基因芯片篩選畜禽熱應激差異表達基因的研究進展
2012-09-11 07:35:26劉麗莉謝紅兵楊永生許丹寧李江長賀建華
動物營養(yǎng)學報 2012年12期
關鍵詞:差異
劉麗莉 謝紅兵 楊永生 許丹寧 李江長 賀建華*
(1.湖南農業(yè)大學動物科學技術學院,長沙 410128;2.湖南科技大學生命科學學院,湘潭 411201)
高溫應激是影響夏季畜禽生產的重要因素,可造成畜禽生長和代謝紊亂、免疫力下降及肉品質改變等,給畜牧業(yè)帶來很大的經濟損失。熱應激反應是動物機體通過動員自身防御機能克服應激因子以避免組織器官損傷的一種非特異性防御反應,若熱應激作用時間過長、強度過大,機體會逐漸失去這一應對能力,出現(xiàn)病理及衰竭狀態(tài)。因此,迫切需要對畜禽熱敏感、熱耐受性進行準確判斷和對熱應激損傷的分子機制進行更全面系統(tǒng)的研究。目前,基因芯片技術已成功運用于生物分子標志物的篩選,并在全基因組內同時分析待測樣本中成千上萬個基因的表達情況及其相互關系,篩選出一系列差異表達的候選基因,為尋找目標基因提供了一個強有力的手段[1]。熱應激引起畜禽各階段的發(fā)生發(fā)展都與組織器官的多基因表達異常有關,基因芯片技術可以識別出動物熱應激進程中基因表達的改變[2-3],在大鼠、豬的熱應激研究中,其基因的差異表達主要表現(xiàn)為熱休克蛋白、氧化還原基因、代謝相關基因、轉錄調控基因、細胞凋亡基因、免疫基因的表達發(fā)生改變[3-4]。本文旨在就近年來基因芯片技術在篩選畜禽熱應激差異表達基因的研究和應用進行綜述,探討熱應激動物生理過程中熱敏感基因的變化以及各基因間互作的動態(tài)效應,為深入研究畜禽熱應激損傷的分子營養(yǎng)調控機制提供理論基礎。
1 基因芯片技術的原理
基因芯片也稱為基因微陣列(cDNA microarray)或寡核苷酸微陣列(oligonucleotide microarray),應用已經破解的全基因組已知核酸序列作為探針與互補的靶核苷酸序列雜交,使得合成、固定高密度的數(shù)以萬計的探針分子以及對雜交信號進行實時、靈敏、準確的檢測分析(圖1)[2]。與傳統(tǒng)檢測RNA表達水平的技術相比,基因芯片可高敏感地定量、定性檢測差異基因表達水平,最大限度地達到同步、多靶點和高通量地研究成百上千個基因差異表達情況。基因芯片已廣泛應用于功能基因組研究、疾病分子診斷、突變檢測及藥物篩選等領域,在畜禽生產研究中發(fā)揮越來越大的作用。
畜禽熱應激反應機制是一個多步驟、多基因調控的復雜過程,并不是一兩個或少數(shù)幾個基因所能決定的,利用基因芯片技術對動物熱應激過程中的基因表達譜進行研究,成功發(fā)現(xiàn)了涉及動物生長代謝、炎癥反應、細胞內信號傳遞及生長、凋亡因子等上百條表達上調或下調基因[3,5],篩選了與熱應激損傷修復密切相關的基因,繼而開展了關鍵基因的功能研究,使該技術在探討畜禽熱應激發(fā)生機制中產生了重要的應用價值。
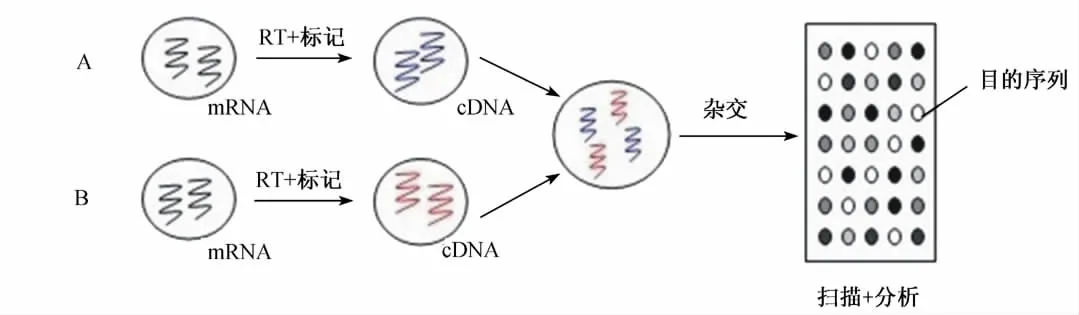
圖1 cDNA微陣列技術分析基因表達的原理Fig.1 The principle of gene expression analysis by cDNA microarray technology
2 基因芯片技術在熱應激研究中的應用
2.1 熱應激引起畜禽器官氧化損傷相關的差異表達基因研究
2.1.1 腸道
熱應激時,動物胃腸道血流量急劇減少,缺血缺氧產生大量活性氧自由基(reactive oxygen species,ROS)而造成腸道黏膜損傷,從而誘導細胞凋亡[6],而通過調節(jié)各種生長因子表達水平來促進隱窩細胞的分裂增殖,可使受損小腸上皮細胞得以修復[7-8]。Liu 等[7]將含有 23 937 條探針代表已知豬20 201個基因的Affmetrix GeneChip檢測熱應激下豬空腸組織的基因表達譜,可知差異表達基因有143個,其中上調明顯的基因有趨化因子2(CXCL2)、谷胱甘肽S轉移酶(GST)、谷胱甘肽過氧化物酶2(GPX2)等68個,下調明顯的基因有表皮生長因子(EGF)、金屬硫蛋白(MT1A)、細胞色素P450家族(CYP2B22)等75個,基因本體論(GO)分類表明,熱應激影響豬免疫、內分泌、細胞損傷修復、信號轉導相關基因的表達;Pathway分析表明,占1/3比例的差異基因信號通道與小腸上皮EGF介導的細胞再生、修復有關。熱應激可誘導CXCL2上調,促進中性粒細胞的游走和趨化且吸引免疫細胞到達免疫應答區(qū),參與免疫調控和免疫病理反應[9],GST和 GPX2的上調催化還原型谷胱甘肽與親電子有害化合物的結合,并清除體內沉積的各種內生或外源性過氧化物而減緩機體熱應激損傷[10]。
Yu等[4]利用基因芯片篩選到203個與豬小腸熱應激相關的差異表達基因,包括熱休克蛋白(HSP)70、HSP90、HSP27等93個表達顯著上調和EGF、表皮生長因子受體(EGFR)等110個顯著下調基因,表達差異基因主要與熱休克、物質代謝、抗氧化、信號轉導、細胞凋亡、細胞分裂和增殖等有關。Lu等[11]通過基因芯片檢測熱應激對大鼠小腸差異表達基因有422個,包括290個上調基因和132個下調基因,差異表達基因主要集中為熱休克蛋白家族(HSPs)超表達,如HSP27(HSPb1)、HSP70(HSPa1a、HSPa11、HSPa8)、HSP90(HSPcb)和HSP110(HSPh1)高表達以抵抗熱應激的損傷,一些細胞因子參與抵抗炎癥并調節(jié)免疫功能。熱應激狀態(tài)下HSPs表達量增多,多種蛋白質與其形成復合體,通過其解離與結合來協(xié)助折疊新生蛋白質,防止發(fā)生聚集現(xiàn)象;同時幫助變性蛋白質的復性和蛋白質的線粒體跨膜移位,并經由泛素-蛋白酶體通路清除嚴重受損蛋白質等作用機制[12],抵御熱應激以維持機體正常的生命活動。
熱應激導致腸道上皮細胞嚴重受損且明顯減退小腸屏障和吸收功能,Yu等[4]利用基因芯片并結合miRNA和mRNA的基因表達譜研究了熱應激對大鼠空腸組織的差異基因表達,mRNA基因芯片分析發(fā)現(xiàn)382個差異表達基因,其中上調270個,下調122個,mRNA的差異表達分析主要從分子功能、生物進程、細胞組成和KEGG(kyoto encyclopedia of genes and genomes)代謝途徑進行分類;miRNA基因芯片發(fā)現(xiàn)29個差異表達基因,其中上調18個,下調11個,熱應激誘導小腸上皮細胞損傷和凋亡,導致 miR-34a、miR-34b、miR-137等明顯上調。而 miR-34a、miR-34b 等 miRNAs作為凋亡基因(p53)的作用靶標可參與調控DNA損傷修復、細胞周期阻滯、細胞凋亡等事件[13-14],miR-34(包括 miR-34a、miR-34b 和 miR-34c)通過抑制多能性相關聯(lián)的基因從而阻止體細胞重編程[15],大 鼠 局 部 缺 血 可 造 成 miR-137 表達上調[16],組織損傷加劇使得 miR-137 表達顯著增加[17]。此外,通過生物信息學分析證明與熱應激相關的mRNA表達變化與miRNA表達呈負相關[4]。
2.1.2 肝臟
Bhusari等[18]對熱應激大鼠肝臟組織的基因芯片結果進行分析并確定了19個差異表達的靶標基因,其中包括乙醛脫氫酶2(ALDH2)、金屬硫蛋白1(MT-1)、ATP合酶 β 亞基(ATP5b)、蛋白酶體 β5(PSMB5)、酪蛋白激酶 2α(CSNK2a1)、細胞周期依賴激酶K4(CDK4)等12個涉及抗氧化通路和代謝相關的上調基因,及ATP合酶亞基8(ATP8)、過氧化氫酶(CAT)、細胞色素 P450(CYP2e1)等7個與活性氧自由基和線粒體表達相關的下調基因。ALDH2可通過在線粒體基質內清除 ROS 而起抗氧化作用[19],MT-1 能有效清除羥自由基并螯合有毒重金屬[20]。熱應激24~48 h可使CAT表達量呈現(xiàn)2倍峰值而后下降[21],在小鼠中過表達人的CAT能顯著減少氧化損傷、DNA變異及H2O2等產物[22]。熱應激發(fā)生時,HSP1可在熱誘導激酶的信號傳導下激活細胞內絲氨酸/蘇氨酸蛋白激酶相關的信號通路[23],CSNK2a1能修復斷裂的染色體DNA鏈且阻止半胱天冬酶8(caspase-8)所誘導的細胞凋亡[24],CDK4 是一類絲氨酸/蘇氨酸蛋白激酶,由催化亞基和調節(jié)亞基(周期蛋白)組成,對細胞周期G1期的分裂增殖起到重要的調控作用,在抗氧化調控途徑中上調ALDH2、MT-1、CSNK2a1、CDK4 等基因而對熱應激起抵抗作用,以減緩肝臟組織的應激損傷[18]。
肝細胞微粒體中的CYP2e1參與異生物質代謝,依賴NADPH通過電子傳遞途徑完成催化反應。CYP2e1能通過NADPH氧化酶和異生代謝產物直接產生氧化應激[25],如果CYP2e1過表達,那么肝細胞內產生的ROS將導致脂質過氧化而促使肝細胞壞死,甚至凋亡。熱應激可刺激CYP2e1產生ROS而引起肝臟毒性損傷,下調CYP2e1能調節(jié)小鼠適應慢性熱應激下的內源ROS發(fā)生機制,啟動并上調CAT、ALDH2、SOD等抗氧化防御反應系統(tǒng)[18]。PSMB5是26S蛋白酶體的重要組成部分,主要對細胞內發(fā)生錯誤折疊、突變或非正常短期存在的蛋白質進行修飾或降解,并直接參與氧化蛋白的降解和泛素-蛋白酶體通路[26]。PSMB5啟動子區(qū)域參與核轉錄因子(nuclear transcription factor 2,Nrf2)介導的抗氧化反應元件(antioxidantresponse elements,AREs)的結合,在一定程度上PSMB5表達水平可成為氧化應激狀態(tài)的征兆,PSMB5表達上調則能有效清除熱應激所造成的機體損傷性蛋白,以抵抗熱應激來維持細胞的生命活力[18]。肝臟分解代謝過程中,高密度脂蛋白(HDL)通過干擾作用于內皮細胞的促凋亡因子而起保護作用,HDL攜帶的脂蛋白和酶具有抗氧化的功能[27]。ATP5b是ATP合酶的β鏈,肝細胞膜上的一種載脂蛋白-I(APOA-I)的高親和力受體,這種受體在HDL的代謝機理和ATP的生物合成中起重要作用,APOA-I激活ATP5b受體從而介導HDL顆粒的胞吞作用,APOA-I和這一受體的結合嚴格依賴于 ADP的產生[28],上調 ATP5b為增強HDL的胞吞作用提供動力。
2.2 熱應激影響畜禽生產性能相關的差異表達基因研究
2.2.1 肉品質
通過基因芯片分析熱應激下肉雞胸肌的差異表達基因有110個,其中包括F-框蛋白家族成員(FBXO11)、泛素B/C(UBB/UBC)、多聚腺苷酸結合蛋白1(PABPC1)、乙酰輔酶 A轉乙酰酶2(ACAT2)、長鏈脂肪酸延長酶2(ELOVL2)等67個上調基因,肌肉磷酸果糖激酶(PFKM)、LEPROT等43個下調基因,差異表達基因主要與細胞結構、信號傳導、蛋白質代謝和修飾等相關[5]。FBXO11在泛素介導的蛋白質水解過程中具有對底物識別的特性[29],泛素可指導蛋白質的細胞內吞并作為胞內蛋白質運輸體的組成部分[30],泛素-蛋白酶體蛋白質水解途徑可能對肌肉蛋白質的合成和肉的嫩度存在影響,當動物遭受嚴重應激時,肌肉蛋白質降解以提供肝、腎糖異生原料而導致肌肉萎縮,此時,骨骼肌組織中泛素-蛋白酶體通路表達上調以降解大量骨骼肌蛋白[31]。熱應激條件下,與泛素相關的基因,如UBB/UBC、PABPC1、FBXO11,均出現(xiàn)表達量增加[32],同時 Kuskwaha等[33]也進一步證實了泛素-蛋白酶體通路參與熱應激的調控。ACAT2是參于脂肪酸降解代謝的酶,ACAT表達上升,表明肝臟利用脂肪酸的能力加大,導致脂肪酸氧化代謝增強。ELOVL2編碼延長酶且參與長鏈多不飽和脂肪酸的生物合成,調控動物體內的脂肪構成或脂肪沉積[34]。瘦素(leptin,LP)是由脂肪細胞合成的基因產物[35],并通過抑制神經多肽mRNA的表達及分泌而影響采食量、導致體重下降[36],使下丘腦弓狀核中的LEPROT發(fā)生沉默并調動LP受體信號通路來控制采食量[37],熱應激也直接影響到了牛外周血中瘦素基因的合成,并降低LP受體及其異構型受體基因mRNA的表達[38]。PFKM 是糖酵解途徑中重要的調控因子,熱應激引起PFKM的下調導致肌肉攝取葡萄糖減少致使糖酵解降低[39],直接影響肌肉中氨基酸、脂質含量及脂肪酸的組成。
2.2.2 繁殖性能
熱應激損傷牛、羊、豬精子DNA的完整性,導致受精率明顯下降[40],嚴重影響動物繁殖力甚至致使不育是可遺傳的[41]。對熱應激下的雄性小鼠睪丸組織進行基因芯片分析,發(fā)現(xiàn)225個差異表達基因,其中包括分子伴侶鈣連接蛋白(CANX)、HSPcb1、T-復合多肽(TCP1)和催化反應物FK506連接蛋白6(FKPB6)、蛋白酶體亞基(PSMA7)、異檸檬酸脫氫酶1(IDH1)的差異表達,并在其子1、2代雄性小鼠睪丸組織中檢測到CANX、HSPcb1、TCP1的表達,其中 HSPcb1在子代穩(wěn)定地表達[40]。CANX過表達能糾正錯誤折疊蛋白質并延緩細胞凋亡,對潛在的凋亡過程起抑制作用[42];TCP1是一種復合蛋白質,在精子發(fā)生過程中其基因表達量增加[43];HSPcb屬于HSP90家族成員,是一種惰性、不穩(wěn)定的細胞基質蛋白質,上調HSPcb可以阻止錯誤折疊或未折疊蛋白質的組裝,HSP90的表達水平可直接影響精子的發(fā)生甚至不育[41]。Li等[44]利用基因芯片對高溫處理8 h后的C57BL/6小鼠與耐熱應激AKR/N小鼠睪丸組織的基因轉錄本表達分析,發(fā)生顯著上調的9個與熱休克蛋白相關以及4個與類固醇生物合成相關的差異表達基因,精子發(fā)生過程中HSPs分子伴侶的高度表達直接參與調控精母細胞減數(shù)分裂粗線期至精子形成[45]。此外,熱應激可引起一系列與生殖性能相關基因差異表達的變化,明顯上調的基因有類固醇激素合成急性調節(jié)蛋白(STAR)、膽固醇側鏈裂解酶(CYPLLAL)等,明顯下調的基因有精子頂體相關蛋白3(SPACA3)、精細胞核周期RNA結合蛋白(STRBP)等[44]。
2.3 熱應激影響體外細胞生長、凋亡相關的差異表達基因研究
急性熱應激可使體外培養(yǎng)下的細胞周期停滯在 G1/S、G2/M 期[46-47],并誘發(fā)細胞凋亡。利用表達譜基因芯片研究體外培養(yǎng)的奶牛乳腺上皮細胞在熱應激前后的基因表達差異,發(fā)現(xiàn)處理2 h后上調的基因主要與熱應答、DNA修復和蛋白質修復有關,下調的基因主要與細胞循環(huán)、代謝和結構蛋白有關;應激8 h后HSP70下降到基礎水平,熱耐受能力消失,與凋亡相關基因開始上調表達[48],與Soto等[49]的報道相似,40℃高溫應激可使凋亡基因(Bax)表達顯著上調,B細胞淋巴瘤2(Bcl-2)表達顯著下調。Bcl-2主要生物學功能是增加細胞對多種凋亡刺激因素的抵抗力來減少細胞凋亡,Bcl-2 和 Bax 的表達失衡會改變細胞中 Bcl-2/Bax異二聚體形成,產生 Bcl-2/Bcl-2 或 Bax/Bax 同二聚體,后者增多則會引起細胞線粒體通透性增加,引起線粒體蛋白細胞色素C釋放到胞質中,激活caspase-9,從 而 導 致 細 胞 凋 亡。p53 是 Bcl-2 和Bax的上游調節(jié)基因,許多凋亡基因啟動子都存在p53 反應元件(p53 responsive element,pRE),結合p53后被激活轉錄表達,如腫瘤壞死因子(TNF)受體家族Bax、脂肪酸合酶(Fas)及其配體(Fasl)等表達增加,Bcl-2表達受抑制。p53還能促進內質網(wǎng)釋放 TNF-α和 Fas,也能與 DNA復制蛋白 A(replication protein A,RPA)結合,阻止 DNA 復制,最終激活 caspase-7、6、3,促進細胞凋亡[50]。在熱應激誘導腫瘤細胞凋亡的研究中,發(fā)現(xiàn)核酸內切酶的活化可能依賴于胞內鈣離子介導Bax升高而增加線粒體膜通透性,促進鈣離子外流調節(jié)細胞凋亡的發(fā)生[51]。熱應激嚴重影響小鼠腸上皮細胞系-6(IEC-6)的培養(yǎng),引起一些生長因子的差異表達,如上調基因有轉分化生長因子(GDF)、血小板生長因子(PDGFA)、生長抑制因子(OK138)、血管生長因子(VEGFA)等,下調基因有白蛋白(ALB)、EGFR、成纖維細胞生長因子(FGF)等[3],而生長因子通過與細胞膜上相應配體結合,激活受體,使受體的酪氨酸激酶活性上升,催化細胞內多種蛋白質底物的酪氨酸殘基磷酸化,繼而促進DNA、RNA和蛋白質等大分子物質的生物合成,加速細胞的增殖和分化[52]。
3 小結
基因芯片技術是對基因組學、轉錄組學和生物信息學等系統(tǒng)生物學研究領域的應用,在畜禽生產中進行多種差異基因表達譜的研究起重要作用。通過基因芯片技術篩選畜禽熱應激差異表達基因,可以給出熱應激的分子表達譜與非熱應激以及應激損傷修復之間的差異基因,從而識別出熱敏感的特異性基因及熱損傷的特異性基因,有助于揭示熱應激發(fā)生與進展的分子機制;可以識別熱應激損傷特異性和修復特征相關的分子標志物,有助于開展利用熱應激分子特征進行緩解應激損傷修復的研究,進一步調控差異表達基因來應對熱應激對畜禽機體損傷,并顯示出巨大的潛力和廣泛的應用前景。此外,基因芯片技術能篩選出多條與熱應激相關的細胞信號轉導途徑,可通過調控異常表達基因的水平來深入研究與之相關的信號傳導通路,這對畜禽生產的基礎研究都是非常重要的。
[1]TREVINO V,F(xiàn)ALCIANI F,BARRERA-SALDANA H A.DNA microarrays:a powerful genomic tool for biomedical and clinical research[J].Molecular Medicine,2007,13(9/10):527 -541.
[2]KARAKAC T K,F(xiàn)LIGHT R M,DOUGLAS S E,et al.An introduction to DNA microarrays for gene expression analysis[J].Chemometrics and Intelligent Laboratory Systems,2010,104:28 -52.
[3]YU J,YIN P,LIU F,et al.Effect of heat stress on the porcine small intestine:a morphological and gene expression study[J].Comparative Biochemistry and Physiology-Part A:Molecular and Integrative Physiology,2010,156(1):119 -128.
[4]YU J,LIU F,YIN P,et al.Integrating miRNA and mRNA expression profiles in response to heat stress induced injury in rat small intestine[J].Functional and Integrative Genomics,2011,11(2):203 -213.
[5]LI C,WANG X,WANG G,et al.Expression analysis of global gene response to chronic heat exposure in broiler chickens(Gallus gallus)reveals new reactive genes[J].Poultry Science,2011,90(5):1028 -1036.
[6]MURAKAMI A E,SAKAMOTO M I,NATALI M R,et al.Supplementation of glutamine and vitamin E on the morphometry of the intestinal mucosa in broiler chickens[J].Poultry Science,2007,86(3):488 -495.
[7]LIU F,YIN J,DU M,et al.Heat stress induced damage to porcine small intestinal epithelium associated with down regulation of epithelial growth factor signaling[J].Journal of Animal Science,2009,87(6):1941-1949.
[8]YU J,YIN P,YIN J,et al.Involvement of ERK1/2 signalling and growth-related molecules expression in response to heat stress induced damage in rat jejunum and IEC-6 cells[J].International Journal of Hyperthermia,2010,26(6):538 -555.
[9]RUBIO N,SANZ-RODRIGUEZ F,LIPTON H L.Theiler’s virus induces the MIP-2 chemokine(CXCL2)in astrocytes from genetically susceptible but not from resistant mouse strains[J].Cellular Immunology,2006,239(1):31 -40.
[10]DORTS J,BAUWIN A,KESTEMONT P,et al.Proteasome and antioxidant responses in Cottus gobio during a combined exposure to heat stress and cadmium[J].Comparative Biochemistry and Physiology-Part C:Toxicology and Pharmacology,2012,155(2):318-324.
[11]LU A,WANG H C,HOU X L,et al.Microarray analysis of gene expression profiles of rat small intestine in response to heat stress[J].Journal of Biomolecular Screening,2011,16(6):655 -667.
[12]ADACHI H,KATSUNO M,WAZA M,et al.Heat shock proteins in neurodegenerative diseases:pathogenic roles and therapeutic implications[J].International Journal of Hyperthermia,2009,25(8):647 -654.
[13]ANTONINI D,RUSSO M T,ROSA L D,et al.Transcriptional repression of miR-34 family contributes to p63-mediated cell cycle progression in epidermal cells[J].Journal of Investigative Dermatology,2010,130(5):1249-1257.
[14]HEMEKING H.The miR-34 family in cancer and apoptosis[J].Cell Death and Differentiation,2010,17(2):193-199.
[15]CHOI Y J,LIN C P,HO J J,et al.miR-34 miRNAs provide a barrier for somatic cell reprogramming[J].Nature Cell Biology,2011,13(11):1353 -1360.
[16]DHARAP A,BOWEN K,PLACE R,et al.Transient focal ischemia induces extensive temporal changes in rat cerebral microRNAome[J].Journal of Cerebral Blood Flow and Metabolism,2009,29(4):675 -687.
[17]LEI P,LI Y,CHEN X,et al.Microarray based analysis of microRNA expression in rat cerebral cortex after traumatic brain injury[J].Brain Research,2009,1284:191-201.
[18]BHUSARI S,HEARNE L B,SPIERS D E,et al.Transcriptional profiling of mouse liver in response to chronic heat stress[J].Journal of Thermal Biology,2008,33(3):157 -167.
[19]OHTA S,OHSAWA I,KAMINO K,et al.Mitochondrial ALDH2 deficiency as an oxidative stress[J].Annals of the New York Academy of Sciences,2004,1011:36-44.
[20]CHIAVERINI N,DELEY M.Protective effect of metallothionein on oxidative stress-induced DNA damage[J].Free Radic Research,2010,44(6):605 -613.
[21]ZHANG H J,DRAKE V J,MORRISON J P,et al.Molecular biology of thermoregulation:selected contribution:differential expression of stress-related genes with aging and hyperthermia[J].Journal of Applied Physiology,2002,92(4):1762 -1769.
[22]SCHRINER S E,LINFORD N J,MARTIN G M,et al.Extension of murine life span by overexpression of catalase targeted to mitochondria[J].Science,2005,308(5730):1909-1911.
[23]SONCIN F,ZHANG X,CHU B,et al.Transcriptional activity and DNA binding of heat shock factor-1 involve phosphorylation on threonine 142 by CK2[J].Biochemical and Biophysical Research Communications,2003,303(2):700 -706.
[24]LOIZOU J I,EL-KHAMISY SF,ZLATANOU A,et al.The protein kinase CK2 facilitates repair of chromosomal DNA singlestrand breaks[J].Cell,2004,117(1):17-28.
[25]GONZALEZ F J.Role of cytochromes p450 in chemical toxicity and oxidative stress:studies with cyp2e1[J].Mutation Research-Fundamental and Molecular Mechanisms of Mutagenesis,2005,569(1/2):101 -110.
[26]KWAK M K,WAKABAYASHI N,GREENLAW J L,et al.Antioxidants enhance mammalian proteasome expression through the keap1-nrf2 signaling pathway[J].Molecular Biology of the Cell,2003,23(23):8786-8794.
[27]ECKARDSTEIN A V.Differential diagnosis of familial high density lipoprotein deficiency syndromes[J].Atherosclerosis,2006,186(2):231 -239.
[28]MARTINEZ L O,JACQUET S,ESTEVE J P,et al.Ectopic beta-chain of ATP synthase is an apolipoprotein A-I receptor in hepatic HDL endocytosis[J].Nature,2003,421(6918):75 -79.
[29]ABIDA W M,NIKOLAEV A,ZHAO W,et al.FBXO11promotes the Neddylation of p53 and inhibits its transcriptional activity[J].The Journal of Biological Chemistry,2007,282(3):1797 -1804.
[30]AGUILAR R C,WENDLAND B.Ubiquitin not just for proteasomes anymore[J].Current Opinion in Cell Biology,2003,15(2):184 -190.
[31]MURTON A J,CONSTANTIN D,GREENHAFF P L,et al.The involvement of the ubiquitin proteasome system in human skeletal muscle remodelling and atrophy[J].Biochimica et Biophysica Acta(BBA)-Molecular Basis of Disease,2008,1782(12):730 -743.
[32]BUCKLEY B A,GRACEY A Y,SOMERO G N.The cellular response to heat stress in the goby Gillichthys mirabilis:a cDNA microarray and protein-level analysis[J].Journal of Experimental Biology,2006,209(14):2660-2677.
[33]KUSKWAHA R S,ROSILLO A,RODRIGUEZ R,et al.Expression levels of ACAT1 and ACAT2 genes in the liver and intestine of baboons with high and low lipemic responses to dietary lipids[J].Journal of Nutritional Biochemistry,2005,16(12):714 -721.
[34]GUILLOU H,ZADRAVEC D,MARTIN P G,et al.The key roles of elongases and desaturases in mammalian fatty acid metabolism:insights from transgenic mice[J].Progress in Lipid Research,2010,49(2):186-199.
[35]ZHANG Y,PROENZA R,MAFFEI M.Positional cloning of the mouse obese gene and its human homologue[J].Nature,1994,372(6505):425 - 432.
[36]PICO C,JILKOVA Z M,KUS V,et al.Perinatal programming of body weight control by leptin:putative roles of AMP kinase and muscle thermogenesis[J].American Journal of Clinical Nutrition,2011,94(6):1830-1837.
[37]COUTURIER C,SARKIS C,SERON K,et al.Silencing of OB-RGRP in mouse hypothalamic arcuate nucleus increases leptin receptor signaling and prevents diet-induced obesity[J].Proceedings of the National Academy of Sciences of the United States of Ameri-ca,2007,104(49):19476 -19481.
[38]LACETERA N,BERNABUCCI U,BASIRICO L,et al.Heat shock impairs DNA synthesis and down-regulates gene expression for leptin and Ob-Rb receptor in concanavalin A-stimulated bovine peripheral blood mononuclear cells[J].Veterinary Immunology and Immunopathology,2009,127(1/2):190 -204.
[39]DERVISEVIK M,DINEVSKA-KJOVKAROVSKA S,MIOVA B,et al.Heat acclimation-induced changes in heart glycogen/glucose metabolism in rats[J].Journal of Physiology Science,2011,61(5):395 -402.
[40]CAMMACK K M,ANTONIOU E,HEARNE L,et al.Testicular gene expression in male mice divergent for fertility after heat stress[J].Theriogenology,2009,71(4):651 -661.
[41]CAMMACK K M,MESA H,LAMBERSON W R.Genetic variation in fertility of heat-stressed male mice[J].Theriogenology,2006,66(9):2195 -2201.
[42]TAKIZAWA T,TATEMATSU K,WATANABE K,et al.Cleavage of calnexin caused by apoptotic stimuli:implication for the regulation of apotosis[J].Journal of Biochemistry,2004,136(3):399 -405.
[43]LI E,GUO Y,NING Q,et al.Research for the effect of octylphenol on spermatogenesis and proteomic analysis in octylphenol-treated mice testes[J].Cell Biology International,2011,35(4):305 - 309.
[44]LI Y,ZHOU Q,HIVELY R,et al.Differential gene expression in the testes of different murine strains under normal and hyperthermic conditions[J].Journal of Andrology,2009,30(3):325 -337.
[45]VYDRA N,MALUSECKA E,JARZAB M,et al.Spermatocyte-specific expression of constitutively active heat shock factor 1 induces HSP70i-resistant apoptosis in male germ cells[J].Cell Death and Differentiation,2006,13(2):212 -222.
[46]LI X,CAI M.Recovery of the yeast cell cycle from heat shock-induced Gl arrest involves a positive regulation of G1 cyclin expression by the S phase cyclin Clb5[J].Journal of Biological Chemistry,1999,274(34):24220-24231.
[47]KUHL N M,RENSING L.Heat shock effects on cell cycle progression[J].Cellular and Molecular Life Sciences,2000,57(3):450 -463.
[48]COLLIER R J,STIENING C M,POLLARD B C.Use of gene expression microarrays for evaluating environmental stress tolerance at the cellular level in cattle[J].Journal of Animal Science,2006,84:1 - 13.
[49]SOTO P,SMITH L C.BH4 peptide derived from BclxL and Bax-inhibitor peptide suppresses apoptotic mitochondrial changes in heat stressed bovine oocytes[J].MolecularReproduction and Development,2009,76(7):637 -646.
[50]KUROKAWA M,KORNBLUTH S.Caspases and kinases in a death grip[J].Cell,2009,139(5)838 -854.
[51]NICHOISON D W,THOMBERRY N A.Apoptosis:life and death decision[J].Science,2003,299(5604):214-215.
[52]GREWAL T,ENRIEH C.Annexins-modulators of EGF receptor signalling and trafficking[J].Cellular Signalling,2009,21(6):847 -858.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語文(2016年21期)2016-05-25 13:13:50
