寧夏4種灌木光合固碳能力的比較
2012-09-18 06:32:52李志剛朱強李健
草業科學 2012年3期
李志剛,朱強,李健
(種苗生物工程國家重點實驗室寧夏林業研究所,寧夏銀川750004)
寧夏4種灌木光合固碳能力的比較
李志剛,朱強,李健
(種苗生物工程國家重點實驗室寧夏林業研究所,寧夏銀川750004)
通過對葉片光合速率及植株生物量的測定,研究了寧夏4種常見灌木的固碳潛力。結果表明,從4種灌木單個葉片的光合角度研究固碳能力,生長旺盛季(7月)表現為互葉醉魚草(Buddlejaalternifolia)>蒙古扁桃(Amygdalusmongolica)>沙木蓼(Atraphaxisbracteata)>寧夏枸杞(Lyciumbarbarum);全生長季為寧夏枸杞>互葉醉魚草>沙木蓼>蒙古扁桃。經過對4種灌木的單位葉面積和全株葉面積的測定,研究了全株灌木的固碳潛力,結果顯示,不論是7月的全株日均碳同化量還是全生長季的全株日均碳同化量,都表現出互葉醉魚草>寧夏枸杞>沙木蓼>蒙古扁桃的規律,與生物量測定的結果完全吻合。說明,從植物光合角度研究固碳能力有必要從全株葉片水平的光合進行考慮。另外,由于寧夏枸杞經濟林具有良好的水肥管理措施,推測經濟林有較強的固碳能力,應予以重視。
寧夏;灌木;固碳;光合
隨著人們對全球氣候變化的認識,植物的固碳功能和碳匯造林已備受學者關注。目前,植物固碳現狀和潛力的研究多集中于基于生物量或林木蓄積量的測定而間接測定出陸地生態系統或植物群落的碳儲量,其次是采用渦度相關法或渦度協方差法直接測定生態系統CO2的通量[1-7],對于不同植物種之間固碳能力的研究也多采用生物量比較法[8-9],而植物光合作用是生態系統物質循環和能量流動的基礎,也是植物固碳的最主要的方式,反映了植物改善環境的能力[10]。因此,也有采用測定植物光合作用估測植物固碳能力的研究,但是常常集中于單位葉面積葉片的光合固碳效率研究[11],涉及單株葉面積比較的研究較少,這就可能掩蓋了研究結果的真實性。隨著對植被恢復等生態工程的重要性的認識,高效固碳釋氧生態樹種在植被恢復建設中應該予以考慮,尤其是在特定生態區下篩選既適宜當地自然生態條件又兼具較好生態效益的樹種就顯得更加有實際意義。因此,準確評價不同植物之間的固碳能力也就顯得十分必要。本研究基于以上目的,選擇寧夏常見的4種灌木林比較其固碳能力,旨在為寧夏干旱半干旱地區及其他類似生態區的植被恢復、碳匯造林及經濟建設提供理論支持。
1 材料與方法
1.1 研究區概況 研究區設在銀川植物園,地處寧夏回族自治區銀川市西南,106°10′E,38°25′N,海拔1 166m。位于我國半荒漠地區,即草原向荒漠的過渡地帶,賀蘭山東麓洪積扇下緣沙地,有流動沙丘分布。日照充足、熱量充沛、溫差較大、風大沙多、干旱少雨、蒸發強烈。年平均氣溫8.5℃。日照時數2 710h,年降水量286mm[12]。
1.2 研究方法
1.2.1 研究對象 所研究的4種灌木互葉醉魚草(Buddlejaalternifolia)、蒙古扁桃(Amygdalusmongolica)、沙木蓼(Atraphaxisbracteata)和寧夏枸杞(Lyciumbarbarum)均為寧夏原生境下常見的灌木植物,其中前3種2007年從寧夏賀蘭山引種并在銀川植物園建立種質資源圃(栽培株距為1m),占地面積均為500m2左右,每年通過滴灌系統澆水120~200 m3·667m-2(根據每年降水量不同有所調整);寧夏枸杞(寧杞1號)亦于2007年在該植物園建立栽培試驗地(株行距為1m×1m),面積2hm2,每年通過滴灌系統澆水230~259m3·667m-2。以上4種灌木灌木林每年均有施肥處理,其土壤條件如表1所示。
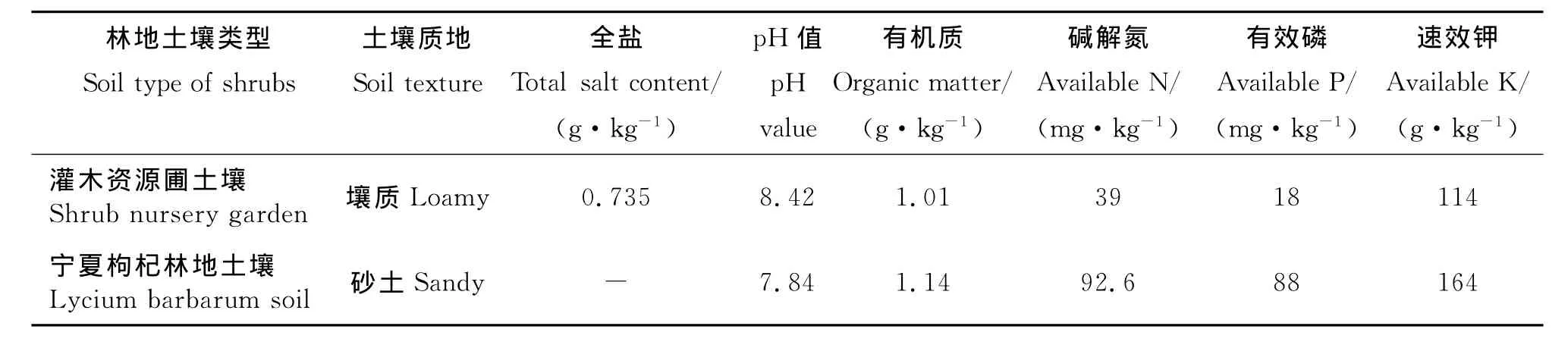
表1 不同灌木林地0~40cm土壤特征Table 1 Characteristics of soil at 0-40cm of different shrub lands
1.2.2 測定方法
同化速率的測定:首先,2010年根據天氣狀況在灌木生長季節(5―10月)的每月中旬選擇無風、晴朗的天氣進行光合潛力的季節動態測定,采用CI-340便攜式光合儀依次在一天中的09:00、13:00和17:00分別測定4種灌木光合速率;同時,在灌木生長最旺盛的季節(7月)選擇晴朗、無風的天氣進行光合潛力的日動態測定(仍選中旬),從08:00-18:00每隔2h測定一次,完成所有灌木的測定。以上測定時均從每個灌木冠層中部位置的東南西北4個方向各選擇一片葉片進行測定。
灌木同化率的估算:7月日碳同化總量是凈光合速率曲線與時間橫軸圍合的面積,利用光合速率對時間的積分求得,日均碳同化率(即平均光合速率)為碳同化總量值與時間的比值。生長季灌木每月的日平均凈光合速率為每月中旬所測得的3個時間點凈光合速率的平均值,平均碳同化率估算方法同7月日均碳同化率。
灌木葉面積及生物量的測量:灌木葉面積的測定在6、8和10月進行,選擇陰天或早晨溫度較低時,在試驗地每種灌木選擇能夠代表群體長勢特征的3株植株,將其葉片全部摘下帶回室內進行測定。再從每株灌木葉片中選擇能夠代表全株群體長勢的葉片20片,利用Li-3000C葉面積儀測定葉面積,同時稱其鮮質量,然后根據已知面積單葉的鮮質量換算出全株的葉面積。在10月完成最后一次單葉面積和全株葉面積的測定工作后,將全株灌木連根盡可能完全挖出,取樣深度為1m,此深度為灌木根系分布的最深深度。在挖取根系時沿主根和須根延伸方向盡可能取盡所有根系。取樣完畢后將灌木的葉片、枝條和根系分開并置于85℃的烘箱中烘至質量恒定,完成生物量的測定。
1.2.3 數據分析 采用Excel 2003建立數據庫并繪圖,數據的顯著性統計分析采用SPSS 13.0軟件。
2 結果與分析
2.1 4種灌木的單位葉面積光合速率 光合速率是反映植物生長潛力和碳同化能力的一個重要指標。在本研究中,生長旺盛季節(7月)4種灌木的單位葉面積光合速率日動態有所不同(圖1A),其中沙木蓼和寧夏枸杞呈單峰曲線,而互葉醉魚草和蒙古扁桃呈現出雙峰曲線,但不易看出4種植物的光合速率大小,而從日均光合速率值可以看出(圖2),其大小順序為互葉醉魚草>蒙古扁桃>沙木蓼>寧夏枸杞,其中蒙古扁桃與沙木蓼間差異不顯著(P>0.05)。全生長季日均光合速率又不同于生長旺盛季(圖1,圖2),從研究結果看,灌木間在全生長季的日均光合速率呈現出寧夏枸杞>互葉醉魚草>沙木蓼>蒙古扁桃的規律,其中前二者差異不顯著,而且從日均光合速率的季節動態亦可以得出類似的結果(圖1B),此結果完全不同于灌木間在生長旺盛季的日均光合速率規律,由此看來,單憑某一日或某一月的單位葉面積光合速率(或日同化量)并不能正確反映植物的單位葉面積碳同化能力,須從整個生長季進行全面研究。另外,從單位葉面積光合速率的季節動態結果可以看出(圖1B),4種灌木的單位葉面積碳同化能力均在7月最大,5月或10月最小,在生長季均呈單峰曲線變化。
2.2 4種灌木葉面積與單株的碳同化潛力
理論上講,植物全株葉片在全生長季的凈光合速率值即全株年碳同化量才是真正意義上反映植物碳同化現狀的值。將6、8和10月所測的得灌木單葉面積和全株葉面積求平均值,可以看出,這2個表征植物碳同化能力的葉面積指標在4種灌木間存在明顯不同(圖3),雖然平均單葉面積表現為沙木蓼>寧夏枸杞>互葉醉魚草>蒙古扁桃,四者差異顯著(P<0.05),但是平均全株葉面積卻表現為互葉醉魚草>寧夏枸杞>沙木蓼>蒙古扁桃,且四者差異顯著。在此基礎上,本研究將平均光合速率和平均全株葉面積相乘得到全株的碳月均同化量(圖4),結果表明,不論是7月的日均碳同化量還是全生長季的日均碳同化量,都表現為互葉醉魚草>寧夏枸杞>沙木蓼>蒙古扁桃,差異顯著。說明研究某一植物的碳同化量必須從全株植株出發。
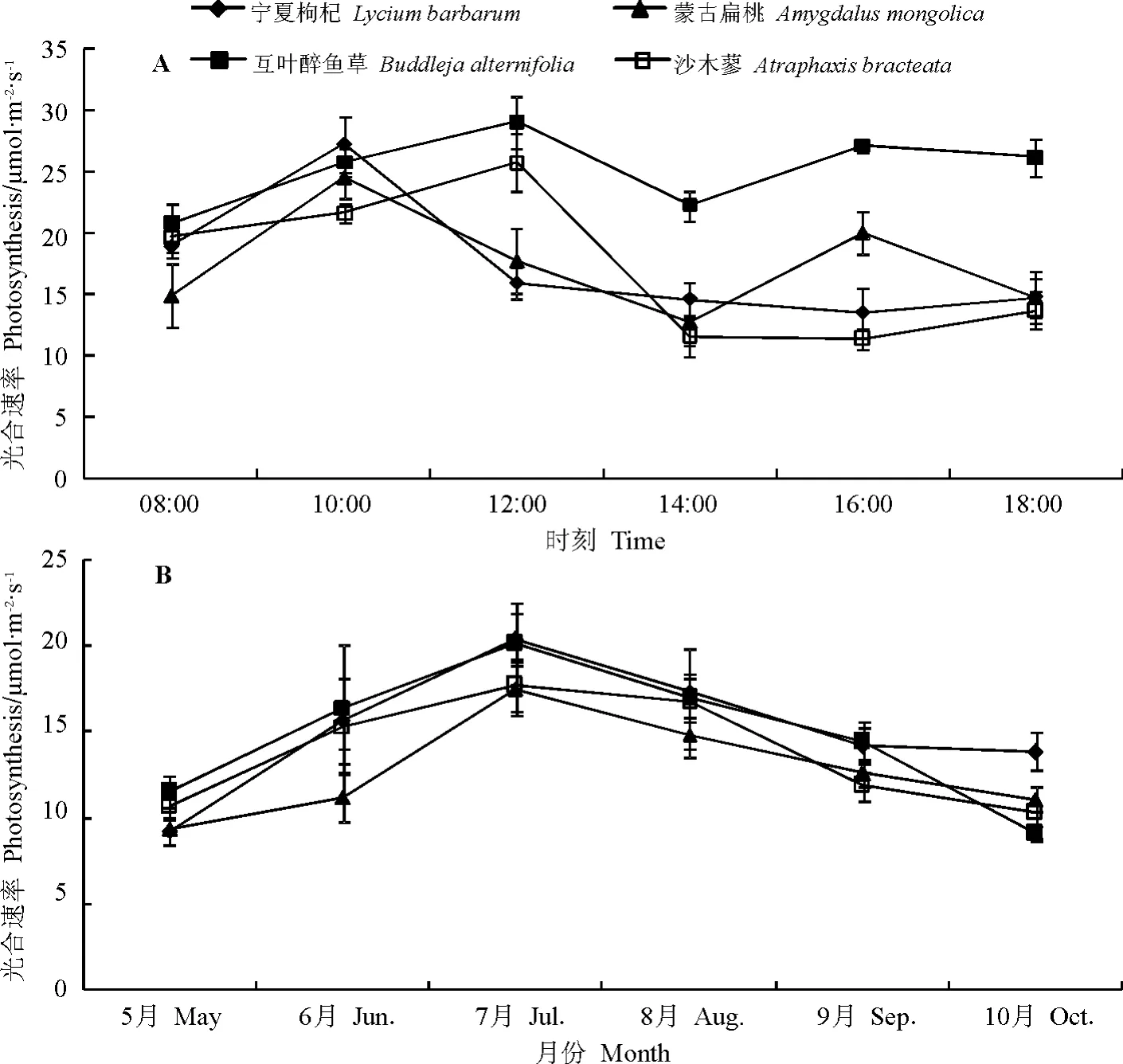
圖1 4種灌木的單位葉面積光合日動態(A)與季節動態(B)Fig.1 Daily and seasonal dynamic of photosynthesis per unit leaf area of the 4shrub species
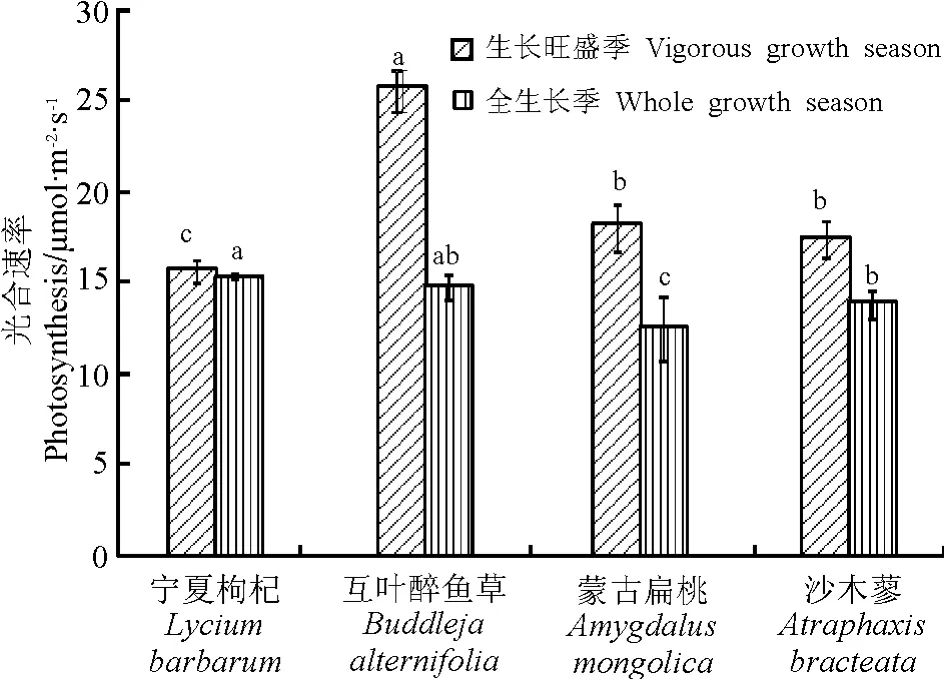
圖2 4種灌木的平均單位葉面積光合速率Fig.2 Average photosynthetic rate per unit leaf area of the 4shrub species
2.3 4種灌木的生物量 植物生物量是反映植物碳匯大小的一個重要指標。在本研究中,4種灌木單位葉面積光合速率、單葉面積及全株日均碳同化量等指標間的表現規律并不一致,但是根據全株葉面積所計算出的全株月均碳同化量是可以反映灌木固碳能力的一個最終指標。然而對生物量的研究結果表明(表2),4種灌木生物量的大小順序為互葉醉魚草>沙木蓼>蒙古扁桃>寧夏枸杞,與全株的日均碳同化量又存在不一致的情況,出現這種情況的原因是沒有將寧夏枸杞每年修剪掉的枝條及采收的果實計算在內(表3),如果將這些生物量(1 114.76g)計算在內,則4年生寧夏枸杞的生物量累計可達到1 790.10g。此時,4種灌木的生物量大小順序為互葉醉魚草>寧夏枸杞>沙木蓼>蒙古扁桃,完全和全株碳平均同化量一致。說明深入研究植物的固碳能力必須從研究全株碳同化量出發。
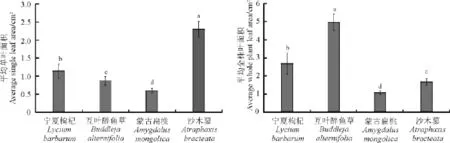
圖3 4種灌木的單葉面積與全株葉面積Fig.3 Leaf area of single and whole plant of the 4shrub species
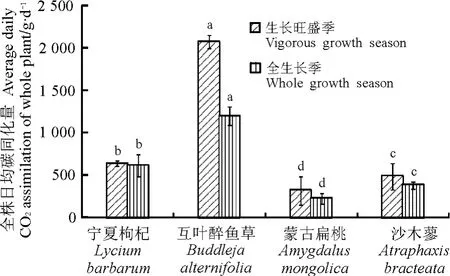
圖4 4種灌木全株日均同化量Fig.4 Carbon sequestration of whole plant of the 4shrub species

表2 不同灌木的生物量Table 2 Biomass of different shrubs g
注:生物量以2010年統計的干物質計算;不同小寫字母表示4種灌木間差異顯著(P<0.05),相同小寫字母表示差異不顯著(P>0.05)。
Note:The biomass was the dry weight of 2010;Different lower case letters within the same row indicate significant difference at 0.05level.
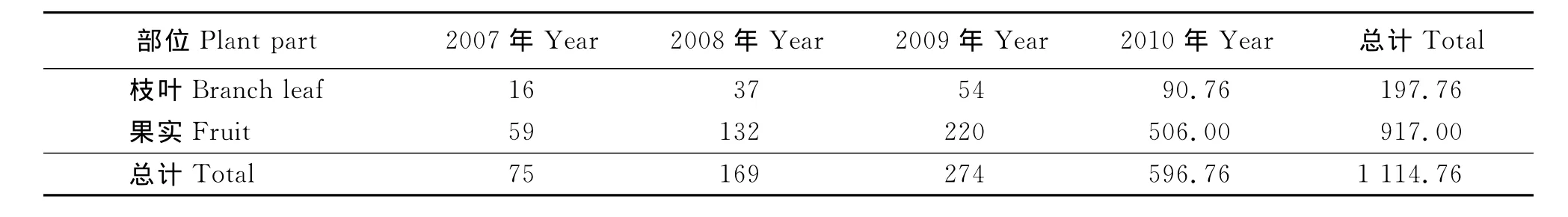
表3 寧夏枸杞歷年枝葉修剪量與干果產量Table 3 Amount of branch leaves pruning and fruit yield of Lycium barbarum g
3 討論
3.1 全生長季單位葉片及全株葉片光合速率是反映植物碳同化率能力的可靠指標 光合速率是指植物單位葉面積單位時間同化CO2的量。國內有學者[13-16]在研究植物固碳釋氧能力時,將單位葉面積的光合速率(或單位葉面積日同化總量)作為衡量不同植物固碳釋氧能力的指標。從本研究結果看,植物單位葉面積的光合速率值和全生長季的光合速率值并不相等,而且認為全年的光合速率才是衡量植物固碳釋氧能力的全面準確的指標,與前人觀點有所不同。
另外,從本研究結果看,單位葉片光合速率(碳同化率)高的植物其全株的日均碳同化量不一定高,因為其全株的葉面積不一定大。因此,研究某一植物的固碳能力及年固碳量就應當研究全株植物在全年的碳同化率或凈光合速率,進而估算出全株植物的年固碳量。也有學者[17-18]采用單位葉面積凈同化率結合植株或植物群體的葉面積指數,有效地比較了不同植物間固碳能力,這與本研究的方法相似。
3.2 具有修剪等農藝措施的寧夏枸杞經濟林具有很大的碳匯潛力 本研究結果表明,由于作為經濟作物的寧夏枸杞具有良好的水肥管理條件,而且修剪的農藝措施還有可能促進其補償性生長,如果將修剪的枝條和采摘的果實累加起來,則隨著栽培時間的延長,寧夏枸杞的生物量積累在4種灌木中最大,其碳匯最終會超過其他3種灌木。因此,可以推測,經濟林較其他灌木林可能具有更大的碳匯潛力。俞益武和徐秋芳[19]的研究結果也顯示,天然林改為經濟林后土壤的含碳量明顯提高,說明其固碳能力得到了提高。根據孫穎等[20]對寧夏森林生態系統服務功能價值的統計結果顯示,經濟林的固碳制氧功能在其生態系統服務功能中居于首位,而且在各類森林生態系統中也是處于前列。據王兵和魯紹偉[21]報道,目前全國經濟林面積為2 139萬hm2,占全國有林地面積的12.66%,其固碳釋氧效益是不可估量的。因此,經濟林是經濟效益和生態效益兼具的生態系統,應當重視經濟林的科學建設。
[1]王俊明,張興昌.退耕草地演替過程中的碳儲量變化[J].草業學報,2009,18(1):1-8.
[2]李琪,王云龍,胡正華,等.基于渦度相關法的中國草地生態系統碳通量研究進展[J].草業科學,2010,27(12):38-44.
[3]王鶯,夏文韜,梁天剛.陸地生態系統凈初級生產力的時空動態模擬研究進展[J].草業科學,2010,27(2):77-88.
[4]吳慶標,王效科,段曉男,等.中國森林生態系統植被固碳現狀和潛力[J].生態學報,2008,28(2):517-524.
[5]王兵,王燕,趙廣東,等.中國森林生態系統碳平衡研究進展[J].內蒙古農業大學學報,2008,29(2):194-199.
[6]趙林,殷鳴放,陳曉非,等.森林碳匯研究的計量方法及研究現狀綜述[J].西北林學院學報,2008,23(1):59-63.
[7]劉盛,李國偉.林分碳貯量測算方法的研究[J].北京林業大學學報,2007,29(4):166-169.
[8]王蕾,張景群,王曉芳,等.黃土高原兩種人工林幼林生態系統碳匯能力評價[J].東北林業大學學報,2010,38(7):75-78.
[9]賀紅早,賀瑞坤,段旭,等.貴陽二環林帶主要造林樹種碳匯研究[J].安徽農業科學,2007,35(32):10270-10271.
[10]李海梅,何興元,宋力.3種灌木樹種光合特性及影響因子研究[J].沈陽農業大學學報,2007,38(4):605-608.
[11]王麗勉,胡永紅,秦俊,等.上海地區151種綠化植物固碳釋氧能力的研究[J].華中農業大學學報,2007,26(3):399-401.
[12]馬驥,倪細爐,史宏勇,等.蒙古扁桃的開花生物學研究[J].西北植物學報,2010,30(6):1134-1141.
[13]陸貴巧,尹兆芳,谷建才,等.大連市主要行道綠化樹種固碳釋氧功能研究[J].河北農業大學學報,2006,29(6):49-51.
[14]焦緒娟,趙文飛,張衡亮,等.幾種綠化樹種降低城市熱島效應的研究[J].江西農業大學學報,2007,29(1):89-93.
[15]劉海榮,宋力,鮮靖蘋.5種常用灌木固碳釋氧能力的比較研究[J].安徽農業大學學報,2009,36(2):204-207.
[16]趙萱,李海梅.11種地被植物固碳釋氧與降溫增濕效益研究[J].江西農業學報,2009,21(1):44-47.
[17]丁向陽.南陽市城市森林主要植物的生態效益[J].中南林業科技大學學報,2007,27(4):142-146.
[18]徐瑋瑋,李曉儲,汪成忠,等.揚州古運河風光帶綠地樹種固碳釋氧效應初步研究[J].浙江林學院學報,2007,24(5):575-580.
[19]俞益武,徐秋芳.天然林改為經濟林后土壤微生物量的變化[J].水土保持學報,2003,17(5):103-113.
[20]孫穎,王得祥,張浩,等.寧夏森林生態系統服務功能的價值研究[J].西北農林科技大學學報(自然科學版),2009,13(27):91-97.
[21]王兵,魯紹偉.中國經濟林生態系統服務價值評估[J].生態學報,2009,20(2):417-425.
A comparison of photosynthetic carbon sequestration of four shrubs in Ningxia
LI Zhi-gang,ZHU Qiang,LI Jian
(The State Key Laboratory of Seedling Bioengineering Ningxia Forestry Institute,Yinchuan 750004,China)
Carbon sequestration of four kinds shrubs in Ningxia was studied by testing leaf photosynthetic rate and plant biomass.The results indicated that if the research was on photosynthesis of each individual leaf,carbon sequestration was in the order ofBuddlejaalternifolia>Amygdalusmongolica>Atraphaxis bracteata>Lyciumbarbarumin the vigorous growth season(in July);orL.barbarum>B.alternifolia>A.bracteata>A.mongolicain the whole growth season.We also measured a single-leaf area,the area of whole plant leaves and the carbon sequestration of whole plant.The results indicated that sequence of the average daily amount of carbon assimilation of the whole plant wasB.alternifolia>L.barbarum>A.bracteata>A.mongolicain both of the vigorous growth season and the whole growth season.The similar results were obtained on the measurement of biomass.Therefore,it can be concluded that the plant carbon sequestration in whole plant leaves should be considered if the photosynthetic rate is used as a testing target.In addition,it should be paid attention that there is a strong carbon sequestration ofL.barbarumforests due to the good maintenance,such as irrigation and fertilizer.
Ningxia;shrubs;carbon sequestration;photosynthesis
LI Jian E-mail:lijian0630@yahoo.com.cn
Q945.11
A
1001-0629(2012)03-0352-06
2011-04-20 接受日期:2011-06-20
寧夏自然基金項目“寧夏枸杞經濟林生態系統碳匯能力的初步研究”(NZ10210);寧夏科技攻關項目“銀川腹部沙地植物碳匯能力評價與沙化土壤恢復模式研究”
李志剛(1985-),男,寧夏海原人,助理研究員,碩士,主要從事生態學與農林廢棄物資源開發利用方向的研究工作。
E-mail:lizg001@sina.com
李健 E-mail:lijian0630@yahoo.com.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
