2006-2010年中國鼠傷寒沙門菌分子分型分析*
2012-09-26 03:23:30陳建才樊粉霞王淑京刁保衛趙英偉閆梅英
中國人獸共患病學報 2012年12期
陳建才,樊粉霞,王淑京,婁 靜,刁保衛,闞 飆,趙英偉,閆梅英
鼠傷寒沙門菌經常引起人類胃腸炎或食物中毒,是常見食源性病原菌之一。隨著生產生活模式的改變,國家之間和地域之間人員、食品往來、物資流通更加廣泛,每年世界范圍包括我們國家在內都會有沙門菌引起的食物中毒事件的報道,特別在歐美等發達國家,沙門菌污染食品致腹瀉暴發由以前的點源性集中暴發,越來越多地轉變為跨地區、跨州(省)的‘散在暴發’形式出現,使污染源及傳播途經的發現越來越依賴于分子分型手段的輔助及確認[1-3]。對于鼠傷寒沙門菌,目前已有比較成熟的國際通用標準化 PFGE(Pulse-Field Gel Electrophoresis,脈沖場凝膠電泳)分型方案及第二代分子分型技術 MLVA(Multiple Loci VNTR Analysis,多位點串聯重復序列分析),并成功應用于多起食源性疾病暴發特別是以實驗室為基礎的暴發的發現及溯源調查、確認[4-5]。
在我國,鼠傷寒沙門菌和腸炎沙門菌是臨床最常見的沙門菌致病血清型[6-8]。由這兩種致病菌引起的集中點源暴發由于有清楚的流行病學背景資料,很容易識別,PFGE僅作為后續的補充確認手段,在我國是否存在類似發達國家的由散在病例構成的暴發尚需進一步證實。而這種暴發需要實驗室及時的PFGE或MLVA分型分析,為提升我國實驗室能力,建立以實驗室為基礎的監測,完善我國分子分型網絡數據庫,本研究對近5年我國不同省份來源的294株鼠傷寒沙門菌,進行PFGE及MLVA分型分析,初步建立我國鼠傷寒沙門菌分子分型數據庫,了解其分子遺傳特征,為該菌可能引起的暴發發現提供本底發病水平。
1 材料與方法
1.1 菌株來源 鼠傷寒沙門菌共294株,其中來源于2006年至2010年北京、上海、四川、廣東、河南、湖北、重慶7省(直轄市)CDC沙門菌監測項目中收集的272株病人菌株,13株來源于食物皮蛋,5株來源于其他食品,2株來源于體檢人群,來源于污水和鴨糞各1株。
1.2 PFGE分型 所有鼠傷寒沙門菌參照國際實驗室分子分型監測網絡PulseNet中沙門菌PFGE分型標準化方案進行操作。使用限制性內切酶XbaⅠ進行酶切,獲得的菌株PFGE圖像錄入BioNumerics(Version5.1,Applied maths,Inc.)軟件包進行處理,經校準后,使用非加權配對算術平均法(unweighted pair group average method,UPGMA)進行聚類,構建聚類樹,分析菌株間的相似性。
1.3 MLVA分型 參照國際實驗室分子分型監測網絡PulseNet公布的鼠傷寒沙門菌MLVA分型標準方案,采用7個VNTR位點,合成相應引物(表1),并在引物5′端標記熒光基團,對294株鼠傷寒沙門菌進行PCR擴增,擴增產物使用ABI3700測序儀進行毛細管電泳檢測,根據擴增片段大小計算每個VNTR位點的重復數。并將數據錄入BioNumerics軟件包進行分析。構建MLVA分型結果的最小生成樹(minimum spanning tree,MST),構建參數為creation of complex:1changes。
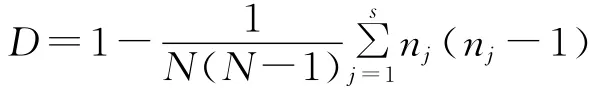
2 結 果
2.1 鼠傷寒沙門菌PFGE分子分型特征 294株鼠傷寒沙門菌經XbaⅠ酶切,PFGE分析后,獲得87種帶型,分型的D值為0.905 0,每種帶型包含176株菌株(圖1)。3株(含)以上帶型有13種,帶型頻率分布見表2。其中優勢帶型JPXX01.CN0001 76株,主要包括重慶8株、四川67株、河南1株,分別來自不同年代的腹瀉病人糞便73株、皮蛋中分離13株;次優勢帶型JPXX01.CN0006 40株,分布于廣東、河南、上海、四川、湖北;24株第3優勢帶型JPXX01.CN0073型,除2株來源于湖北外,其余皆為河南菌株。此外,來自河南的135株菌中54株和北京2009年的9株各自明顯聚集成一簇(圖1中虛線框所示),提示菌株具有一定的地域聚集性,但其他地區來源的菌株則未發現此現象。
2.2 鼠傷寒沙門菌MLVA分型特點 應用7個VNTR位點的MLVA進行分析,獲得198種型別(圖2),平均每種帶型包含1.5株菌。3株(含)以上帶型有19種,其中優勢帶型18 513株,均分離自河南2009年的患者菌株;其次優勢帶型03 312株,主要為2008年河南病人菌株;再次優勢帶型為05 311株,均來自四川2008年,包括7株皮蛋分離菌株,4株病人糞便中分離菌株。
利用軟件BioNumerics對MLVA分型結果進行最小生成樹分析,最小生成樹根據所有VNTRs位點譜號構成生成,最小生成樹將所有的MLVA型分為兩群(圖2):A群和B群。A群包括152個MLVA型,其中75個型別親緣關系較近,形成一簇,構成A群中較大的亞群,主要包括四川腹瀉病人和皮蛋分離株,其次還有河南病人分離株,其他型別相對親緣關系較遠,與菌株來源的散在性相符。
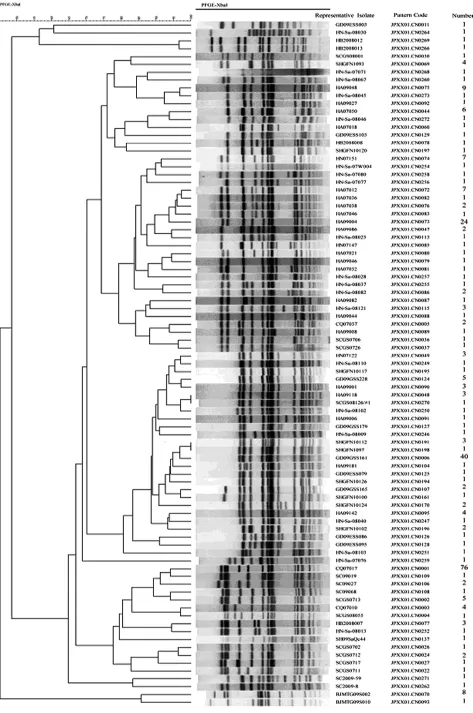
圖1 294株鼠傷寒沙門菌XbaⅠ酶切PFGE分型結果Fig.1 The PFGE patter n with XbaⅠdigestion within 294 S.typhimurium strains
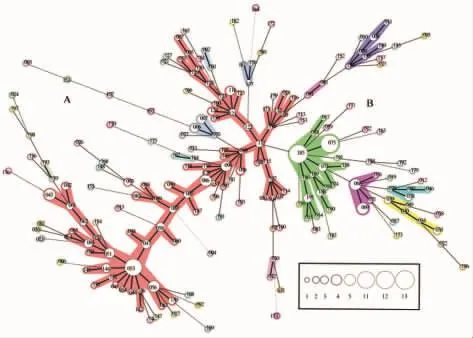
圖2 294株鼠傷寒沙門菌的MLVA分型結果Fig.2 MLVA molecular typing of 294 S.typhi murium isolatesSizes of circles and arcs reflect numbers of isolates.The pattern codes of MLVA are labeled in the circles.The relationships of 294 S.typhi murium isolates are labeled by different colours.
B群分為4個亞群,包括46個MLVA型。B群菌株主要為河南分離株,占74.12%,與PFGE聚類結果一致。
2.3 PFGE與MLVA分型的相互對照和相關聯系2.3.1 PFGE-XbaⅠ單酶切分型方法對鼠傷寒沙門菌的分辨能力不及MLVA分型方法 294株鼠傷寒沙門菌PFGE-XbaⅠ分型的D值為0.9050。MLVA分型的D值為0.992 9,分辨能力高于PFGE,這與 Mia T等的報道一致[2]。除ST3位點 D值為0.221 6,其他VNTR位點均具有較高的多態性,D 值 分 別 為 0.638 8(ST2)、0.821 0(ST5)、0.837 7(ST6)、0.605 6(ST7)、0.626 2(ST8)、0.767 8(STTR10)。
2.3.2 MLVA分型方法的分型能力與兩種酶切的PFGE幾乎相同;MLVA分型方法更有利于聚集性事件的判定 294株鼠傷寒沙門菌經PFGE-XbaⅠ分析后,獲得87種帶型,對其中的主要優勢帶型JPXX01.CN0001菌株76株、JPXX01.CN0006菌株40株和JPXX01.CN0073菌株24株進一步用第2種內切酶Bl nⅠ酶切后,分別獲得10、22和17種帶型。其中優勢帶型JPXX01.CN0001 76株,進一步利用Bl nⅠ酶切(PFGE-Bl nⅠ),獲得10種帶型(表3),JPXA26.CN0029為優勢帶型,包含39株菌。其次為包含13株的JPXA26.CN0035型。MLVA將這76株菌分為44個型,其中9株053型、4株051型和5株043型均對應于PFGE-Bl nⅠ的部分JPXA26.CN0029型及分別對應于部分JPXA26.CN0035、部分JPXA26.CN0074和部分JPXA26.CN0033型;從表2不難看出,除JPXA26.CN0035型、JPXA26.CN0033和JPXA26.CN0074與MLVA分型基本一致外,其他PFGE-Bl nⅠ菌株進一步被MLVA區分為不同型別。當然,也有例外,如JPXA26.CN0029和JPXA26.CN0035則被歸為同一個 MLVA 053型別,共9株,均來自四川同一沙門菌監測網點,分離時間為2008年,具有一定時間地域聚集性,其中屬于同一型別的7株來自食物皮蛋中毒,聯系流行病學資料,進一步證實了兩種分型方法的一致性和可靠性。
2.3.3 PFGE可能提示菌株來源于往年暴發,MLVA更能揭示細菌基因組間的差異 294株鼠傷寒沙門菌經PFGE-XbaⅠ分析后,對其中3株(不含)以上的相同帶型進一步用第2種內切酶Bl nⅠ酶切后,即PFGE-XbaⅠ-Bl nⅠ雙酶切分型。分型條帶總共有159型,D值為0.976 6,接近 MLVA分型區分水平,進一步說明前面所述。統計分型,經過2種酶切后,3株(不含)以上條帶相同的有11個帶型,見圖3(橫坐標為PFGE-XbaⅠ-Bl nⅠ雙酶切帶型與來源地區,縱坐標為菌株數)。從圖3中不難看出,重 慶、四 川 地 區 JPXX01.CN0001-JPXA26.CN0029即0001-0029(ChongQing)和0001-0029(Si-Chuan),重慶可能為局限性暴發,四川2007-2009年都有相同PFGE帶型出現,且2008和2009年均比2007年出現菌株數要多,呈現時間地域聚集,后兩年的菌株聚類現象,可能是2007年延續至2009年,散發菌株引起的沙門菌病暴發。MLVA更能揭示細菌基因組間的差異。比如0070-0120(BeiJing)歸屬同一個PFGE帶型,結合流行病資料,菌株來自同一時間地區的暴發;而MLVA分型卻顯示相似性比較高的8種型別,揭示了菌株基因組間的可變位點的微小差別,比PFGE分型更加精細。綜上所述,在流行病學上,MLVA分型方法的分型能力與雙酶切的PFGE分型相似,考慮到MLVA在操作上較PFGE更方便快捷的特點,故在判斷由鼠傷寒沙門菌引起的聚集性事件時,采用需時較短、操作更方便的MLVA分型方法較兩種內切酶的PFGE分型更有利于及時監測潛在或散在病例形式的疾病暴發。
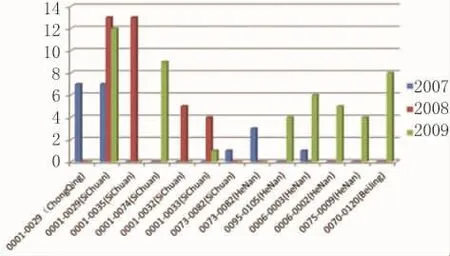
圖3 3株(不含)以上相同PFGE-XbaⅠ-BlnⅠ雙酶切帶型分布Fig.3 The distribution of the same PFGE-XbaⅠ-BlnⅠdouble enzyme digestion patterns

表1 實驗用VNTR位點及其引物Tab.1 The applied VNTR loci and the corresponding primers
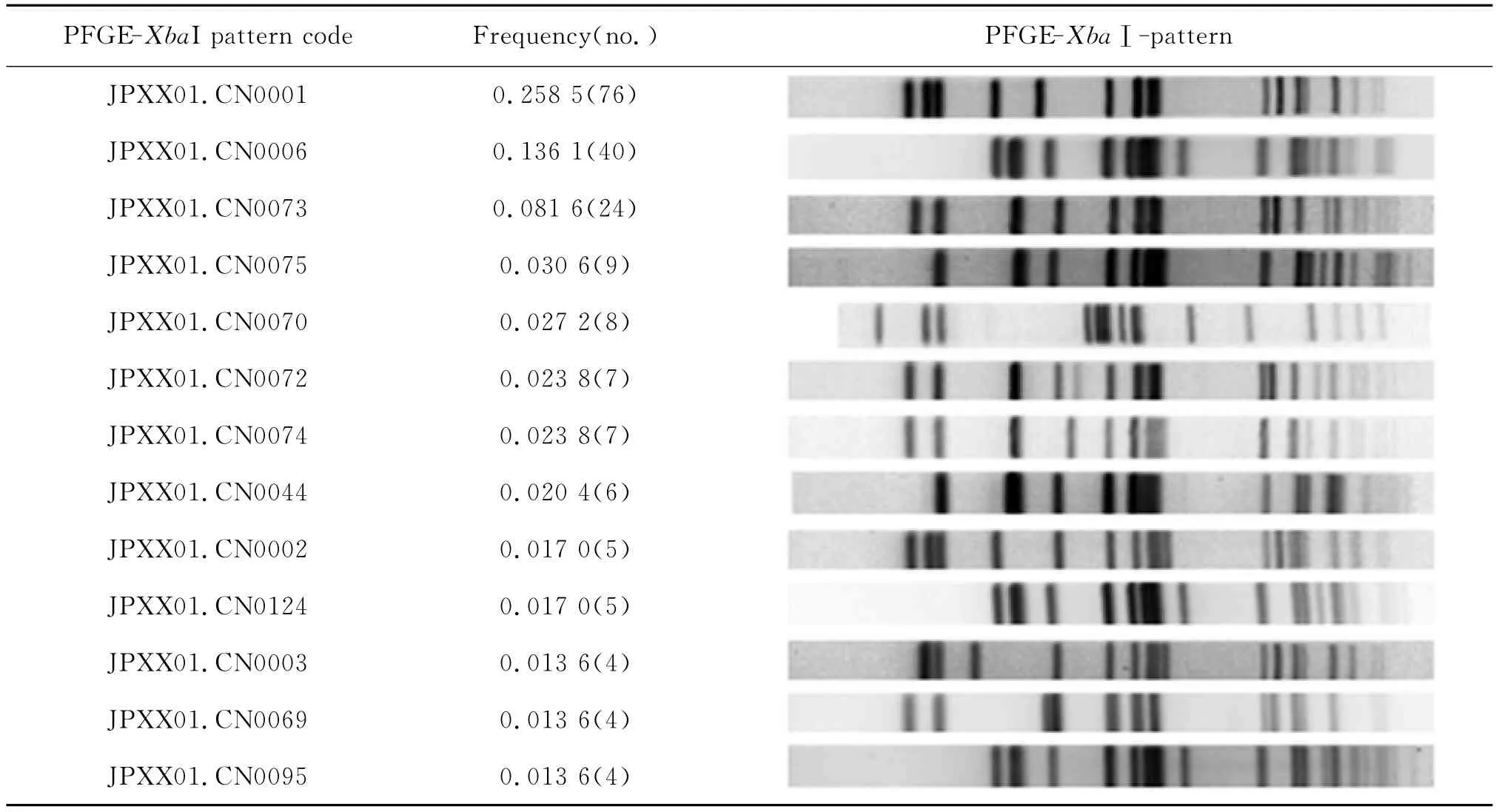
表2 鼠傷寒沙門菌PFGE-XbaⅠ帶型頻率分布Tab.2 PFGE-XbaⅠpattern frequency of S.typhimurium
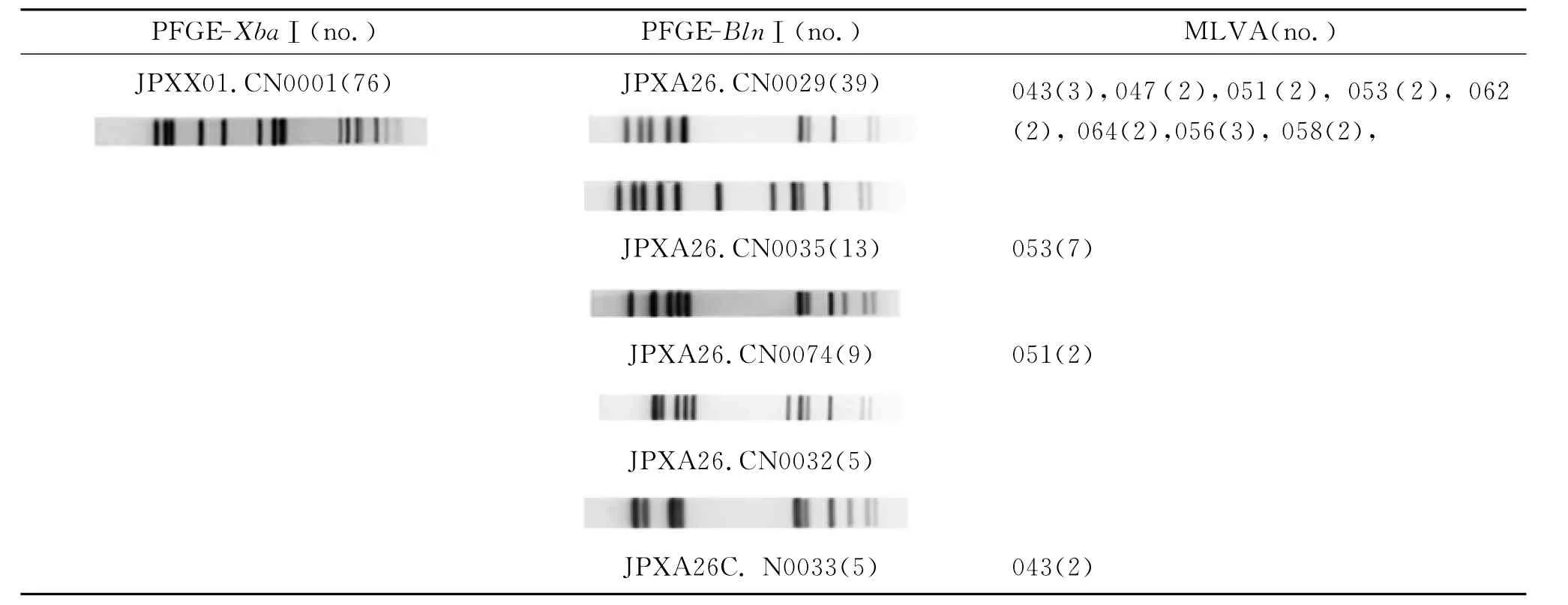
表3 部分鼠傷寒沙門菌分子分型結果及其相對一致性Tab.3 Molecular typing and their compatibility within some S.tphimurium isolates
3 討 論
全球化及食品貿易國際化帶來的食品分發途徑的改變,使食源性疾病發生逐漸由暴發轉為多點散發,單純血清型的分型鑒定已不能滿足在復雜的食源性疾病診斷中對病原體進行溯源的需求。鼠傷寒沙門菌是非傷寒沙門菌性腹瀉或食物中毒中最常見的血清型之一,經常被用于小鼠感染模型模擬人類傷寒發病過程或致病機理研究,由其引起的暴發幾乎每年都有報道[1-8]。同時由于鼠傷寒沙門菌廣泛存在于外界環境中,形成一定的環境、動物、動物性食品帶菌狀態及帶菌率,人食用一定數量的病原菌后導致疾病,如何判定暴發病例感染的鼠傷寒沙門菌與食物及環境中的鼠傷寒沙門菌為一個克隆來源,需要相對精確地分子分型手段輔助,及對鼠傷寒沙門菌在人群中的本底發病水平的了解。隨著分子生物學技術的不斷發展,從DNA水平對致病菌進行分型及鑒定的技術也日趨成熟。
沙門菌監測必須通過細致的流行病學調查以及對病原體的監測分析才能揭示內在的聯系,當前的傳染病控制中,對病原體不僅僅局限于檢測,而是要成為監測的重要內容,形成網絡化,并結合流行病學信息。自1996年發展至今,食源性疾病分子分型網絡已將細菌染色體DNA大片段的長度多態性分析、PFGE、計算機技術3者有機結合,是高技術、標準化、網絡化、能夠資源共享的網絡。PFGE作為大多數細菌分子分型的金標準在暴發的發現及確認中得到了廣泛的關注及應用。實驗室網絡工作人員依據已經完成的鼠傷寒沙門菌PFGE圖譜,進行聚類分析,并與以往本底資料比較,發現新的聚集性PFGE圖譜,或與往年同時期發病水平比較發現2倍(含2倍)以上增高時,通知流行病學人員進行現場調查分析,追蹤可疑食品,并獲得相同PFGE帶型的食品來源菌株,最終確認暴發。因此,沙門菌引起的食源性疾病的暴發在很大程度上依賴于分子分型。分子分型區分能力是指分辨兩種沒有關聯的亞型的能力。實驗中采用Si mpso m指數的D值計算方法,它可以用于評價兩種或多種分型方法對菌株分型能力的高低。應用該指數也可以評價區分散發的菌株與暴發的菌株來源的分辨能力[10],D值取值范圍為0~1,D值越大說明該分型方法的分辨力越高。多數食源性病原菌已有標準化PFGE操作方案,但對于血清型別復雜的沙門菌來說,使用一種PFGE方法存在一定的局限性,如PFGE很難區分不同來源的腸炎沙門菌,鼠傷寒沙門菌單獨使用一種限制性內切酶(XbaⅠ)在判定菌株聚集性時可靠性較差,最好同時使用兩種酶切(XbaⅠ與Bl nⅠ)。但采用雙酶切時,不僅導致試驗時間延長,而且增加試驗成本。加之PFGE需要特定的試劑、儀器設備、軟件及操作熟練的技術人員,這些都在一定程度上制約了PFGE的廣泛推廣。由于上述問題,MLVA作為第二代分子分型技術,以其操作方便、快捷、易檢測的特點顯示出一定的優勢。對于鼠傷寒沙門菌,我們的結果顯示MLVA的區分能力明顯高于單酶切PFGE,與雙酶切PFGE幾乎相同,且兩種分型結果存在一致性,進一步驗證了MLVA分型方法的適用性及可靠性。
為完善我國沙門菌分子分型網絡數據庫,建立可用于暴發識別的基本資料,本研究利用PFGE和MLVA,同時對我國294株分離自病人及皮蛋的鼠傷寒沙門菌進行分子分型分析。結果顯示PFGEXbaⅠ單酶切后的優勢帶型JPXX01.CN0001占25.8%,分離自2007-2009年,且主要集中于四川及重慶。次優勢帶型JPXX01.CN0006(占13.6%)為我國特有PFGE帶型(通過國際Pulse Net數據庫比對),雖然首次在廣東發現該菌型異常增高,但通過加強監測及其他省市菌株PFGE分析發現,該型別在河南省2007年即存在,且雙酶切帶型一致,菌株分離時間相近(一個月內),提示可能存在暴發,但由于PFGE分型的延遲,未能及時開展污染源及傳播途徑的流行病學調查。同樣,對于JPXX01.CN0073型,也存在類似的情況,也未能及時開展流行病學調查。這些均提示實驗室應加強菌株分子分型的及時性。另一方面,這些型別的持續存在也說明污染源的持續存在,應加強監測及數據的及時分析,防止可能的大規模跨區域暴發。
此外,由于缺少詳細的流行病學調查數據,有些菌株缺少具體的暴發地點、來源、分離時間。當實驗室數據顯示有聚集性時,由于缺少足夠的現場證據,不能做出客觀的結論。如圖3中重慶、四川出現相同的 PFGE 雙 酶切帶型0001-0029(XbaⅠ -Bl nⅠ),分子分型呈現菌株聚集性,由于流行病學證據不足,缺少說服力;可能是2007年延續到2010年散發或者散發引起的暴發,對進一步追溯傳染源有一定困難。可見,在傳染病疫情發生時,搞好流行病現場調查,對預防和控制疫情顯得尤為重要,并為今后的疫情防控提供借鑒。否則,實驗室分子分型網絡,只是理論研究。流行病學調查與實驗菌株分析的有效結合,是現在傳染病預防控制的模式,而且新傳染病的發現也越來越多地依靠實驗室數據。因此在我國,應加強流行病學調查與實驗室的溝通,流行病學調查與實驗室檢測同步并行,才能在食源性疾病暴發的早期發現、早期干預方面有所突破。
[1]Chiou CS,Hung CS,Tor pdahl M,etal.Develop ment and evaluation of multilocus variable number tandem repeat analysis for fine typing and phylogenetic analysis of Sal monell a enterica serovar typhimurium[J].Int J Food Microbiol,2010,142(1/2):67-73.DOI:10.1016/j/ijt ood micr o.2010.06.001
[2]Tor pdahl M,Sorensen G,Lindstedt BA,etal.Tandem repeat analysis for sur veillance of hu man Sal monella t yphimurium infections[J].Emer g Infect Dis,2007,13(3):388-395.DOI:10.3201/eid1303.060460
[3]Lindstedt BA,Heir E,Gjernes E,etal.DNA fingerprinting of Sal monell a enterica subsp.enterica serovar typhi murium with emphasis on phage type DT104 based on variable number of tandem repeat loci[J].J Clin Microbiol,2003,41(4):1469-1479.DOI:10.1128/JCM.41.4.1469-1479.2003
[4]Fuller CC,Jawahir SL,Leano FT,etal.A multi-state Sal monell a typhimurium outbreak associated with frozen vacuu mpacked rodents used to feed snakes[J].Zoonoses Public Health,2008,55(8/10):481-487.
[5]Malorny B,Junker E,Hel muth R.Multi-locus variable-number tandem repeat analysis for outbreak studies of Sal monell a enterica serotype Enteritidis[J].BMC Microbiol,2008,8:84.DOI:10.1186/1471-2180-8-84
[6]Torpdahl M,Sorensen G,Ethelberg S,etal.A regional outbreak of S.typhimurium in Den mar k and identification of the source using MLVA typing[J].Euro Surveill,2006,11(5):134-136.
[7]Ran L,Wu S,Gao Y,etal.Laboratory-based surveillance of nontyphoidal Sal monella infections in China[J].Foodborne Pathog Dis,2011,8 (8):921-927. DOI:10.1089/f pd.2010.0827
[8]Deng X,Ran L,Wu S,etal.Laboratory-based surveillance of non-typhoidal Sal monell a infections in Guangdong Province,China[J].Foodborne Pathog Dis,2012,9(4):305-312.DOI:10.1089/f pd.2011.1008
[9]Hunter PR,Gaston MA.Nu merical index of the discri minator y ability of typing syste ms:An application of Si mpson's index of diversity[J].J Clin Micr obiol,1988,26(11):2465-2466.
[10]Soyer Y,Alcaine SD,Schoon maker-Bopp DJ,etal.Pulsedfield gel electr ophoresis diversity of hu man and bovine clinical Sal monella isolates[J].Foodbor ne Pat hog Dis,2010,7(6):707-717.DOI:10.1089/f pd.2009.0424
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:12
現代畜牧科技(2021年3期)2021-07-21 08:42:02
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
電子競技(2019年22期)2019-03-07 05:17:26
電子競技(2019年21期)2019-02-24 06:55:52
電子競技(2019年20期)2019-02-24 06:55:35
電子競技(2019年19期)2019-01-16 05:36:09
電子制作(2018年18期)2018-11-14 01:48:24
首都公共衛生(2017年5期)2018-01-03 08:15:39
