疏棉狀嗜熱絲孢菌耐熱脂肪酶在無孢黑曲霉中的高效表達
2012-09-11 13:11:20李德明
食品工業科技 2012年20期
陳 婧,王 斌,李德明,潘 力
(華南理工大學生物科學與工程學院,廣東廣州510006)
疏棉狀嗜熱絲孢菌耐熱脂肪酶在無孢黑曲霉中的高效表達
陳 婧,王 斌,李德明,潘 力*
(華南理工大學生物科學與工程學院,廣東廣州510006)
綜合考慮密碼子偏愛性、RNA穩定性以及自由能等因素,對疏棉狀嗜熱絲孢菌(Thermomyces lanuginosus)耐熱脂肪酶(tll)基因序列進行密碼子優化并全基因合成。將合成的tll基因連接到克隆載體,然后亞克隆到表達載體pHGWPT-tll,采用根癌農桿菌介導轉化無孢黑曲霉SH-2。經過三丁酸甘油酯平板、SDS-PAGE以及Western Blotting鑒定,成功表達了tll基因。發酵72h,轉化子脂肪酶酶活最高達36U/mL。轉化子經等離子誘變得到的突變株的脂肪酶比活力比出發菌株提高約37%。
疏棉狀嗜熱絲孢菌,耐熱脂肪酶,黑曲霉,重組表達
Abstract:We optimized the codon usage of Thermomyces lanuginosus lipase(tll) gene based on codon usage bias,RNA stability and free energy,and synthesized the optimized tll gene.The obtained tll gene was inserted into a clone vector,and then subcloned to the expression vector pHGWPT-tll.The vector pHGWPT-tll was transformed into non-spore Aspergillus niger SH-2 via Agrobacterium tumefaciens-mediated transformation using hygromycin B resistant gene as the selection marker.A.niger transformants bearing the tll gene,were selected by tributyrin hydrolyzing plate.The recombinant protein was verified by SDS-PAGE and Western blotting.After 72h fermentation,the lipase activity of transformants reached 36U/mL.Furthermore,the tll transformants was subjected to mutagenesis via atmospheric and room temperature plasma(ARTP).The specific activity of the mutants was 37%higher than that of the original transformants,suggesting that tll gene could be highly expressed in A.niger.
Key words:Thermomyces lanuginosus;thermostable lipase;Aspergillus niger;recombinant expression
黑曲霉具有很強的蛋白分泌能力[1],蛋白產量可達到30g/L以上[2],是重要的蛋白生產平臺之一[2-4],被美國FDA認定為安全菌株(GRAS),使其在各領域中受到的關注與日俱增[5]。隨著根癌農桿菌介導的轉化(Arobacterium tumefaciens mediated transformation,ATMT)在絲狀真菌中的成功應用[6-10],黑曲霉更多被作為轉化宿主進行外源基因表達[11]。一般黑曲霉分生孢子頭呈褐黑色、分生孢子梗長短不一(圖1A),本研究使用的無孢黑曲霉為工業誘變菌,有很強的糖化酶分泌能力,不產孢子、菌落呈乳白色且菌絲短小(圖1B)。粗短的菌絲易形成小而松散的菌球,有益于代謝產物的產生[12],因此該菌擁有良好的工業應用前景。疏棉狀嗜熱絲孢菌(T.lanuginosus)是一種嗜熱真菌,其脂肪酶具有專一性強、選擇性好、穩定性高等優點[13],廣泛應用于有機酸合成[14]、食物加工及油的水解等行業,具有重要的經濟價值。Wai Prathumpai等[15]將其在黑曲霉(NW297-14和NW297-24)中表達,以硝基苯基丁酸為底物,酶活達到6U/mL;鄭艷等[16]以畢赤酵母為宿主,以橄欖油為底物,酶活達到7.2U/mL。目前,還沒有將疏棉狀嗜熱絲孢菌耐熱脂肪酶在無孢黑曲霉中表達的報道。本研究使用黑曲霉糖化酶啟動子構建表達載體,利用根癌農桿菌介導tll基因轉化至無孢黑曲霉SH-2,然后使用突變率高、突變譜廣、重復性好[17]的等離子誘變技術進行誘變,實現tll在無孢黑曲霉中的高效重組表達。
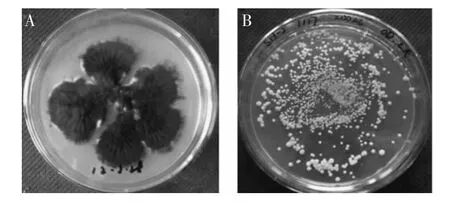
圖1 淀粉培養基培養的黑曲霉MA169.4和無孢黑曲霉SH-2Fig.1 Aspergillus niger MA169.4 and Sporeless Aspergillus niger SH-2 cultured on solid starch medium
1 材料與方法
1.1 材料與儀器
黑曲霉SH-2 糖化酶生產企業;根癌農桿菌LBA4404 廣東省農科院;疏棉狀嗜熱絲孢菌耐熱脂肪酶基因tll(3’端添加His6組氨酸標簽)南京金斯瑞生物科技有限公司合成;雙元表達載體質粒pHGW(含潮霉素抗性基因HygBr)比利時植物學研究中心(VIB);大腸桿菌Mach1T1、ccdB Survival 本實驗室保藏;潮霉素 美國Merck公司;乙酰丁香酮sigma-aldrich公司;三丁酸甘油酯 百靈威公司;限制性內切酶 美國Fermentas公司;PCR引物 Invitrogen合成;LC、IM和MM培養基[9],淀粉培養基(g/L) 牛肉膏3,蛋白胨10,酵母粉2,淀粉10,NaCl 2,瓊脂粉17,pH5.5;CD培養基(g/L) 葡萄糖20,硝酸鈉3,KCl 0.5,MgSO40.5,K2HPO41、FeSO40.01,瓊脂粉15,pH5.5。
Centrifuge5418型離心機、Mastercycler gradient PCR儀 德國eppendorf公司;?KTA explorer蛋白純化系統 美國GE公司;ARTP生物育種機 北京思清源生物科技公司等。
1.2 實驗方法
1.2.1 耐熱脂肪酶基因表達載體的構建 利用引物Sig-forward/Sig-reverse PCR擴增黑曲霉糖化酶基因的信號肽及起始密碼子ATG上游10bp序列(Kozak序列);利用引物TLL-forward/TLL-reverse PCR擴增經密碼子優化的tll基因,再利用融合PCR,將上述兩個片段連接成一個DNA片段,命名為Sig-tll。
以黑曲霉基因組DNA為模板,利用引物PglaA-forward/PglaA-reverse擴增黑曲霉糖化酶啟動子(PglaA),利用引物TglaA-forward/TglaA-reverse擴增糖化酶終止子(TglaA),再利用融合PCR,將上述兩個片段連接成一個DNA片段,命名為PT。PT片段經連接克隆載體pSIMPLE-18 EcoRV/BAP Vector后,測序驗證正確后用HindIII/XbaI雙酶切亞克隆到農桿菌雙元載體pHGW,命名為pHGWPT。
用PvuII酶切質粒pHGWPT并去磷酸化處理,將片段Sig-tll經磷酸化處理后連接到質粒pHGWPT上的啟動子(PglaA)和終止子(TglaA)之間,通過PCR篩選方向性,挑選方向正確的重組子測序驗證,質粒命名為pHGWPT-tll。所有引物序列見表1。
1.2.2 根癌農桿菌介導耐熱脂肪酶表達載體轉化無孢黑曲霉SH-2 參照朱錦輝等[18]報道的方法,將構建的pHGWPT-tll表達載體轉化根癌農桿菌LBA4404,挑選陽性轉化子并提取質粒進行鑒定。
參照Caroline B Michielse等[9]報道的轉化方法,利用根癌農桿菌介導pHGWPT-tll轉化無孢黑曲霉SH-2并得到轉化子。將轉化子單菌落轉接至含0.5%(v∶v)三丁酸甘油酯的CD固體培養基(三丁酸甘油酯平板)上,30℃培養2d,觀察轉化子周圍有無水解圈出現,對轉化子進行初步鑒定。分別提取宿主菌SH-2和轉化子的基因組DNA,利用引物glaA-forward/glaA-reverse、TLL-forward/TLL-reverse及hygB-forward/hygB-reverse分別擴增黑曲霉糖化酶基因(glaA,約1kb)、耐熱脂肪酶基因(Sig-tll,約1kb)及潮霉素抗性基因(HygBr,約1kb)進行鑒定。
使用2%糖(1%葡萄糖+1%麥芽糖)為碳源的YPD培養基,30℃,250r/min,分別對宿主菌SH-2和轉化子發酵3d。攜帶His6組氨酸標簽的tll蛋白能與親和層析柱牢固結合而進行有效洗脫,因此將發酵液離心過濾后使用His-tag親和層析柱純化,純化后轉化子的tll蛋白使用考馬斯亮藍法測定濃度;以橄欖油為底物,采用標準方法[19-20]測定脂肪酶活力;并進行SDS-PAGE電泳檢測和Western Blotting進一步鑒定。酶活力單位定義為每分鐘催化底物水解產生1μmol脂肪酸的酶量。
1.2.3 等離子突變株的選育及鑒定 使用ARTP育種機對轉化子進行等離子誘變。參照樣品制備流程制備樣品,選取時間梯度30、60、90、120、180s進行誘變,誘變后的菌液分別稀釋50、500、5000倍涂布于淀粉培養基,30℃培養5d。挑取菌落大的單菌落轉接于三丁酸甘油酯平板上,通過水解圈的大小初步判斷脂肪酶活力,對突變株進行篩選。

表1 PCR引物序列Table 1 The sequences of PCR primers
經過二次篩選,選定水解圈最大的突變株單菌落研究,30℃、250r/min對其發酵,測定純化后突變株tll蛋白的濃度和脂肪酶活力。
1.2.4 耐熱脂肪酶的生物學特性 耐熱脂肪酶的最適反應溫度:選取溫度梯度50、55、60、65、70、75、80℃,以橄欖油為底物進行脂肪酶活測定,酶活最高時的溫度即為重組耐熱脂肪酶的最適反應溫度。
耐熱脂肪酶的最適反應pH:以橄欖油為底物,調整反應緩沖液pH分別為3.0、5.0、7.0、8.0、9.0、11.0,進行脂肪酶活測定,酶活最高時的pH即為重組耐熱脂肪酶的最適反應pH。
2 結果與討論
2.1 耐熱脂肪酶基因表達載體的構建
通過融合PCR擴增耐熱脂肪酶基因(Sig-tll),電泳結果顯示基因片段大小與預計一致(圖2A),通過亞克隆得到耐熱脂肪酶的表達載體pHGWPT-tll(圖2B)。HindIII/XbaI雙酶切表達載體pHGWPT-tll,得到與目的片段大小相符的片段(約2.7K,圖2C),表明耐熱脂肪酶表達載體構建成功。

圖2 Sig-tll基因的擴增及耐熱脂肪酶表達載體的酶切鑒定Fig.2 PCR amplification of Sig-tll gene and identification of its expression vector by enzyme digestion
2.2 黑曲霉轉化子的鑒定
挑選MM培養基上菌落大的單菌落(圖3A),轉移至三丁酸甘油酯平板。培養2d,出現水解圈的菌落,初步確定為轉化子,而宿主菌SH-2菌落周圍無水解圈(圖3B)。

圖3 pHGWPT-tll表達載體轉化無孢黑曲霉SH-2Fig.3 Transformation of sporeless A.niger SH-2 with expression vector pHGWPT-tll
分別以宿主菌SH-2和轉化子的基因組為模板,經PCR擴增所得的glaA、HygBr及Sig-tll片段大小與預計一致(圖4A),且Sig-tll基因測序結果與原始序列一致,證實所得轉化子為陽性克隆。SDS-PAGE電泳結果顯示,轉化子在32ku處有蛋白條帶,與tll蛋白大小相符[16],Western Blotting鑒定表明,轉化子有對應耐熱脂肪酶tll的條帶,而宿主菌SH-2沒有(圖4B),進一步確定轉化子成功表達疏棉狀嗜熱絲孢菌耐熱脂肪酶。

圖4 轉化子的鑒定Fig.4 Verification of transformant
2.3 等離子突變株的鑒定
對轉化子進行等離子誘變,經二次篩選得到突變株。結果顯示,宿主菌SH-2(圖5A,1)在三丁酸甘油酯平板不形成水解圈,轉化子(圖5B,1)形成明顯的水解圈。對水解圈最大的突變株(圖5B,2)進行發酵,純化后轉化子、突變株tll蛋白的濃度、脂肪酶活力及酶的比活力見表2。結果顯示,雖突變株和轉化子的脂肪酶活力相差不大,但同轉化子相比,突變株的比活力提高37%。此外,轉化子和突變株的脂肪酶活力分別達到36、35U/mL,遠高于疏棉狀嗜熱絲孢菌脂肪酶在畢赤酵母中的表達水平(7.2U/mL)[16]。
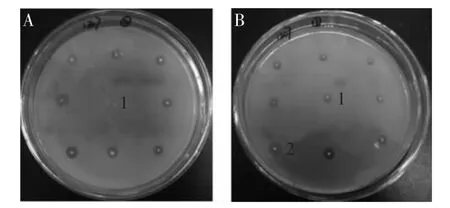
圖5 等離子突變株的篩選Fig.5 Screening of lipase mutant

表2 純化后轉化子、突變株tll蛋白濃度、酶活力及酶的比活力Table 2 Protein concentration,enzyme activity and specific activity of tll after purification
2.4 耐熱脂肪酶最適反應溫度及最適反應pH
不同pH緩沖液環境下,分別測定宿主菌SH-2和突變株的脂肪酶活力。結果顯示,宿主菌SH-2的脂肪酶活力在緩沖液pH為3.0~11.0之間較低且沒有明顯變化;突變株在緩沖液pH為3.0~11.0區間脂肪酶活力較高,且在緩沖液pH8.0時,酶活力最高,超過pH8.0酶活力顯著下降。
在不同溫度下分別測定宿主菌SH-2和突變株的脂肪酶活力。結果顯示,宿主菌SH-2的脂肪酶活力在50~80℃之間較低且無較大變化;突變株則在50~60℃區間保持較高酶活力,且酶活力在60℃時達到最高,超過60℃,酶活力顯著下降。上述結果與疏棉狀嗜熱絲孢菌脂肪酶在畢赤酵母中表達時基本一致[18]。

圖6 耐熱脂肪酶最適反應pH及最適反應溫度Fig.6 The optimum pH and optimum temperature of the tll lipase
3 結論
疏棉狀嗜熱絲孢菌耐熱脂肪酶具有作用溫度高、副產物少、作用穩定等優點,得到越來越多的關注。該酶傳統的生產方法是通過內源分泌得到的,產物量少、質量參差不齊、無法產業化等限制了其應用。目前,對疏棉狀嗜熱絲孢菌耐熱脂肪酶的重組表達研究較少,國外Wai Prathumpai等使用米曲霉淀粉酶啟動子將其在黑曲霉(NW297-14和NW297-24)中表達,但是酶活力較低。本研究對疏棉狀嗜熱絲孢菌耐熱脂肪酶基因進行密碼子優化,使其自由能由-291.9kcal/mol降至-303.3kcal/mol。自由能降低,大大減少了mRNA兩端二級結構的產生,增加mRNA穩定性從而提高蛋白表達量。研究使用黑曲霉自身糖化酶強啟動子構建質粒,經根癌農桿菌介導轉化至無孢黑曲霉SH-2,得到的轉化子經過連續傳代培養后,穩定性良好且脂肪酶活力達到36U/mL。在此基礎上,使用等離子誘變技術對轉化子進行誘變,效果顯著,實現了耐熱脂肪酶在無孢黑曲霉的重組高效表達。
本文采用的潮霉素抗性基因(HygBr)雖然在真菌研究中應用極為廣泛,但潮霉素是一種細菌毒素,屬于氨基環多醇類抗生素,它可通過干擾蛋白合成而抑制原核和真核生物的生長,這極大的限制了其在食品等工業的應用。因此選擇一種能夠應用于食品工業的篩選標記如營養互補型選擇標記,使其能夠廣泛應用于食品工業,為該酶的工業化生產提供條件,這將是本課題今后深入研究的方向。
[1]De Bekker C,van Veluw G J,Vinck A,et al.Heterogeneity of Aspergillus niger microcolonies in liquid shaken cultures[J].Applied and environmental microbiology,2011,77(4):1263-1267.
[2]Punt P J,van Biezen N,Conesa A,et al.Filamentous fungi as cell factories for heterologous protein production[J].Trends in biotechnology,2002,20(5):200-206.
[3]Conesa A,Punt P J,van Luijk N,et al.The secretion pathway in filamentous fungi:a biotechnological view[J].Fungal genetics and biology:FG&B,2001,33(3):155-171.
[4]Lubertozzi D,Keasling J D.Developing Aspergillus as a host for heterologous expression[J].Biotechnology advances,2009,27(1):53-75.
[5]Perrone G,Stea G,Epifani F,et al.Aspergillus niger contains the cryptic phylogenetic species A.awamori[J].Fungal biology,2011,115(11):1138-1150.
[6]Combier J.P,Melayah D,Raffier C,et al.Agrobacterium tumefaciens-mediated transformation as a tool for insertional mutagenesis in the symbiotic ectomycorrhizal fungus Hebeloma cylindrosporum[J].FEMS microbiology letters,2003,220(1):141-148.
[7]Michielse C B,Hooykaas P J,van den Hondel C A,et al.Agrobacterium-mediated transformation as a tool for functional genomics in fungi[J].Current genetics,2005,48(1):1-17.
[8]Yang L,Yang Q,Sun K,et al.Agrobacterium tumefaciens mediated transformation of ChiV gene to Trichoderma harzianum[J].Applied biochemistry and biotechnology,2011,163(8):937-945.
[9]Michielse C B,Hooykaas P J,van den Hondel C A,et al.Agrobacterium-mediated transformation of the filamentous fungus Aspergillus awamori[J].Nature protocols,2008,3(10):1671-1678.
[10]潘力,李俊星,苗小康.根癌農桿菌介導酸性蛋白酶在泡盛曲霉中表達的研究[J].食品工業科技,2010,32(5):5.
[11]Schuster E,Dunn-Coleman N,Frisvad JC,et al.On the safety of Aspergillus niger-a review[J].Applied microbiology and biotechnology,2002,59(4-5):426-435.
[12]楊海龍,吳天祥,章克昌.絲狀真菌發酵生產中形態的影響與發酵罐設計[J].生物技術,2003,13(1):3.
[13]Roberto Fernandez-Lafuente.Lipase from Thermomyces lanuginosus:Uses and prospects as an industrial biocatalyst[J].Journal of Molecular Catalysis B:Enzymatic,2010,62:15.
[14]Brzozowski AM,Derewenda U,Derewenda ZS,et al.A model for interfacial activation in lipases from the structure of a fungal lipase-inhibitor complex[J].Nature,1991,351:491-494.
[15]Prathumpai W,Flitter SJ,McIntyre M,et al.Lipase production by recombinant strains of Aspergillus niger expressing a lipaseencoding gene from Thermomyceslanuginosus[J].Applied microbiology and biotechnology,2004,65(6):714-719.
[16]鄭艷,周波,宋寧寧,等.疏棉狀嗜熱絲孢菌脂肪酶基因的克隆及其在畢赤酵母中的高效表達[J].菌物學報,2009,28(2):6.
[17]曾憲賢,武寶山,呂杰.離子束生物技術在生命科學中的應用[J].核技術,2006,29(2):3.
[18]朱錦輝,權軍利,何玉科,等.根癌農桿菌感受態細胞的制備以及質粒ProkⅡ對其轉化的研究[J].西北農林科技大學學報,2006,34(7):4.
[19]K.ARIMA,W-H.Liu,T.BEPPU.Studies on the Lipase of Thermophilic Fungus Humicola lanuginosa[J].Agr Biol Chem,1972,36(5):2.
[20]滕宏飛,王丹靜,徐青,等.橄欖油乳化法測定脂肪酶活性的優化研究[J].食品工業,2011(6):3.
Effectively expression of Thermomyces lanuginosus thermostable lipase in non-spore Aspergillus niger
CHEN Jing,WANG Bin,LI De-ming,PAN Li*
(School of Bioscience and Bioengineering,South China University of Technology,Guangzhou 510006,China)
TS201.3
A
1002-0306(2012)20-0160-40
2012-04-23 *通訊聯系人
陳婧(1987-),女,碩士研究生,研究方向:工業微生物。
國家自然科學基金項目(30970045);“十二五”農村領域國家科技計劃(2011AA100905)。
