, 羅 楓
(1 廣東醫學院病理學教研室,廣東 湛江524023;2 暨南大學醫學院病理學教研室,廣東 廣州510632;3清遠市人民醫院病理科,廣東 清遠511518)
目前臨床上對食管鱗狀細胞癌(esophageal squamous-cell carcinoma,ESCC)主要采用手術切除結合術后放化療的治療方案。隨著腫瘤化療藥物的廣泛應用,腫瘤治療過程中產生的耐藥性問題越來越突出,已成為腫瘤聯合化療失敗的主要原因之一[1]。腫瘤細胞的多藥耐藥(multidrug resistance,MDR)是指腫瘤細胞對多種具有不同結構和作用靶位的抗腫瘤藥物產生耐藥性[2]。研究發現,ATP 結合盒(ATP-binding cassette,ABC)轉運蛋白家族成員:ABCB1(ATP-binding cassette,subfamily B,member 1)基因編碼的P 糖蛋白(P-glycoprotein,P-gp)、ABCC2(ATP-binding cassette,subfamily C,member 2)基因編碼的多藥耐藥相關蛋白2(multidrug resistance-associated protein 2,MRP2)通過水解ATP 獲得能量依賴性的跨膜藥物外排泵的功能,可將抗腫瘤藥物逆濃度從細胞內泵出到細胞外,降低細胞內藥物濃度并導致腫瘤細胞產生多藥耐藥(multidrug resistance,MDR)[3-4]。研究發現抗凋亡蛋白B 細胞淋巴瘤-2(B-cell lymphoma-2,Bcl-2)蛋白在腫瘤細胞中高表達,可阻斷多種化療藥物誘導的腫瘤細胞凋亡[5]。P-gp、MRP2、Bcl-2 與腫瘤多藥耐藥密切相關,在腫瘤化療耐藥的發生發展中起著重要作用。
Yamada 等[6]發現,編碼P-gp 的ABCB1 基因是Wnt 通路效應元件TCF4/β-catenin 復合物的一個直接目的基因,TCF4/β-catenin 途徑的激活可促進P-gp 表達增加。Lim 等[7]研究發現,將與ATP 競爭糖原合成酶激酶3β (glycogen synthase kinase 3β,GSK-3β)催化活性部位的ATP 競爭性GSK-3 抑制劑6-溴靛玉紅-3'-肟(6-bromoindirubin-3'-oxime,BIO)作用于細胞會導致β-catenin、P-gp 和MRP2 的表達增加,β-catenin 在胞漿及胞核內出現累積,P-gp 對其底物的轉運功能增強,同時還激活了ABCB1 基因的啟動子。
本研究將ATP 競爭性GSK-3 抑制劑BIO 作用于ESCC 細胞后,檢測β-catenin 與P-gp、MRP2 和Bcl-2 表達之間的關系,細胞內游離ATP 水平的改變對P-gp 轉運功能的影響,研究GSK-3β 活性和細胞內游離ATP 能量水平的改變與ESCC 的MDR的關系,以期為腫瘤臨床治療尋找新的靶點。
材 料 和 方 法
1 材料
1.1 細胞來源 人類食管鱗癌細胞系EC-109 由汕頭大學醫學院饋贈。
1.2 主要試劑 RPMI-1640 培養液購自Gibco;新生胎牛血清購自杭州四季青公司,CY3 標記山羊抗小鼠IgG、BCA 法蛋白質定量測定試劑盒和ATP 檢測試劑盒均購自上海碧云天公司;鼠抗人MRP2 單克隆抗體購自Abcam;鼠抗人P-gp 單克隆抗體和鼠抗人β-catenin 單克隆抗體和鼠抗人Bcl-2 單克隆抗體均購自Santa Cruz;羅丹明123 染色試劑盒購自南京凱基公司;二甲基亞砜(dimethyl sulfoxide,DMSO)購自北京普博欣公司;BIO 購自Sigma。
2 方法
2.1 細胞培養 人類食管鱗癌細胞EC-109 按常規方法培養于含10%小牛血清、1 ×105U/L 含青霉素和100 mg/L 鏈霉素的RPMI-1640 培養液完全培養基中,置于5% CO2、37 ℃恒溫密閉式培養箱(相對濕度為95%)內,倒置顯微鏡觀察生長情況。約3~4 d 傳代1 次,取對數生長期細胞用于實驗。
2.2 細胞處理 EC-109 細胞常規培養后,取對數生長期細胞制備成細胞懸液,將2.8 ×108/L EC-109 細胞1 mL 接種到培養瓶中,48 h 后根據不同組別分別加入2 mL 含不同濃度BIO 藥液的RPMI-1640 培養液或無BIO 藥液的RPMI-1640 培養液,使BIO 的終濃度分別為1 μmol/L 和2 μmol/L,無藥液組為空白對照組(藥物劑量的選擇參考文獻[7-8])。24 h 后將6 孔板從培養箱內取出,倒置顯微鏡下觀察活細胞形態。
2.3 免疫熒光細胞化學 加入BIO 處理細胞24 h后取出有細胞的蓋玻片,PBS 洗5 min ×3 次,丙酮冰上固定細胞10 min,山羊血清封閉75 min,加入適量按適當比例稀釋的Ⅰ抗(MRP2、P-gp、β-catenin 和Bcl-2 抗體的稀釋濃度均為1∶100),4 ℃冰箱孵育過夜,滴加適量的Cy3 標記Ⅱ抗(稀釋比例1∶100),室溫孵育1 h(避光),緩沖甘油封片,用PBS 代替Ⅰ抗為陰性對照,其它步驟相同。運用專業CCD 圖像采集系統在顯微鏡下對各實驗組的免疫熒光細胞化學結果隨機選取5 個以上的200 倍視野進行圖像采集。應用Image-Pro Plus 6.0 專業圖像分析軟件,對所采集的熒光圖片進行圖像分析,計算陽性區域的平均吸光度(mean absorbance)值;以平均吸光度值代表各蛋白的熒光強度,反映各個蛋白的表達情況。
2.4 羅丹明123 染色試劑盒檢測P-gp 轉運功能
加入BIO 處理細胞24 h 后常規消化細胞脫壁,細胞懸液離心管內離心,各用PBS 洗滌3 次。將細胞制備成細胞懸液重懸于無血清RPMI-1640 培養基中,細胞計數為1 ×109/L。加入羅丹明123 染液,濃度為4 mg/L,37 ℃、5% CO2細胞培養箱孵育20 min。離心后以PBS 洗滌3 次。重懸細胞于培養基中,37 ℃、5% CO2培養60 min。流式細胞儀檢測,激發波長490 nm,發射波長530 nm;熒光顯微鏡觀察,滴加100 μL 上述混合液于載玻片上,激發濾光片波長488 nm,阻斷濾光片波長515 nm 觀察,拍照。
2.5 ATP 檢測試劑盒檢測細胞內游離ATP 濃度
加入BIO 處理細胞24 h 后吸除培養液,以PBS 洗滌3 次。6 孔板每孔加入200 μL 裂解液裂解細胞,使用細胞刮刮凈6 孔板上的細胞,收集細胞碎片和裂解液于離心管。4 ℃、12 000 ×g 離心10 min,取上清,用于后續的測定。把ATP 標準溶液用ATP 檢測裂解液稀釋成0.1 μmol/L、1 μmol/L、10 μmol/L、20 μmol/L 濃度梯度。加100 μL ATP 檢測工作液到檢測孔內。室溫放置3 ~5 min,以使本底ATP 全部被消耗,從而降低本底。在檢測孔內加上50 μL 樣品或標準品,迅速用微量移液器混勻,間隔2 s 后,立即用酶標儀檢測相對發光單位(relative light unit,RLU)值,同時用酶標儀以562 nm 檢測樣品的吸光度。根據標準品所測得的RLU 值制作標準曲線,根據ATP標準曲線計算出樣品內ATP 的量。BCA 法按試劑盒說明書制作蛋白質標準曲線,根據樣品所測得的吸光度計算樣品內蛋白質的量。
3 統計學處理
運用SPSS 13.0 軟件進行統計學分析。數據以均數±標準差(±s)表示,均數比較采用獨立樣本t檢驗或單因素方差分析,以P <0.05 為差異有統計學意義。
結 果
1 免疫熒光細胞化學結果
與對照組相比,BIO 組β-catenin 和Bcl-2 在胞漿中的表達增強,并且β-catenin 出現胞核內累積(P <0.01);MRP2 在胞漿和胞膜中表達增強(P <0.01);P-gp 在胞漿和胞膜中表達減弱(P <0.01),見圖1、2。

Figure 1. The immunofluorescence cytochemistry showing the expression of MRP2,P-gp,β-catenin and Bcl-2 in EC-109 cells(CY3 staining,×200).圖1 免疫熒光細胞化學檢測EC-109 細胞內β-catenin、Bcl-2、MRP2 和P-gp 的表達
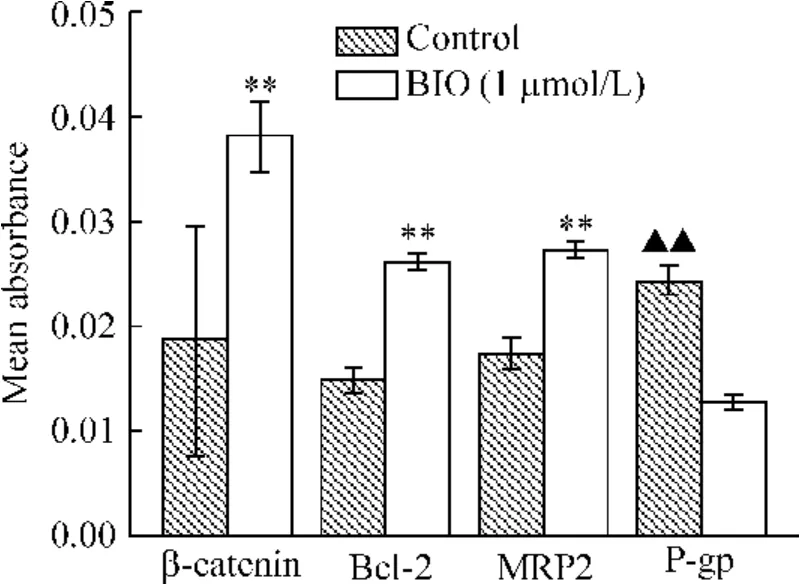
Figure 2. The mean absorbance of immunofluorescence of β-catenin,Bcl-1,MRP2 and P-gp in ESCC cells. ±s.n=3. **P <0.01 vs control group;▲▲P <0.01 vs BIO (1 μmol/L)group.圖2 ESCC 細胞內β-catenin、Bcl-2、MRP2 和P-gp 免疫熒光的平均吸光度值
2 P-gp 對其轉運底物羅丹明123 的轉運功能
與對照組相比,1 μmol/L 和2 μmol/L BIO 處理組細胞內羅丹明123 的熒光強度減弱(P <0.05),而這兩組之間無顯著差別(P >0.05),見圖3。
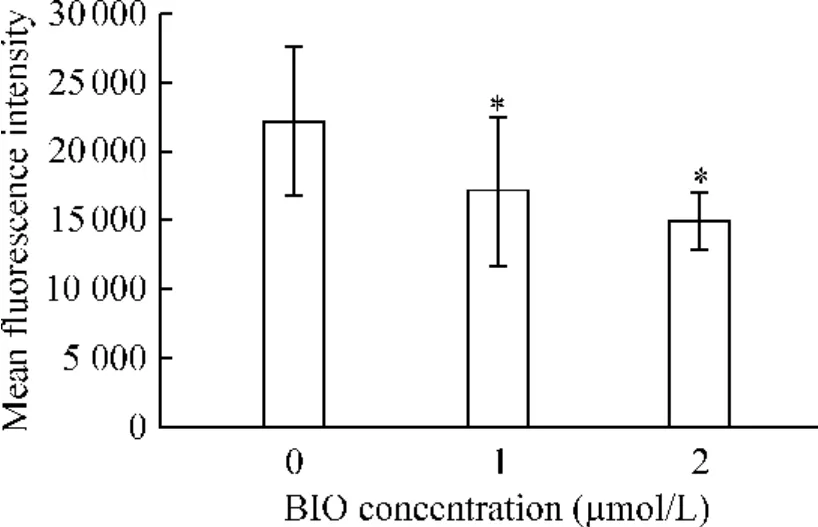
Figure 3. The fluorescence intensity of rhodamine 123 in ESCC cells. ±s.n=4. * P <0.05 vs 0 μmol/L.圖3 ESCC 細胞內羅丹明123 的熒光強度
3 ATP 檢測試劑盒檢測細胞內游離ATP 濃度
2 μmol/L BIO 組細胞內游離ATP 濃度高于1 μmol/L BIO 組和對照組(P <0.05),而1 μmol/L BIO 組與對照組之間無顯著差別(P >0.05),見圖4。
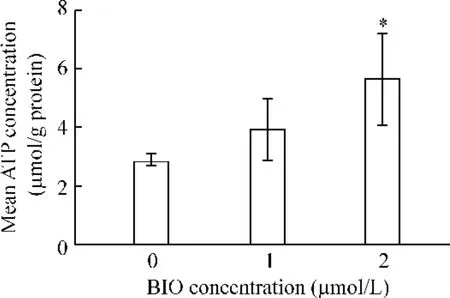
Figure 4. The free ATP concentration in ESCC cells. ±s. n =3. * P <0.05 vs 0μmol/L or 1 μmol/L.圖4 ESCC 細胞內游離ATP 濃度
討 論
β-catenin 是與細胞增殖、分化、凋亡以及腫瘤發生發展密切相關的Wnt 信號通路的關鍵信號元件。β-catenin 在胞漿和胞核的蓄積是Wnt 通路被激活的標志。當Wnt 通路被激活時,GSK-3β 的活性受到抑制,β-catenin 泛素化障礙,β-catenin 在胞漿中累積,并進入細胞核,與淋巴細胞增強子或T細胞因子(lymphoid enhancer factor / T cell factor,LEF/TCF)結合,調節目的基因的表達,促進細胞增殖,對抗凋亡,促進腫瘤的發生發展[9]。本研究運用GSK-3 抑制劑BIO 處理食管鱗癌EC-109 細胞后,BIO 組與對照組相比,β-catenin 在胞漿及胞核中的表達增強,并且β-catenin 在胞核內出現累積,與Lim 等[7]和Chang 等[8]的研究結果相符;提示使用BIO 處理ESCC 細胞后,GSK-3β 的活性被抑制,導致β-catenin 的降解發生障礙,從而使β-catenin在細胞漿及胞核內出現累積。而β-catenin 作為TCF/β-catenin 途徑的關鍵信號元件,其在胞漿及胞核中的表達增強,提示TCF/β-catenin 途徑被激活。同時,運用BIO 處理食管鱗癌EC-109 細胞后,免疫熒光細胞化學的結果顯示,與對照組相比,加藥組抗凋亡蛋白Bcl-2 在胞漿中的表達增強。Yan等[10]在ESCC 細胞的研究中發現,STAT3 是TCF/β-catenin 途徑的靶基因,隨著β-catenin 在胞漿和胞核內的累積,TCF/β-catenin 途徑被激活,從而上調STAT3 的表達;同時已有的研究還證實,bcl-2 是STAT3 的靶基因[11]。因此,本研究運用GSK-3 抑制劑BIO 處理食管鱗癌EC-109 細胞,激活TCF/β-catenin 途徑后,可能上調EC-109 細胞內STAT3的表達,從而間接上調了Bcl-2 的表達。
本研究運用BIO 處理ESCC 細胞后,與對照組相比,加藥組MRP2 在胞漿和胞膜中表達增強,與Lim等[7]的研究結果相符。然而卻意外發現P-gp 在胞漿和胞膜中表達減弱,與Lim 等[7]以及Yamada 等[6]的研究結果不相符;提示在ESCC 細胞中,BIO 處理ESCC 細胞激活TCF/β-catenin 途徑后,P-gp 的表達還受到其它因素的調控。P-gp 表達與TCF/β-catenin 途徑的關系還需繼續進行深入的研究。
本研究運用BIO 處理ESCC 細胞后,分別運用流式細胞術檢測ESCC 細胞內P-gp 轉運底物羅丹明123 在細胞內的熒光強度以及運用ATP 檢測試劑盒檢測ESCC 細胞內游離ATP 水平。流式細胞術結果顯示,與對照組相比,1 μmol/L 與2 μmol/L BIO 組細胞內羅丹明123 的熒光強度減弱,提示使用BIO處理ESCC 細胞后,P-gp 對其作用底物的轉運功能增強,與Lim 等[7]的研究結果相符。而2 μmol/L BIO組內的ESCC 細胞內游離ATP 濃度高于1 μmol/L BIO 組以及對照組的ESCC 細胞內游離A1 μmol/L與2 μmol/L BIO 組TP 濃度,提示2 μmol/L BIO 處理ESCC 細胞后,細胞內游離ATP 水平增加。
1 μmol/L BIO 處理ESCC 細胞后,P-gp 在胞漿和胞膜中表達減弱;同時流式細胞術的結果顯示,P-gp 介導的轉運功能增強。但是由于1 μmol/L BIO組細胞內游離ATP 水平與對照組無顯著差別,因此不能判斷是否是因為使用BIO 處理ESCC 細胞后,BIO 通過與ATP 競爭GSK-3β 的ATP 結合位點,在抑制GSK-3β 活性的同時,通過改變細胞內游離ATP 水平,使ATP 能量依賴性的ABC 轉運蛋白P-gp 的轉運功能增強。細胞內游離ATP 水平的改變與ATP 能量依賴性的P-gp 蛋白轉運功能之間的關系以及ATP 競爭性GSK-3 抑制劑BIO 對其轉運功能的調控機制還需繼續進行深入的研究。
MRP2、P-gp 和Bcl-2 與腫瘤MDR 的發生發展密切相關,在腫瘤MDR 的發生發展中起著重要作用。本研究運用BIO 處理ESCC 細胞后,激活了TCF/β-catenin 通路,增加了MRP2 和Bcl-2 的表達,增強了P-gp 對其作用底物的轉運功能,提示BIO 處理ESCC 細胞后增強了細胞的MDR。這為進一步闡明ABC 轉運蛋白在腫瘤細胞多藥耐藥中的機制提供了新的思路。
[1] Falasca M,Linton KJ. Investigational ABC transporter inhibitors[J]. Expert Opin Investig Drugs,2012,21(5):657-666.
[2] Wind NS,Holen I. Multidrug resistance in breast cancer:from in vitro models to clinical studies[J]. Int J Breast Cancer,2011,2011:967419.
[3] Sun YL,Patel A,Kumar P,et al. Role of ABC transporters in cancer chemotherapy[J]. Chin J Cancer,2012,31(2):51-57.
[4] Sodani K,Patel A,Kathawala RJ,et al. Multidrug resistance associated proteins in multidrug resistance[J]. Chin J Cancer,2012,31(2):58-72.
[5] Patel MP,Masood A,Patel PS,et al. Targeting the Bcl-2[J]. Curr Opin Oncol,2009,21(6):516-523.
[6] Yamada T,Takaoka AS,Naishiro Y,et al. Transactivation of the multidrug resistance 1 gene by T-cell factor 4/beta-catenin complex in early colorectal carcinogenesis[J]. Cancer Res,2000,60(17):4761-4766.
[7] Lim JC,Kania KD,Wijesuriya H,et al. Activation of β-catenin signalling by GSK-3 inhibition increases p-glycoprotein expression in brain endothelial cells[J]. J Neurochem,2008,106(4):1855-1865.
[8] Chang HW,Roh JL,Jeong EJ,et al. Wnt signaling controls radiosensitivity via cyclooxygenase-2-mediated Ku expression in head and neck cance[J]. Int J Cancer,2008,122 (1):100-107.
[9] Lam AP,Gottardi CJ. β-catenin signaling:a novel mediator of fibrosis and potential therapeutic target[J]. Curr Opin Rheumatol,2011,23(6):562-567.
[10] Yan S,Zhou C,Zhang W,et al. β-catenin/TCF pathway upregulates STAT3 expression in human esophageal squamous cell carcinoma[J]. Cancer Lett,2008,271(1):85-97.
[11] Huang S,Chen M,Shen Y,et al. Inhibition of activated Stat3 reverses drug resistance to chemotherapeutic agents in gastric cancer cells[J]. Cancer Lett,2012,315(2):198-205.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
