急性內毒素損傷對奶山羊肝臟營養代謝的影響
2012-09-20 00:32:28王林楓趙志偉楊改青王月影朱河水韓立強楊國宇
動物營養學報 2012年12期
王林楓 趙志偉 楊改青 王月影 朱河水 韓立強 張 震 楊國宇*
(1.河南農業大學農業部動物生化與營養重點實驗室,鄭州 450002;2.平頂山市畜牧局,平頂山 467000;3.河南農業大學生命研究中心,鄭州 450002)
內毒素(lipopolysaccharide,LPS)是革蘭氏陰性細菌(G-)細胞壁分解后釋放出的含脂多糖類物質[1],可導致機體的熱源性反應。正常情況下,機體內少量LPS隨門靜脈血進入肝臟,與肝臟枯否細胞上的Toll樣受體4(TLR4)結合,啟動腫瘤壞死因子 -α(TNF-α)、白細胞介素 -1β(IL-1β)等促炎癥因子的轉錄,進一步引起炎癥反應。少量的LPS刺激可以提高機體的免疫能力,當過量LPS進入肝臟則會引起強烈的炎癥反應,影響肝臟營養代謝并降低動物的免疫能力,嚴重的還會發生內毒素血癥[2]。研究報道,當動物發生內毒素血癥時,肝臟[3]、腎臟[4]、肺臟[5]等內臟器官受到嚴重的損傷。由此可見,肝臟是最易受LPS傷害的主要靶器官之一,但過量的LPS進入肝臟后對肝臟營養代謝的影響還未見報道。本試驗以奶山羊為研究對象,通過研究LPS所致急性肝臟損傷對肝臟營養物質代謝的影響,探索LPS對肝臟營養代謝的作用機制,為在生產中消除或減少LPS對動物健康和生產性能的影響提供科學依據。
1 材料與方法
1.1 試驗動物與分組
選用健康、體況相近、年齡2.5~3.0歲、體重35~40 kg的關中奶山羊母羊18只,隨機分為3組,每組6只,分別作為對照組(CTL組)、試驗Ⅰ組(TⅠ組)和試驗Ⅱ組(TⅡ組)。
1.2 飼養管理及飼糧組成
試驗羊飼喂于代謝籠(專利號:2009203148043)內,單籠飼養,飼糧和飲水分別從料槽和水槽供給,定量飼喂,自由飲水。根據NRC(2007)[6]山羊飼養標準設計飼糧配方(表 1),飼糧精粗料比為30∶70(干物質基礎),干物質中粗蛋白質含量為13.21%。飼喂量按山羊體重的3%供給,預試期14 d。
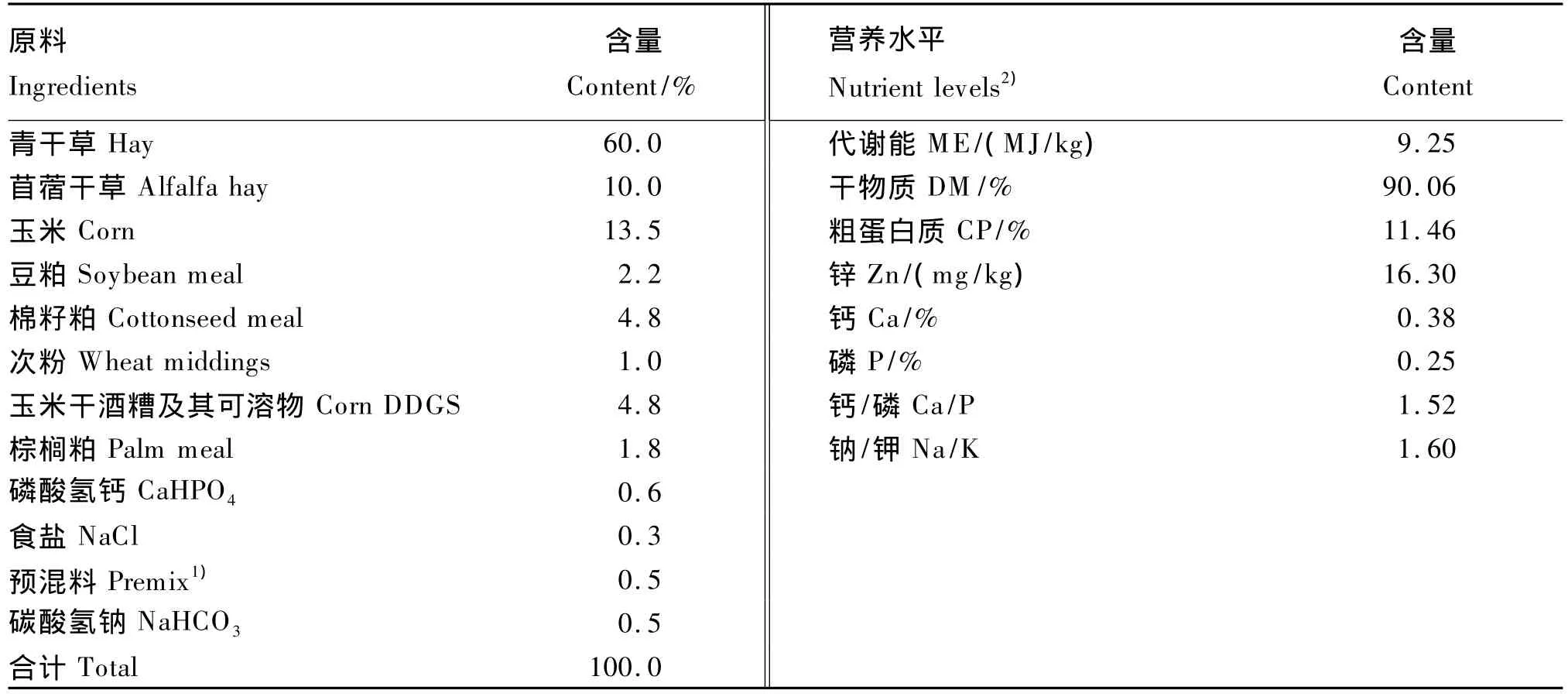
表1 飼糧組成及營養水平(風干基礎)Table1 Composition and nutrient levels of the diet(air-dry basis)
1.3 試驗處理
試驗前對所有試驗羊稱重,并依據體重調整LPS注射液的濃度[試驗前預先把LPS配制成0.5 mg/mL的溶液,LPS購自Sigma公司,其組成為大腸桿菌(E.coli)055∶B5]。試驗開始時,TⅠ組、TⅡ組奶山羊分別按照100和200 μg/kg BW的劑量從頸靜脈注射不同濃度的LPS注射液30 mL,CTL組奶山羊注射相同體積的生理鹽水,詳細方法及劑量參考Dow等[7]。
1.4 血樣采集與處理
分別在注射 LPS 后 0、1、4、8、12、24 h 從奶山羊的頸靜脈采血,每次采血時用真空抗凝采血管采血5 mL,立即以3 000 r/min離心15 min(離心機型號:TDZS-WS,湘儀離心機廠),分離血漿,置于EP管中,于-20℃保存待測。
1.5 指標測定及方法
肝臟代謝功能指標包括谷丙轉氨酶(ALT)、谷草轉氨酶(AST),營養代謝指標包括葡萄糖(GLU)、總蛋白(TP)、白蛋白(ALB)、甘油三酯(TG)、低密度脂蛋白(LDL)、高密度脂蛋白(HVDL)、總膽固醇(CHOL)、尿素氮(UN)。上述指標均采用全自動生化分析儀(Olympus AU640)檢測,ALT、AST測定試劑盒購自上海長征復星生物科技有限公司,其他指標測定試劑盒購自山東奧期幫生物科技有限公司。
1.6 數據統計分析
試驗數據用Excel 2007進行初步整理,應用SPSS 18.0軟件進行統計學分析。用協方差分析校正初始值,用單因素方差分析(one-way ANOVA)進行組間比較,以Duncan氏法進行多重比較,結果均以“平均值±標準差”表示。方差分析中,以P<0.05為差異具有顯著性意義。
2 結果與分析
2.1 LPS對奶山羊肝臟代謝功能的影響
從表2可以看出,血漿ALT活性平均值表現為TⅠ組>CTL組>TⅡ組,且 TⅠ組顯著高于CTL組(P<0.05),CTL組顯著高于TⅡ組(P<0.05)。從各時間點血漿ALT活性的變化趨勢來看,在各時間點均表現為TⅠ組>CTL組>TⅡ組,在6 h前的各時間點3組之間差異不顯著(P>0.05);12 h時,TⅠ組明顯升高,TⅡ組和CTL組基本穩定,但各組之間仍差異不顯著(P>0.05);24 h時,TⅠ組顯著高于TⅡ組和CTL組(P<0.05),TⅡ組和CTL組之間差異不顯著(P>0.05)。
與血漿ALT活性不同,血漿AST活性平均值表現為TⅠ組>TⅡ組>CTL組,TⅠ組顯著高于TⅡ組和CTL組(P<0.05),但TⅡ組和 CTL組之間差異不顯著(P>0.05)。從各時間點血漿ALT活性的變化趨勢來看,1 h后TⅠ組就明顯升高,并始終顯著高于TⅡ組和CTL組(P<0.05),而TⅡ組和CTL組在各時間點均差異不顯著(P>0.05)。
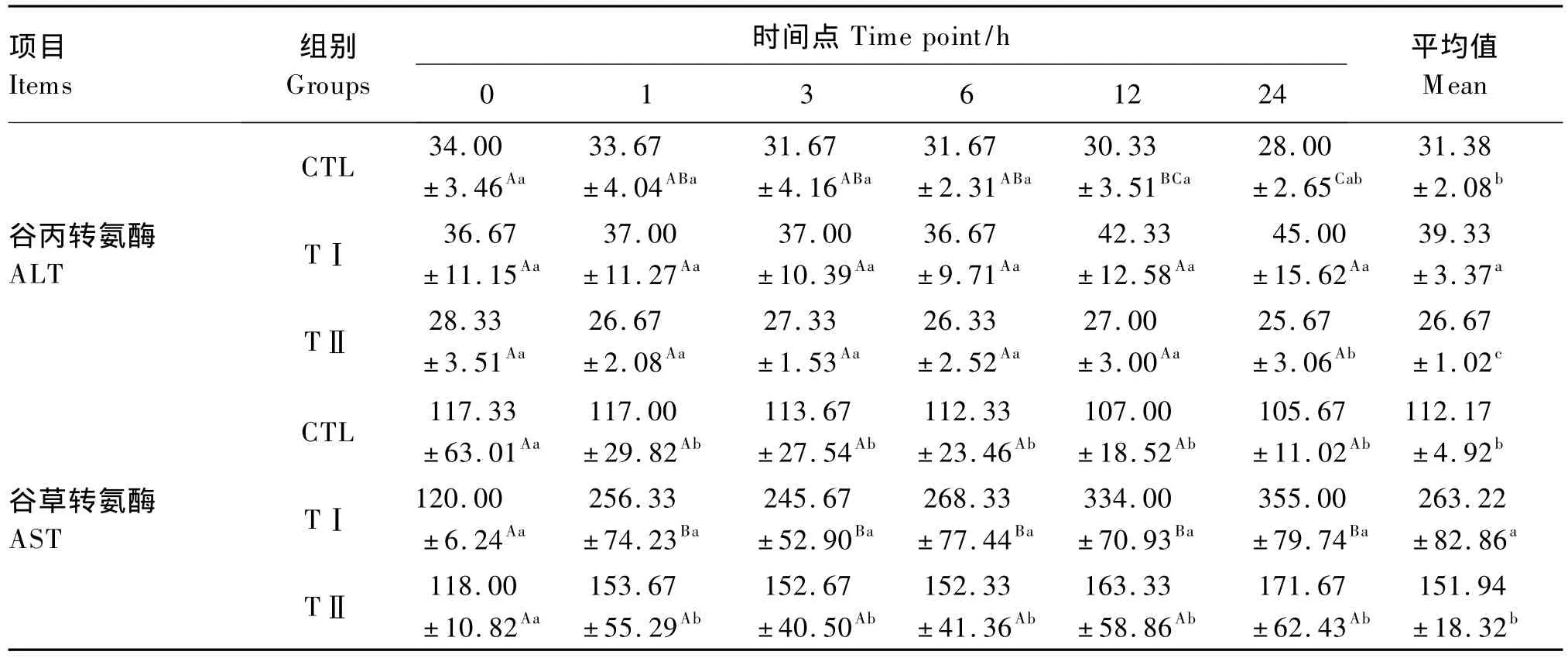
表2 各組奶山羊血漿ALT、AST活性Table2 Plasma ALT and AST activities of dairy goats in different groups U/L
2.2 LPS對奶山羊肝臟糖代謝的影響
從表3可以看出,血漿GLU含量平均值表現為CTL組>TⅡ組>TⅠ組,但各組之間均差異不顯著(P>0.05)。隨著時間的推移,CTL組血漿GLU含量基本穩定;TⅠ組和TⅡ組血漿GLU含量有較大的變化,表現為先升高,隨后迅速降低,6 h時2組都降至最低,以后緩慢回升,TⅠ組回升的速度高于TⅡ組,但2組之間差異不顯著(P>0.05)。
2.3 LPS對奶山羊肝臟蛋白質代謝的影響
由表4可以看出,血漿TP含量平均值表現為CTL組>TⅡ組>TⅠ組,TⅠ組和TⅡ組顯著低于CTL組(P<0.05),而TⅠ組和TⅡ組之間差異不顯著(P>0.05)。從各時間點血漿TP含量的變化趨勢來看,CTL組基本穩定,各時間點變化不大;TⅠ組在6 h前持續降低,各時間點差異顯著(P<0.05),6 h后稍有回升,各時間點差異不顯著(P>0.05);TⅡ組表現為先降低,后穩定,6 h時最低,顯著低于0和1 h時(P<0.05),6 h后基本穩定,各時間點差異不顯著(P>0.05)。
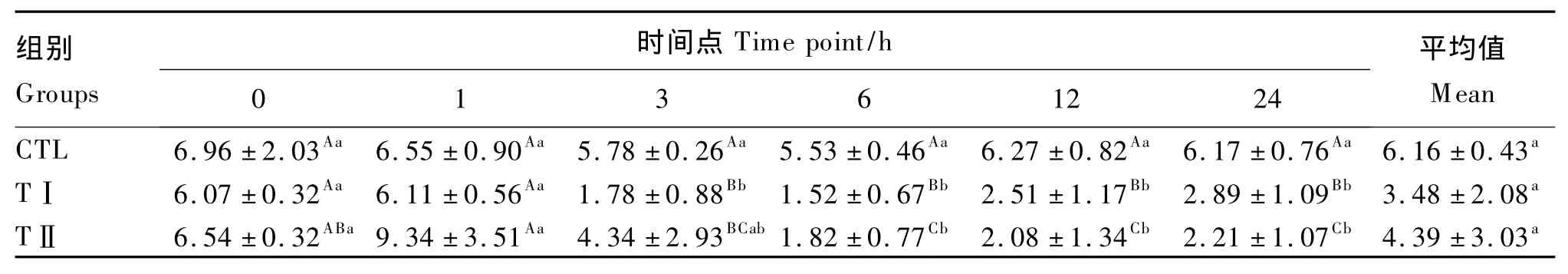
表3 各組奶山羊血漿葡萄糖含量Table3 Plasma glucose content of dairy goats in different groups mmol/L
血漿ALB含量平均值表現為CTL組>TⅠ組>TⅡ組,但各組之間均差異不顯著(P>0.05)。隨著時間的推移,CTL組血漿ALB含量基本穩定;TⅠ組和TⅡ組血漿ALB含量逐漸降低,TⅡ組降低的幅度大于TⅠ組,但2組之間差異不顯著(P>0.05)。
血漿UN含量平均值表現為TⅡ組>TⅠ組>CTL組,CTL組顯著低于 TⅠ組和 TⅡ組(P<0.05),但TⅠ組和TⅡ組之間差異不顯著(P>0.05)。隨著時間的推移,CTL組血漿UN含量基本穩定;TⅠ組和TⅡ組血漿LDL含量逐漸升高,1 h后顯著高于CTL組(P<0.05),TⅡ組升高的幅度大于TⅠ組,但除在3和12 h時差異顯著(P<0.05)外,其他時間點均差異不顯著(P>0.05)。

表4 各組奶山羊血漿蛋白質類及尿素氮含量Table4 Plasma protein and urea nitrogen contents of dairy goats in different groups
2.4 LPS對奶山羊肝臟脂肪代謝的影響
由表5可以看出,血漿TG含量平均值表現為TⅠ組>TⅡ組>CTL組,TⅠ組和TⅡ組顯著高于CTL組(P<0.05),而TⅠ組和TⅡ組之間差異不顯著(P>0.05)。從各時間點血漿TG含量的變化趨勢來看,從1 h開始的各時間點TⅠ組、TⅡ組均顯著地高于CTL組(P<0.05),在各時間點TⅠ組均高于TⅡ組,且在3和6 h時達到顯著水平(P<0.05)。
血漿LDL含量平均值表現為TⅠ組>TⅡ組>CTL組,TⅠ組顯著高于TⅡ組和 CTL組(P<0.05),而TⅡ組和CTL組之間差異不顯著(P>0.05)。隨著時間的推移,TⅡ組血漿LDL含量基本呈下降趨勢,在個別時間點間達到顯著差異(P<0.05);TⅠ組和CTL組血漿LDL含量在各時間點變化不顯著(P>0.05)。
血漿HDL含量平均值表現為TⅠ組>TⅡ組>CTL組,TⅠ組顯著高于TⅡ組和 CTL組(P<0.05),TⅡ組顯著高于 CTL組(P<0.05)。隨著時間的推移,TⅠ組血漿HDL含量表現為緩慢升高的趨勢,除個別時間點外,大多數時間點差異不顯著(P>0.05);TⅡ組和CTL組血漿HDL含量在各時間點變化不顯著(P>0.05)。
血漿CHOL含量平均值表現為TⅠ組>TⅡ組>CTL組,TⅠ組顯著高于TⅡ組和 CTL組(P<0.05),TⅡ組顯著高于CTL組(P<0.05)。隨著時間的推移,CTL組血漿CHOL含量在各時間點變化不顯著(P>0.05);TⅠ組血漿CHOL含量表現為先降低后升高的趨勢,6 h后開始升高,24 h時顯著高于其他各時間點時(P<0.05);TⅡ組血漿CHOL含量表現為先升高后降低再升高的趨勢,6和12 h時較低,顯著低于1 h時(P<0.05),其他各時間點之間差異不顯著(P>0.05)。
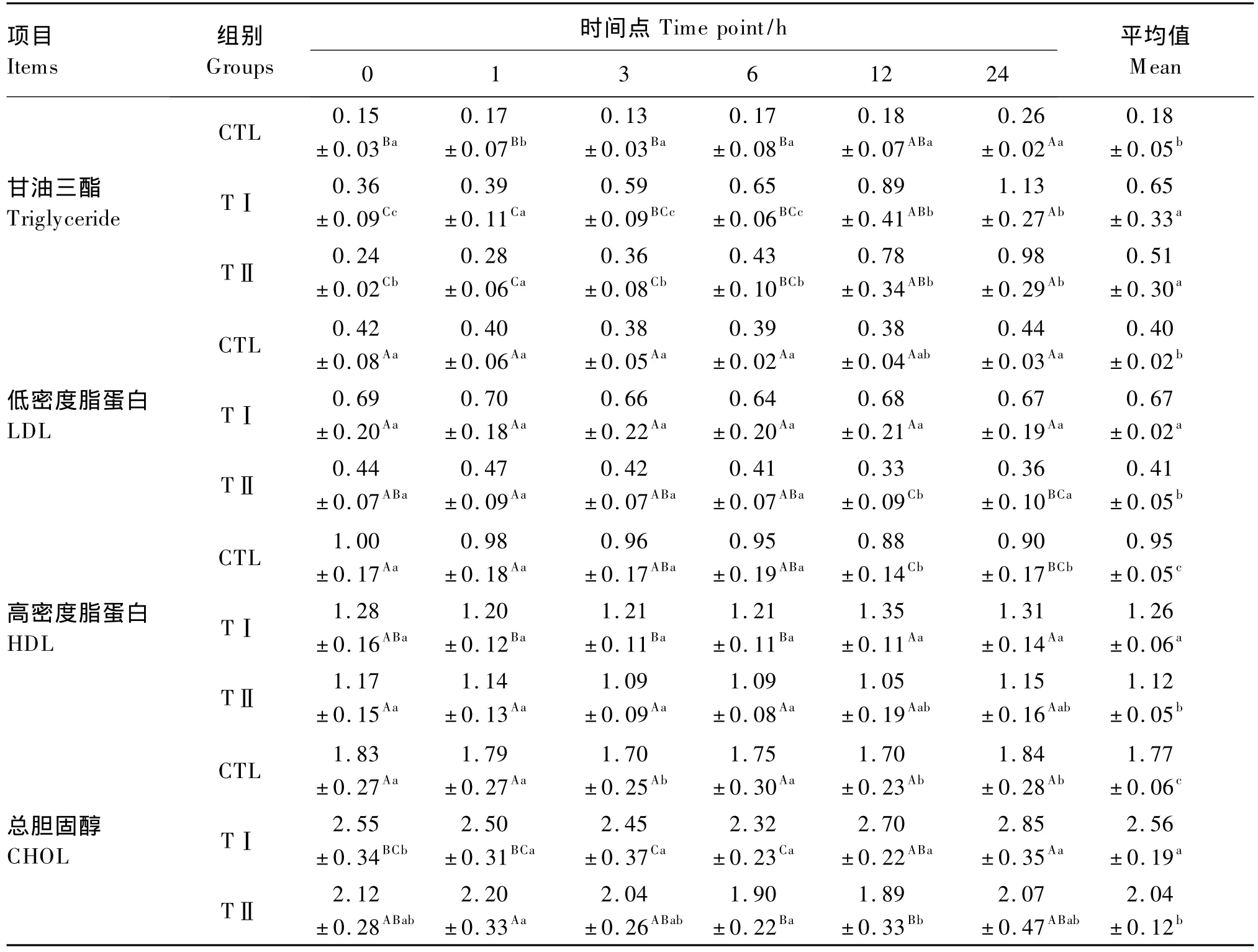
表5 各組奶山羊血漿中脂類的含量Table5 Plasma lipid contents of dairy goats in different groups mmol/L
3 討論
3.1 LPS對奶山羊肝臟代謝功能的影響
ALT是肝細胞中主要的代謝酶,是可逆地催化丙酮酸和谷氨酸之間氨基酸轉移的酶,以磷酸吡哆醛作為輔助因子。ALT主要存在于肝細胞漿,很少外泄,胞內濃度是血漿濃度的1 000~3 000倍,只有肝細胞受到損傷或壞死,才有大量ALT進入血液,引起血漿ALT活性升高,肝細胞壞死增加1%就可使血漿中ALT活性升高1倍。因此,ALT是反映肝功能損傷程度最敏感的指標之一。本試驗中,TⅠ組血漿ALT活性顯著高于CTL組,表明100 μg/kg BW 劑量的 LPS對肝細胞已造成明顯損傷;TⅡ組ALT活性低于CTL組,表明200 μg/kg BW劑量的LPS對肝臟已不具有損傷,證明LPS對肝臟的損傷與劑量有關,高劑量的LPS可減輕對肝臟的傷害,至于多高劑量時開始減輕,目前還未見研究報道。關于高劑量LPS對肝臟損傷降低的原因,目前的解釋是LPS對細胞的作用需通過LPS結合蛋白(LBP)介導與CD14(一種細胞因子,參與LPS的受體組成)形成復合物后才能夠與TLR4結合,激活下游信號通路,產生炎癥因子,損傷肝細胞。已研究表明,LBP的增敏效應與劑量有關,高劑量的LPS對LBP的增敏效應有抑制作用,抑制了LPS與CD14的結合,從而阻止了LPS所引起的炎癥反應,減輕了其對肝細胞的傷害[8]。此外,殺菌通透性增加蛋白(BPI)和HDL均可抑制LPS與受體的結合,降低其對肝細胞的損傷[9-10]。
AST是一種具有磷酸吡哆醛依賴性、由細胞核基因編碼的線粒體酶,催化天冬氨酸的氨基轉移到α-酮戊二酸形成草酰乙酸和谷氨酸及其逆反應。肝臟內的谷草轉氨酶有2種同工酶,分別存在于肝細胞的胞漿內(sAST)和線粒體內(mAST)。當肝細胞受損或任何原因引起肝細胞膜通透性增加時,位于肝細胞內的AST逸出細胞進入血液,使血漿AST活性增高[11]。在肝細胞輕度病變時,僅 sAST釋放入血;當病變嚴重時,mAST也會相繼釋放入血,故AST活性的高低可反映肝細胞的損傷程度和壞死量[12]。另有研究表明,mAST易于被網狀免疫系統清除,其清除率比sAST快5倍。若肝細胞急性損傷得以緩解,則血漿中mAST很快下降,甚至恢復到正常水平[13]。因此,mAST不僅作為判斷細胞壞死的指標,也是對肝臟疾病動態監測的指標[14]。本試驗中,TⅠ組、TⅡ組奶山羊血漿AST活性高于CTL組,表明TⅠ組、TⅡ組奶山羊的肝臟在LPS的作用下受到損傷;而TⅡ組AST活性低于TⅠ組,表明TⅡ組奶山羊的肝臟損傷有所緩和,證明LPS對肝臟的傷害與劑量有關。
3.2 LPS對肝臟糖代謝的影響
肝臟是反芻動物糖異生的重要器官,大量LPS進入機體以后,會造成急性肝功能衰竭內毒素血癥,同時刺激機體產生多種內源性介質和細胞因子,肝臟糖異生受到抑制,導致血糖含量降低[15]。但由于應激,在注射LPS后的短時間內血糖含量表現為升高,之后,隨著時間的延長,表現為低血糖癥狀。此外面,注射LPS后,由于LPS的毒性作用導致細胞缺血缺氧,細胞有氧代謝減弱,無氧酵解增強,對葡萄糖的需求增加,糖異生增強,使血糖含量在短時間內升高。隨著時間的推移,由于糖異生受到抑制,最終導致血糖含量降低[16]。本試驗中,注射LPS后血糖(血漿葡萄糖)含量表現出先升高后降低的趨勢,證明了上述結論。LPS抑制肝臟糖異生的分子機制有待進一步研究。
3.3 LPS對肝臟蛋白質代謝的影響
肝臟在蛋白質代謝過程中起重要作用,血漿內蛋白質幾乎全部由肝臟合成,TP主要包括ALB和球蛋白(GBL),其中ALB占到40% ~60%。血漿ALB是機體各組織合成自身蛋白質的原料,是許多脂溶性物質的非特異性運輸載體,在維持血漿膠體滲透壓方面也起著重要作用,因此,血漿TP、ALB含量是反映肝臟健康狀況的重要指標。本試驗中,TⅠ組和TⅡ組TP、ALB含量不同程度地降低,表明肝臟受到損傷后合成蛋白質的能力減弱。
本試驗中,TⅠ組和TⅡ組的血漿UN含量升高,表明肝臟內蛋白質的分解增加,產生大量的尿素,在LPS中毒的情況下,奶山羊的采食量和消化活動受到很大影響,尿素循環和再利用減少,使血漿UN含量升高。腎臟是尿素的主要排泄器官。肝損傷后,腎臟的功能受到影響,氮的排泄受到影響。研究發現,LPS誘導后小鼠血清尿素的水平較誘導前顯著升高[17]。LPS引起腎功能損傷后,導致尿素排泄障礙,是血漿UN含量升高的另外一個重要原因[18]。本試驗中,奶山羊血漿UN含量隨著劑量的加大而升高,表明LPS對奶山羊腎臟功能的損傷與劑量呈正相關。
3.4 LPS對脂肪代謝的影響
研究表明,LPS促進脂肪酸氧化,使脂肪分解增強、血脂增高,從而引起肝臟脂代謝紊亂[19-20]。LPS及其他的炎癥因子,如TNF-α、白細胞介素-1(IL-1)、白細胞介素-2(IL-2)等,通過刺激肝臟中富含TG的VLDL的釋放而使血漿TG含量升高[21];LPS通過降低脂蛋白酯酶(LPL)活性抑制其對TG的清除,炎癥也能減少血漿富含TG的脂蛋白的清除[22]。由于肝臟損傷,一方面,VLDL的合成和分泌減少,使血液中游離脂肪酸(FFA)的含量增多,導致大量的TG在肝臟內沉積;另一方面,肝臟內發生大量的氧化,肝臟損傷后其抗氧化能力減弱,大量的自由基在肝臟中蓄積,進一步加重了肝臟的損傷[22]。本試驗中,TⅠ組、TⅡ組血漿TG含量均高于CTL組,并隨時間延長逐漸升高,與上述研究結果相同;且血漿TG含量TⅡ組較TⅠ組有所下降,表明LPS對TG的升高作用與劑量有關,高劑量的LPS對TG的升高作用反而降低。
HDL和CHOL是肝臟脂肪代謝過程中的重要組成物質。膜轉運蛋白三磷酸腺苷結合盒轉運體(ABCA1)可將游離膽固醇從胞內流向胞外,與載脂蛋白AⅠ(ApoAⅠ)等結合形成HDL,逆向運回肝臟,在清除外周血膽固醇的過程中發揮作用。研究表明,LPS能干擾CHOL的逆轉運過程,通過抑制CHOL逆轉運相關蛋白干擾CHOL的逆轉運,造成細胞內 CHOL 積聚[22]。Ruan 等[23]研究證實,LPS可抑制ABCA1基因的表達,使細胞內CHOL含量升高;Castrillo等[24]證實,通過肝臟 X受體(LXR)和Toll樣受體(TLR)通路交互作用,LPS激活TLR4通路后抑制LXR通路,使巨噬細胞膜轉運蛋白ABCA1和載脂蛋白E(ApoE)的基因表達減少,減弱CHOL的逆轉運。研究表明,高脂負荷顯著下調卵磷脂膽固醇脂酰轉移酶(LCAT)mRNA的表達水平,導致血漿LCAT的活性降低,進而使HDL表面游離CHOL的酯化減少,降低了HDL對外周組織CHOL的清除能力,增加動脈粥樣硬化(AS)發生的可能性[25]。本試驗中,TⅠ組、TⅡ組CHOL含量升高可能是LPS干擾CHOL的逆向轉運,導致血液中CHOL無法正常運往肝臟而積聚,并導致HDL含量的上升。
HDL是將肝外組織的CHOL運送到肝臟的運載工具,可以防止游離CHOL在肝外組織細胞上的沉積。HDL主要在肝臟、小腸中合成,降解主要在肝臟中。HDL通過CHOL的逆向轉運,把外周組織中衰老細胞膜上以及血漿中的CHOL運回肝臟代謝,是脂代謝中的重要物質。據報道,各種脂蛋白,如 LDL、VLDL、HDL 和 VHDL,對 LPS 均有一定解毒作用,其中以HDL與LPS的結合能力最高,可有效地清除血液中的 LPS[8,26]。但不同劑量的LPS對其與HDL的親和力以及清除效率的影響,尚待進一步研究。
4 結論
①LPS可引起肝臟損傷,進而對奶山羊肝臟營養代謝造成影響。
②LPS對奶山羊肝臟營養代謝的影響與LPS的劑量有關。
[1]王磊,于麗萍,竇科峰.內毒素致枯否細胞中NF-κB的激活及其意義[J].寧夏醫學院報,2003,25(4):239-241.
[2]MORRISON D S,RYAN J C.Endotoxin and disease mechanism[J].Annual Review of Medicine,1987,38:417-432.
[3]畢銘華,張淑文,王寶恩,等.內毒素血癥對大鼠肝細胞線粒體的損傷及其機制[J].中國應用生理學雜志,2004,20(1):90-92.
[4]董宇杰,丁春華,張智,等.褪黑素對內毒素致大鼠急性肺損傷的保護作用[J].中國應用生理學雜志,2010,26(4):481-484.
[5]郭德玉,譙怡然.內毒素介導的腎臟病變及其發生機理的實驗研究[J].臨床與實驗病理學雜志,1995,11(3):226-229.
[6]NRC.Nutrientrequirementsofsmallruminants:sheep,goats,cervids and New World camelids[S].Washington,D.C.:National Academy Press,2007.
[7]DOW T L,ROGERS-NIEMAN G,HOLáSKOVá I,et al.Tumor necrosis factor-α and acute-phase proteins in early pregnant ewes after challenge with peptidoglycan-polysaccharide[J].Domestic Animal Endocrinology,2010,39:147-154.
[8]鄭瑞丹,徐成潤.脂多糖/脂多糖結合蛋白與殺菌通透性增進行蛋白研究進展[J].臨床軍醫雜志,2007.35(6):933-936.
[9]SCHRODER N W,SCHUMANN R R.Non-LPS targets and actions of LPS binding protein(LBP)[J].Journal of Endotoxin Research,2005,11(4):237-242.
[10]AZUMA M,MATSUO A,FUJIMOTO Y,et al.Inhibition of lipid A2 mediated typeⅠinterferon induction by bactericidal/permeability-increasing protein(BFI)[J].Biochemical Biophysical Research Community,2007,354(2):574-578.
[11]査錫良,周春燕,周愛儒,等.生物化學[M].北京:人民衛生出版社,2008:409.
[12]鄭曉豐.m-AST、RBP、PA、ADA和 FN在肝臟疾病診斷中的臨床評價[J].檢驗醫學,2009,24(12):886-888.
[13]康格非.臨床生物化學與生物化學檢驗[M].2版.北京:人民衛生出版社,1999:166.
[14]HASANJANI-ROUSHAN M R,HAJIAHMADI M,SHAFAIE S.Histopathological features of liver and its relation to serum transaminase levels in 91 cases of anti-HBe-positive chronic hepatitis B[J].The International Journal of Clinical Practice,2005,59(7):79l-794.
[15]朱永芬,田德英.內毒素對SD大鼠急性肝功能衰竭模型血糖代謝影響的研究[J].內科急危重癥雜志,2006,12:119-121.
[16]JANICE J,STEVE S C,ANNA M D.Mechanisms of disease progression in nonalcoholic fatty liver disease[J].Seminar in Liver Disease,2008,28(4):370-379.
[17]KENT D,ASADA L,PETER S T,et al.Animal models of sepsis and sepsis-induced kidney injury[J].The Journal of Clinical Investigation,2009,119(10):2868-2878.
[18]章蔚,方才,言虎.不同劑量6%羥乙基淀粉130/0.4預先給藥對大鼠內毒素性急性腎損傷的影響[J].安徽醫科大學學報,2007,42(6):672-675.
[19]李生,吳萬春.內毒素與脂肪肝的關系研究進展[J].國際消化病雜志,2007,27(3):205-207.
[20]YOSHIMATSU M,TERASAKI Y,SAKASHITA N,et a1.Induction of macrophage scavenger receptor M ARCO in nonalcoholic steato-hepatitis indicates possible involvementofendotoxin in its path-ogenic process[J].International Journal of Experimental Pathology,2004,85:335-343.
[21]FABBRINI E,SULLIVAN S,KLEIN S.Obesity and nonalcoholic fatty liver disease:biochemical,metabolic,and clinical implications[J].Hepatology,2010,51:679-689.
[22]葉強,雷寒.內毒素、炎癥與脂質代謝紊亂的研究進展[J].心血管病學進展,2008,29(5):782-784.
[23]RUAN X Z,MOORHEAD J F,FERNADO R.Regulation of lipoprotein trafficking in the kidney role of inflammatory mediators and transcription factors[J].Biochemical Society Transactions,2004,32:88-91.
[24]CASTRILLO A,JOSEPH SB,VAIDYA SA.Crosstalk between LXR and toll-like receptor signaling mediates bacterial and viral antagonism of cholesterol metabolism[J].Molecular Cell,2003,12:805-816.
[25]張娜,趙斐,張勇.有氧運動改善高脂血癥分子機理的研究Ⅲ.運動上調飲食性高膽固醇血癥大鼠肝臟LCAT和apoAI基因表達[J].中國運動醫學雜志,2001,20(3):232-235.
[26]LEVELS J H,AHRAHAM P R,VANDEN E A,et al.Distribution and kinetics of lipoprotein-bound endotoxin[J].Infection and Immunity,2001,69(5):2821-2828.
猜你喜歡
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
現代臨床醫學(2022年4期)2022-09-29 07:38:00
音樂探索(2022年2期)2022-05-30 21:01:37
昆明醫科大學學報(2021年4期)2021-07-23 01:21:50
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
海南醫學(2016年8期)2016-06-08 05:43:00
醫學研究雜志(2015年9期)2015-07-01 17:28:15
