表皮生長因子在牙齒萌出通道形成中的作用研究
2012-11-05 14:43:36劉宗霞楊春俞
牙體牙髓牙周病學雜志 2012年3期
關鍵詞:小鼠
劉宗霞,王 建,李 紓,楊春俞
(山東:1.濰坊醫學院口腔醫學系,濰坊 261053;2.濰坊口腔醫院,濰坊 261011;3.山東大學口腔醫學院牙周病科,山東省口腔生物醫學重點實驗室,濟南 250012)
牙的萌出是牙胚、牙槽骨以及諸多細胞及其分子相互作用且復雜有序的生理過程。這一過程需要在牙胚的冠方形成萌出通道,即由鄰近牙槽骨吸收形成硬組織通道,并且軟組織通過吸收改建而形成軟組織通道。表皮生長因子(epidermal growth factor,EGF)是參與牙萌出的關鍵性調控因子之一,Cohen(1962)首先發現將EGF注射到新生小鼠體內可以使切牙早熟性地萌出[1],并且Hoath等[2]證實在大鼠出生后0~3 d注射EGF,其促萌效果最佳,但其是否參與軟、硬組織通道的形成尚不清楚。
本研究通過觀察EGF/EGFR(epidermal growth factor receptor,表皮生長因子受體)在牙齒萌出前后口腔黏膜中的表達差異,研究EGF是否參與牙齒萌出通道的形成;通過研究EGF與牙囊細胞中單核細胞趨化蛋白-1(monocyte chemotactic protein-one,MCP-1)的關系,探討其在牙槽骨的吸收,即硬組織通道形成過程中的作用。
1 材料和方法
1.1 主要材料和試劑
出生后不同發育期SPF級BALB/c小鼠、SPF級4~6 d齡Wistar大鼠(山東大學動物中心提供);PV二步法免疫組化檢測試劑盒和DAB試劑盒(邁新生物技術公司);α-MEM培養液(Hyclone,美國);胎牛血清(FBS)(杭州四季青);EGF(Sigma,美國);四唑鹽(MTT)、Trizol(上海生工生物工程有限公司);逆轉錄試劑盒(Fermentas);PCR試劑盒(大連寶生物公司)。
1.2 EGF/EGFR在牙齒萌出過程中表達的組織學定位觀察
1.2.1 標本處理
分別取出生后13、15 d以及牙齒已萌出完成的成年BALB/C小鼠各4只,引頸處死后,分離解剖含下頜第一磨牙的下頜骨,置新鮮配制40 g/L多聚甲醛中固定過夜,根據發育時期的不同,分別用100 g/L EDTA脫鈣液常溫脫鈣1~3月不等。梯度乙醇脫水,石蠟包埋,近中遠中向5 μm連續切片,多聚賴氨酸包被的玻片撈片,60℃烤片2~3 h。HE染色觀察牙齒發育的情況。
1.2.2 免疫組化PV二步法檢測EGF/EGFR的表達
取上述各發育期小鼠的標本,二甲苯脫蠟,梯度水化至水,30 mL/L過氧化氫液室溫孵育5 min,以阻斷內源性過氧化物酶;復合消化液消化5 min,蒸餾水洗,血清封閉,室溫孵育20 min,不洗,滴加一抗(1∶100稀釋)37℃孵育1 h,PBS漂洗;滴加羊抗鼠IgG抗體-HRP多聚體,37℃孵育20 min,PBS漂洗;DAB溶液顯色,蒸餾水沖洗,蘇木素復染,中性樹膠封片,顯微鏡下觀察并照相。陰性對照片用PBS分別代替一抗或二抗。
1.3 EGF對牙囊細胞增殖和MCP-1表達的影響
1.3.1 大鼠牙囊細胞分離培養和鑒定
取出生后4~6 d的SPF級Wistar大鼠,參照Wise等[3]方法進行牙囊細胞分離培養。即大鼠引頸處死后,浸入750 mL/L乙醇2~3 s,體視顯微鏡下分離下頜第一磨牙的牙胚,10 g/L胰蛋白酶液消化1.5 h,在體視顯微鏡下分離牙囊組織。然后將分離的牙囊組織置于培養瓶中,加入含200 mL/L FBS的 α-MEM 培養液,在37℃,50 mL/L CO2以及飽和濕度下培養,每天在倒置顯微鏡下嚴密觀察細胞生長狀況,每3 d換液1次。待細胞單層生長達培養瓶底80%以上時,用胰蛋白酶消化法傳代培養,取第3代牙囊細胞進行角蛋白、波絲蛋白免疫組化染色法鑒定細胞的組織來源,備用。
1.3.2 EGF對牙囊細胞增殖的影響
取生長良好的第3代牙囊細胞以2×104/孔接種于 96孔板,同時加入含 150 mL/L FBS的α-MEM培養液,37℃ 50 mL/L CO2條件下培養24 h,倒置相差顯微鏡下觀察,見大多數牙囊細胞貼壁并伸展后,棄去孔內液體和未貼壁細胞,用無血清α-MEM培養液沖洗3次后,將細胞隨機分為5個實驗組和1個對照組共6組,每組復5孔。5個實驗組分別加入含EGF終末濃度為0.5、1、5、10和50 ng/mL的含5 mL/L FBS的α-MEM培養液,對照組加含5 mL/L FBS的α-MEM培養液,每孔100 μL。然后將96孔培養板放入37℃,50 mL/L CO2的細胞培養箱中繼續培養5 d后,每孔中加入5 mg/mL的MTT 20 μL,繼續培養4 h后,吸棄孔內液體,每孔加入150 μL 的DMSO,振蕩10 min,在分光光度儀490 nm波長下測吸光度(A)值。
1.3.3 RT-PCR 檢測 EGF 對牙囊細胞MCP-1mRNA表達的影響
1.3.3.1 牙囊細胞處理
取生長良好的第3代牙囊細胞,用2.5 g/L的胰蛋白酶液消化后,配成單細胞懸液并接種于直徑為70 mm培養皿中,加入含50 mL/L FBS的α-MEM培養液,37℃,50 mL/L CO2條件下培養。次日去除未貼壁細胞,待細胞生長至匯合點時,將細胞隨機分為4個實驗組和1個對照組。4個實驗組先用無血清的α-MEM培養液培養5 h后,均換用含EGF終末濃度為10 ng/mL的無血清α-MEM培養液,并分別繼續孵育0.5、1、3、6 h 后終止培養。對照組用無血清的α-MEM培養液培養5 h后終止培養。
1.3.3.2 RT-PCR 檢測
上述各組細胞終止培養后,Trizol法提取總RNA,反轉錄合成 cDNA,以 cDNA為模板擴增MCP-1和內參照β-actin。引物由上海博尚生物科技公司合成。
MCP-1分子引物序列:上游引物:5'GCAGGTCTCTGTCACGCTT3',下游引物:5'GTGCTTGAGGTGGTTGTGGAAA3',該引物產生413 bp的cDNA片段。β-actin分子引物序列:上游引物:5'CCCTGAAGTACCCCATTGAA3',下 游 引 物:5'CTTTTCACGGTTGGCCTTAG3',該引物產生158 bp的cDNA片段。采用2 ×Taq PCR MasterMix 25 μL反應體系。反應條件一致為:94℃預變性3 min,94 ℃變性 35 s、54.4 ℃退火35 s、72 ℃延伸1 min,35個循環;最后72℃延伸5 min。
1.3.3.3 PCR 產物鑒定和分析
取8 μL PCR產物經12 g/L瓊脂糖凝膠電泳,溴化已啶染色,DL 2000 Maker 5 μL作參照,80 V電壓下電泳,在紫外線箱中觀察并照相記錄。用UVP凝膠成像系統照相,并用JD801顯微圖像分析系統分析軟件測量其積分光密度值。
1.4 統計學分析
2 結果
2.1 EGF/EGFR的組織學定位
小鼠出生后13 d時,在下頜第一磨牙冠方可觀察到退化的成釉細胞、口腔上皮、固有層,EGFR在其冠方口腔黏膜的全層包括基底層、棘層和角化層呈強陽性表達(圖1),EGF在口腔黏膜上皮呈弱陽性表達(圖2)。小鼠出生后15 d時,牙齒冠方的軟組織融合為實性的上皮細胞團,部分細胞呈現為空泡狀的凋亡狀態(圖3)。此時,EGFR僅在縮余釉上皮處呈強陽性表達,其余部分表達明顯減弱,處于凋亡狀態的細胞無表達(圖4)。而EGF在實性上皮團中呈陰性表達(圖3)。
在牙齒萌出后,EGFR的表達集中于上皮基底層(圖5),EGF的表達集中于口腔黏膜的固有層(圖6)。
2.2 牙囊細胞的培養和鑒定
與葛少華等[4]的實驗結果相似,牙囊組織培養24 h后便可見少數細胞從組織周圍爬出,14~21 d后細胞即可達單層匯合。主要有兩種細胞形態,一種細胞呈梭形,另一種細胞呈多角形,兩種細胞間雜在一起生長。免疫細胞化學染色顯示細胞的波形蛋白表達陽性(圖7),角蛋白表達陰性(圖8),表明細胞源自間充質。
2.3 EGF對牙囊細胞增殖的影響(表1)
EGF對牙囊細胞增殖的影響在濃度為5 ng/mL時開始顯效,10 ng/mL時達到最高,兩者與對照組相比差異均有統計學意義(P<0.05)。50 ng/mL時增殖活性開始降低,且與對照組無顯著性差異(P >0.05)。
表1 EGF對牙囊細胞增殖的影響(A490,)
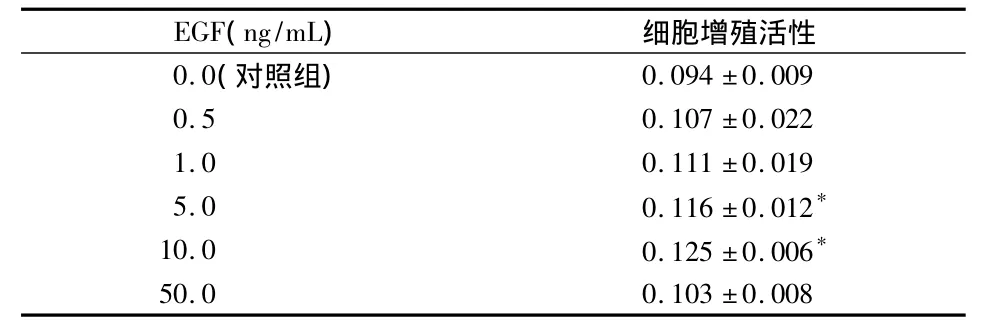
表1 EGF對牙囊細胞增殖的影響(A490,)
*與對照組比較 P<0.05
EGF(ng/mL)細胞增殖活性0.0(對照組)0.094 ±0.0090.5 0.107 ±0.0221.0 0.111 ±0.0195.0 0.116 ±0.012*10.0 0.125 ±0.006*50.0 0.103 ±0.008
2.4 EGF對體外培養的牙囊細胞MCP-1 mRNA表達的影響
PCR產物測序結果與基因庫中公布的MCP-1(M -57441)序列100%一致。MCP-1、β-actin 的擴增產物分別為413、158 bp(圖9)。牙囊細胞與10 ng/mL EGF 共同孵育0.5 h,牙囊細胞中MCP-1 mRNA表達開始升高,3 h表達最高,以后逐漸恢復,但仍比對照組高,即共同孵育 0.5、1、3、6 h 各時間點MCP-1 mRNA表達量均明顯高于對照組,差異有統計學意義(P<0.05)(圖10)。
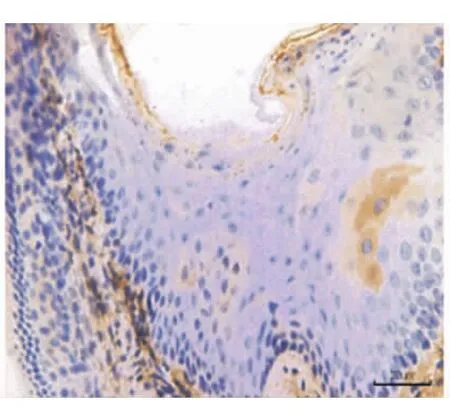
圖2 13 d小鼠,EGF在下頜第一磨牙冠方上皮層呈弱陽性表達(×20)
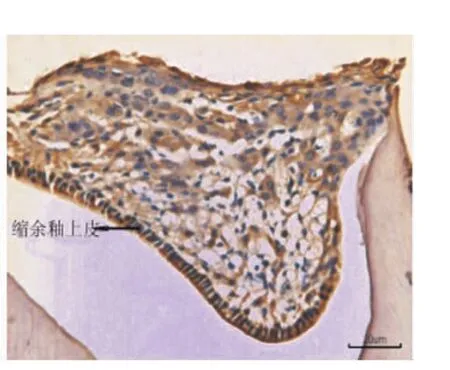
圖4 15 d小鼠,EGFR在縮余釉上皮中呈陽性表達,在實性上皮團其余部分表達明顯減弱(×20)
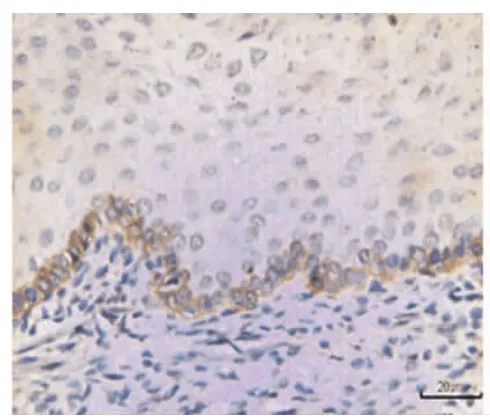
圖5 牙齒萌出完成后,EGFR在牙齦上皮基底層呈陽性表達,在固有層中無表達(×20)
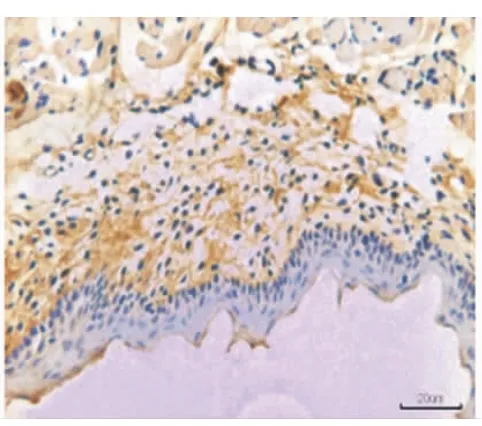
圖6 牙齒萌出完成后,EGF在牙齦固有層呈陽性表達,在上皮層中無表達(×20)
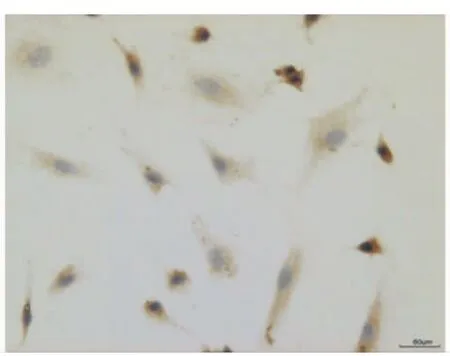
圖7 牙囊細胞抗波形絲蛋白染色呈陽性(棕黃色),位于細胞漿中,胞核著色陰性(免疫細胞化學染色,×20)
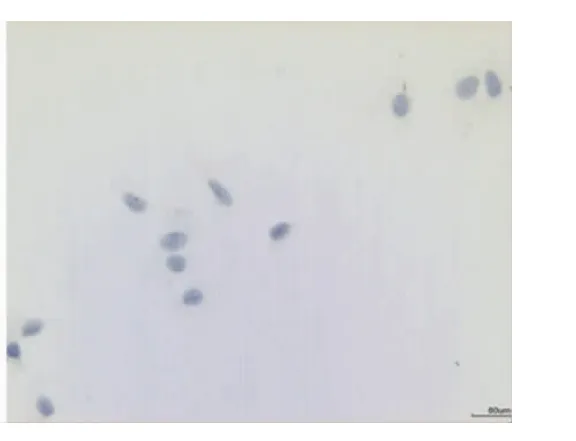
圖8 牙囊細胞抗角蛋白染色呈陰性,藍色為蘇木素復染的細胞核(免疫細胞化學染色,×20)
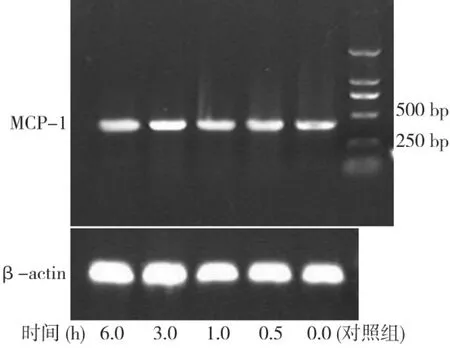
圖9 EGF對牙囊細胞MCP-1表達的影響
3 討論
牙齒的萌出是在牙冠形成后,牙胚向牙合面移動,穿過骨隱窩和口腔黏膜,直至達到功能位置的一個復雜的過程。這一過程可分為3個時期:萌出前期、萌出期和萌出后期(或功能期)[5]。其中,萌出前期的主要變化是牙根形成時牙胚在牙槽骨中的移動,這一過程需要相應部位的牙槽骨發生吸收和改建,以形成利于牙萌出的硬組織通道。在萌出期,覆蓋在牙冠表面的縮余釉上皮分泌多種酶,溶解結締組織,形成一個有上皮襯里的軟組織通道。因此我們可以認為軟、硬組織通道的建立是牙得以順利萌出的關鍵所在。
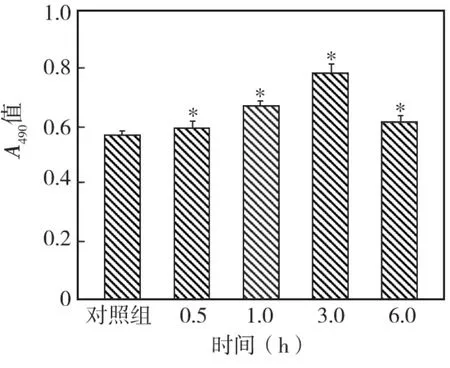
圖10 EGF作用后牙囊細胞MCP-1mRNA表達的積分光密度值(*與對照組相比P<0.05)
EGF屬于小分子多肽,是細胞生長因子的一類,與其受體結合,在靶細胞內產生一系列生物學效應,包括促進表皮生長和角化[6-7],促進 DNA合成,引起細胞增殖等。EGF是參與牙萌出的重要調控因子之一,將其注射到新生大鼠體內,可有效促進磨牙的萌出[2],但其具體的促萌機制尚不清楚。
Huvsseune等[8]通過光鏡和電子顯微鏡觀察斑馬魚第一副牙齒萌出過程,認為上皮的改建在牙萌出過程中發揮著重要的作用。牙萌出時,縮余釉上皮分泌酶,溶解結締組織,加之萌出時上皮對結締組織的壓力,使結締組織破壞。這時,縮余釉上皮外層細胞和口腔上皮細胞增殖并移動到退變的結締組織處,在萌出牙的上方融合形成上皮團,繼而上皮團中央細胞死亡,形成一個有上皮襯里的牙萌出通道,通過該通道,牙萌出時不會發生出血[5]。本研究在15 d小鼠下頜第一磨牙的冠方觀察到實性的上皮細胞團,符合上述理論。同時還發現,13 d時EGF在小鼠下頜第一磨牙冠方的口腔黏膜上皮層中呈弱陽性表達,而其在成年鼠的口腔黏膜中的表達僅限于固有層。Wise等[9]報道,在剛出生大鼠體內注射EGF可以增加星網狀層細胞IL-1α的表達,提示在體外EGF也可促進星網狀層細胞 IL-1α以及 IL-1α mRNA的表達。Modeer等[10]研究發現,EGF可促進 IL-1誘導前列腺素E2(PGE2)的生成。PGE2可以抑制成纖維細胞的增殖及膠原的合成,并使基質金屬蛋白酶(MMPs)增加,從而促進結締組織的溶解吸收。上述研究結果提示EGF可促進結締組織的退變吸收。本研究發現,牙齒萌出后,EGFR表達于口腔黏膜上皮的基底層,隨細胞向棘層分化而逐漸消失。而出生13 d時,EGFR在口腔黏膜的全層呈強陽性表達,高水平表達的EGFR明顯增強了上皮細胞對生長因子的反應性,促進它與受體結合形成復合物,這樣勢必促進了口腔黏膜上皮的增殖分化進程。這進一步證實了牙齒萌出時,其冠方結締組織的退化吸收,上皮的增殖改建是實性上皮團得以順利形成的關鍵。出生15 d時,在實性上皮團中,EGF呈陰性表達,EGFR僅在退化的成釉上皮處維持強陽性表達,其余部分表達明顯減弱,處于凋亡狀態的細胞無表達。表明EGF/EGFR的相互作用集中于上皮改建的初期,即EGF可能主要在實性上皮團的形成過程中發揮著積極的作用。由此可見EGF可能通過間接誘導PGE2的合成,促進結締組織的溶解吸收,并且可促進冠方上皮的增生融合,參與了上皮團的形成,這可能在軟組織通道的形成中發揮著重要的作用。
單核細胞進入牙囊并在其中聚集是牙萌出的關鍵[11],此后單核細胞轉變成破骨細胞,破骨細胞吸收牙槽骨以形成牙齒萌出的硬組織通道。本研究發現:EGF具有促進體外培養的牙囊細胞增殖的作用,且呈劑量依賴性,5~10 ng/mL EGF可顯著促進牙囊細胞的增殖,10 ng/mL時增殖活性最強,50 ng/mL時增殖活性開始減弱。推測EGF可能通過促進牙囊細胞中EGFR的表達,并與之結合,從而有效的促進了牙囊細胞的增殖。EGF是否可通過促進牙囊細胞的有絲分裂而參與牙的萌出有待進一步研究,但現在至少可以證實EGF在維持牙囊細胞的增殖活性方面發揮著積極的作用。
選擇EGF的較佳效應濃度10 ng/mL來檢測其對MCP-1的影響,結果發現10 ng/mL的EGF具有促進MCP-1表達的作用,共同培養0.5 h開始起效,在3 h表達增強最顯著,以后逐漸減弱。MCP-1表達的降低可能是由于大量外源性的EGF抑制了牙囊細胞表面EGFR的表達,受體表達下降會減弱EGF上調MCP-1轉錄和翻譯的過程。Que等[12]研究證實,在大鼠體內注射EGF同樣可促進牙囊細胞中MCP-1的表達,進一步證實了EGF對于MCP-1的上調作用。EGF通過促進牙囊細胞分泌MCP-1,誘使單核細胞遷入牙囊,從而促進硬組織通道的形成。
牙萌出是牙胚和周圍牙槽骨內發生的多種與牙萌出相關的信號分子共同參與的復雜的激聯過程[13],可能存在多種通路,EGF可能在軟、硬組織通道的形成過程中發揮著積極的作用,但其具體的過程仍不清楚,有待進一步的研究。
[1]Cohen S.Isolation of mouse submaxillary gland protein accelerating incisor eruption and eyelid opening in the new-born animal[J].J Biol Chem,1962,237:1555-1562.
[2]Hoath SB.Treatment of the neonatal rat with epidermal growth factor:differences in time and organ response[J].Pediatr Res,1986,20(5):468-472.
[3]Wise GE,Lin F,Fan W.Culture and characterization of dental follicle cells from rat molars[J].Cell Tissue Res,1992,267(3):483-492.
[4]葛少華,李德懿,楊丕山.小鼠牙囊細胞的體外分離培養鑒定及異質性研究[J].上海口腔醫學,2004,13(6):506-509.
[5]于世鳳,汪說之.口腔組織病理學[M].北京:人民衛生出版社,2004.
[6]Cohen S.The stimulation of epidermal proliferation by a specific protein(EGF)[J].Dev Biol,1965,12(3):394 -407.
[7]Steidler NE,Reade PC.Histomorphological effects of epidermal growth factor on skin and oral mucosa in neonatal mice[J].Archs Oral Biol,1980,25(1):37 -43.
[8]Huvsseune A,Sire JY.The role of epithelial remodelling in tooth eruption in larval zebrafish[J].Cell Tissue Res,2004,315(1):85-95.
[9]Wise GE,Lin F,Zhao L.Immunolocalization of interleukin-1 alpha in rat mandibular molars and its enhancement after in vivo injection of epidermal growth factor[J].Cell Tissue Res,1995,280(1):21-26.
[10]Modeer T,Yucel-Lindberg T,Iinuma M,et al.Epidermal growth factor potentiates interleukin 1 and tumor necrosis factorinduced prostaglandin biosynthesis in human gingival fibroblasts[J].Cytokine,1993,5(3):198 -204.
[11]Nakchbandi IA,Weir EE,lnsogna KL,et al.Parathyroid hormone-related protein induces spontaneous osteoclast formation via a paracrine cascade[J].Proc Natl Acad Sci USA,2000,97(13):7296-7300.
[12]Que BG,Wise GE.Tooth eruption molecules enhance MCP-1 gene expression in the dental follicle of the rat[J].Dev Dyn,1998,212(3):346-351.
[13]Wise GE,Frazier-Bowers S,D'Souza RN.Cellular,molecular,and genetic determinants of tooth eruption[J].Crit Rev O-ral Biol Med,2002,13(4):323-334.
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
