GD3 synthase與肝癌的研究進展
2012-09-05 09:46:09余勇張玉勝
海南醫學 2012年2期
余勇,張玉勝
(廣東醫學院,廣東湛江523808)
GD3 synthase與肝癌的研究進展
余勇,張玉勝
(廣東醫學院,廣東湛江523808)
GD3是含唾液酸的神經節糖脂分子,與細胞的多種功能密切相關,在這個過程中包含了細胞凋亡與增殖、細胞分化、信號傳導和細胞遷徙等一系列生物行為。本文主要綜述了GD3的分子結構,生物學作用,尤其是在肝癌的發生與發展過程的作用及臨床研究成果。
GD3;肝癌;NF-KB
脂質和糖脂類介質是細胞凋亡過程中重要的信號分子。神經節苷脂類是唾液酸糖苷神經鞘脂類的一個家族,是脊椎動物細胞質膜外葉的組成部分,在生物多樣性的進程中有著不可或缺的作用,如細胞間的相互作用、粘附、細胞分化、生長調控、受體的功能等等[1-2]。在腫瘤的形成和進展中也是很重要的[3],在神經外胚葉性腫瘤細胞、黑色素瘤細胞、成神經瘤細胞中都有表達[4-6]。GD3是屬于神經節苷脂類,即唾液酸糖苷神經鞘脂類,又稱α-2,8-唾液酸基轉移酶。其是B族神經苷脂類合成的關鍵酶,表達是在細胞內ST8Sia I基因水平調節的[7]。GD3不僅是重要的腫瘤標記物,也是腫瘤靶向治療的一個靶點。盡管GD3未定義為癌基因,但是GD3蛋白的表達量增高在多種組織起源的腫瘤中都有報道。近年來發現GD3與肝癌關系密切,其與肝癌關系是促進還是抑制頗具爭議。GD3與腫瘤關系的報道較少,其與肝癌的進展及預后的關系已經吸引了學術界的眼球。
1 GD3的結構和生物學功能
GD3是一種含唾液酸的神經鞘糖脂。神經節苷脂(Ganglioside,Gg)按唾液酸的殘疾數目有GM、GD、GT、GQ和GP五個家族。Gg是兩性分子,既有脂溶性又有水溶性,分子量在1~3 kD。在水溶液里,由許多Gg分子組成球形聚合體,其糖鏈部分朝外,烴鏈部分朝內,每個聚合體的分子量約為300 kD。其是脊椎動物細胞質膜外葉的組成部分,在大腦灰質中含量最高,此外高爾基體、內質網和溶酶體中也含有。Gg有兩個基本功能:(1)介導細胞—細胞,細胞—微生物以及細胞與基質間的相互作用。(2)調控細胞質膜中蛋白質的功能,如生長因子受體、離子通道等的功能。外源性的Gg可以通過與細胞表面的Gg結合蛋白結合,來改變與Gg相連的激酶活性從而增高或減低細胞生長速率[8]。而且Gg可以作為細胞的特定標志而被識別,對細胞的粘附也起著重要的作用。粘附分子ELAM-1在急性炎癥時可以介導白細胞對內皮細胞的粘附。在這一過程中Gg的唾液酸殘基是必需的,ELAM-1只有識別細胞上的Gg唾液酸殘基才能發揮其粘附作用。
眾所周知,線粒體、內質網、高爾基復合體等亞細胞器參與了細胞程序性凋亡,近年的研究揭示了FAS相關蛋白介導的細胞凋亡是通過改變線粒體等細胞器的膜通透性,從而引起線粒體功能紊亂,而這些是通過調節Caspases來實現的。脂筏是由鞘脂類組成,包括神經節苷脂、神經鞘磷脂、膽固醇和信號蛋白,這就提示它們參與了介導線粒體的功能紊亂并且參與了細胞的重塑,引起細胞程序性凋亡[9]。目前國內外對細胞程序性凋亡的研究主要在通過介導CD95/Fas的T細胞性凋亡。氧自由基、p53及Bax等產物的增加激活T細胞,Sa等[10]研究闡述了GD3調解活性T細胞介導的細胞凋亡。改變線粒體通透性,刺激細胞色素C釋放以及激活Caspase-9都與GD3有關聯。有文獻報道稱GD3能夠介導細胞增殖和分化等生物行為,通過改變細胞水平GD3的定位及結構來影響細胞的生物學行為[11]。在線粒體內,細胞凋亡前體蛋白間的相互作用激活了凋亡途徑,GD3作為一種新出現的細胞凋亡脂質充當介質,在線粒體內負責凋亡信號的發出,García-Ruiz等[12]用激光掃描共焦顯微鏡及免疫電子技術證明了在鼠肝細胞中TNF-α能將GD3轉移至線粒體內,GD3通過唾液酸糖蛋白受體共區域化在肝細胞的高爾基復合體和質膜中有明顯的定位。
2 GD3與腫瘤的關系
神經節苷脂類的研究在國內主要局限于神經系統,在神經傳遞和神經受體功能方面其重要作用,對體外培養的神經細胞有營養和促進作用,有研究指出神經節苷脂類能減輕實驗動物神經損傷和缺血后發生的腦水腫,并加速神經功能的回復。神經節普脂廣泛分布于神經系統,其濃度及組成因腦、脊柱、末梢神經等部位及細胞種類之不同而異。從動物發育中的中樞神經系統神經節普脂變化的研究中,作為分化抗原標志的神經節普脂已被重視起來。乳糖系神經節苷脂中的唾液酸—Le(CA-19-9)的單克隆抗體(N-19-9)在結腸癌,胰腺癌,胃癌的患者血清中均出現,診斷價值較高。另一種叫做SSEA-3(Stage-Specific embryonic antigen)的神經節苷脂已經在畸胎瘤中發現。GD3作為一種小分子神經節苷脂,在一些病理學條件下都有過表達,比如癌癥及神經性紊亂。GD3在人類腫瘤,如黑色素瘤、腦癌、前列腺癌、乳腺癌、白血病、肝癌中都有表達,其在這些組織中的表達呈現的作用不盡相同。目前國內外報道最多的就是GD3在黑色素瘤細胞中的表達,Kang等[13]闡述了GD3在人黑色素瘤細胞中的過表達,并證實了GD3基因表達的啟動區,而且通過實驗提示了NF-KB在黑色素瘤細胞中表達GD3的基因轉錄激活方面有很重要的作用。且Kwon等[14]研究表明雷公藤內酯能夠下調GD3在黑色素瘤細胞中的表達。
目前對GD3的研究主要是在神經損傷方面,有關GD3在腫瘤發生過程中的作用相關報道較少。Fabbri等[15]發現用蒽環類藥物作用的前列腺癌細胞中有GD3的表達,并進一步證明了GD3發揮抗癌的作用是通過高爾基體而發揮作用的。2009年Cazet等[16]發現在乳腺癌中有GD3的過表達,在浸潤性導管癌中大約50%有過表達,并通過GD3、GD2、GT3的對照研究指出GD3可能與癌細胞的惡性程度有關,但是其與乳腺癌的進展關系不大。2008年Mandal等[17]發現在早期急性淋巴細胞白血病中GD3的表達水平較高。
3 GD3與肝癌的關系
GD3作為一種腫瘤標記物,近年來國內外學者對GD3在肝癌組織中的表達進行了一些研究。研究中有實驗結果提示GD3在肝癌細胞中的表達要高于癌旁組織和正常肝癌組織,有的則持相反論斷,但有一點是肯定的,那就是GD3與肝癌有著密切的聯系。2002年Paris等[18]對GD3在肝癌中的作用做了一些研究,研究證明GD3能夠通過阻斷核因子KB (NF-KB)途徑進而達到肝癌治療的目的,但是在組織表達方面卻存在不確定性。國內的一些學者研究闡述了GD3在肝癌組織中的過表達,并提出了GD3的過表達可能與GD3合成酶活性增高有關,還與其水解酶唾液酸酶活性的降低有關[19]。GD3是細胞凋亡的影響因子,有報道稱CD95和N-脂酰鞘氨醇誘導的肝細胞凋亡,這些細胞中的GD3有過表達[20]。有報道指出了GD3與肝癌發生與發展的關系,2007年Chu等[21]的研究指出肝細胞生長因子是重要的細胞因子,能引起多種癌細胞的轉移和侵襲,且能導致血管再生。人神經膠質瘤細胞中的GD3能刺激人類生長因子(HGF)的分泌,這種作用與轉移性生長因子(TGF)-β相似。這說明GD3可能是一種間接的血管生成因子。
Mesari?等[22]闡述了GD3的激活與肝癌有關聯,并通過動物實驗證明了在小鼠的肝細胞中GD3的激活有賴于性激素,但是目前臨床報道較少。但是也有些文獻指出活性氧介導的GD3能夠抑制細胞外信號調節激酶和基質金屬蛋白酶(MMP-9)的分泌,實驗進一步證實了GD3改變血管平滑肌細胞(VSMC)的表型的過程中氧自由基起著重要的作用[23],這就說明GD3能夠影響細胞間的粘附作用,減少腫瘤血管的生成。如圖1所示,GD3生物作用發揮與NF-KB/cScr通道有關[24]。組織缺氧誘導分泌低氧誘導因子1(HIF-1)和NF-KB的激活在癌癥發生和進展,血管發生及腫瘤侵襲方面都有影響[24]。缺氧為環境能誘導細胞凋亡,這可能與GD3誘導HIF-1表達上調有關。這些研究說明了GD3在肝癌的發生發展中有抑制作用。
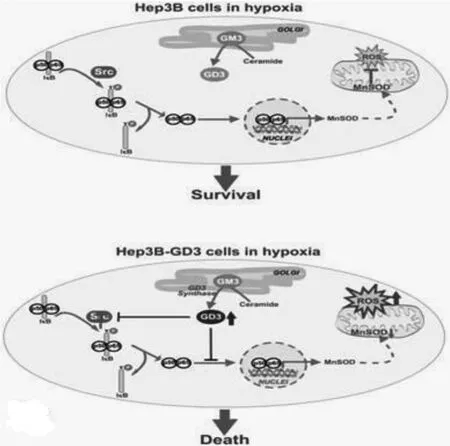
圖1 GD3生物作用途徑
4 GD3與肝癌浸潤與轉移
浸潤與遠處轉移是腫瘤細胞重要的生物學特征之一,肝癌患者的死亡大多與之有關。臨床上大多數腫瘤患者不是死于原發灶,而是死于不同程度的腫瘤轉移。在一個腫瘤細胞群體中,細胞不完全相同,其生物學特性有不完全相同,這就是所謂的異質性。據此認為癌的轉移并不是所有的癌細胞具有的特性,而是腫瘤細胞群體中一些具有轉移能力的癌細胞所致。其轉移過程包含一系列連續的步驟,引起轉移性生長的瘤細胞,首先必須離開原發瘤并侵入周圍組織,及瘤細胞的粘附性降低而活動性增高,最后這些細胞進入血液或淋巴系統,并由此播散全身。GD3與VSMC的關系暗示了GD3在腫瘤侵潤方面有一定的抑制作用。但是此類報道并不多,因此GD3在肝癌組織中的表達規律及其在肝癌發生過程中的作用有待進一步的研究證實。
5 小結
GD3在許多細胞生物行為包括細胞分化,細胞凋亡和信號傳導中都起重要作用。在一些腫瘤細胞中GD3有很重要的臨床作用。在黑色素瘤中,GD3的過表達可以作為獨立診斷標準。但是GD3在肝癌中的作用并不是很清晰,有待深入研究。
[1]Svennerholm L.Gangliosides and synaptic transmission[J].Adv Exp Med Biol,1980,125:533-544.
[2]Hakomori S,Igarashi Y.Gangliosides and glycosphingolipids as modulators of cell growth,adhesion,and transmembrane signaling [J].Adv Lipid Res,1993,25:147-162.
[3]Hakomori S.Tumor malignancy defined by aberrant glycosylation and sphingo(glyco)lipid metabolism[J].Cancer Res,1996,56: 5309-5318.
[4]Hakomori S.Glycosphingolipids in cellular interaction,differentiation,and oncogenesis[J].Annu Rev Biochem,1981,50:733-764.
[5]Portoukalian J,Zwingelstein G,Dore JF.Lipid composition of human malignant melanoma tumors at various levels of malignant growth[J].Eur J Biochem,1979,94:19-23.
[6]Cheung NK,Saarinen UM,Neely JE,et al.Monoclonal antibodies to a glycolipid antigen on human neuroblastoma cells[J].Cancer Res,1985,45:2642-2649.
[7]Kim JK,Kim SH,Cho HY,et al.GD3 accumulation in cell surface lipid rafts prior to mitochondrial targeting contributes to amyloid-β-induced apoptosis[J].Korean Med Sci,2010,25(10): 1492-1498.
[8]Schengrund CL.The role(s)of gangliosides in neural differentiation and repair:a perspective[J].Brain Res Bull,1990,24(1):131-141.
[9]Sorice M,Garofalo T,Misasi R,et al.Ganglioside GD3 as a RAFT component in cell death regulation[J].Anticancer Agents Med Chem,2011,[Epub ahead of print]
[10]Sa G,Das T,Moon C,et al.GD3,an overexpressed tumor-derived ganglioside,mediates the apoptosis of activated but not resting T cells[J].Cancer Res.2009,69(7):3095-3104.
[11]Malisan F,Testi R.GD3 ganglioside and apoptosis[J].Biochim BiophysActa,2002,1585(2-3):179-187.
[12]García-Ruiz C,Colell A,Morales A,et al.Trafficking of ganglioside GD3 to mitochondria by tumor necrosis factor-alpha[J].J Biol Chem,2002,277(39):36443-36448.
[13]Kang NY,Kim CH,Kim KS,et al.Expression of the human CMP-NeuAc:GM3 α 2,8 sialyltransferase(GD3 synthase)gene through the NF-κB activation in human melanoma SK-MEL-2 cells [J].Biochim BiophysActa,2007,1769:622-630.
[14]Kwon HY,Kim SJ,Kim CH,et al.Triptolide downregulates human GD3 synthase(hST8Sia I)gene expression in SK-MEL-2 human melanoma cells[J].Exp Mol Med,2010,42(12):849-855.
[15]Fabbri F,Zoli W,Carloni S,et al.Activity of different anthracycline formulations in hormone-refractory prostate cancer cell lines:role of golgi apparatus[J].J Cell Physiol,2011,doi:10.1002/jcp.22654.
[16]Cazet A,Groux-Degroote S,Teylaert B,et al.GD3 synthase overexpression enhances proliferation and migration of MDA-MB-231 breast cancer cells[J].Biol Chem,2009,390(7):601-609.
[17]Mandal C,Srinivasan GV,Chowdhury S,et al.High level of sialate-O-acetyltransferase activity in lymphoblasts of childhood acute lymphoblastic leukaemia(ALL):enzyme characterization and correlation with disease status[J].Glycoconj J,2009,26(1):57-73.
[18]Paris R,Morales A,Coll O,et al.Ganglioside GD3 sensitizes human hepatoma cells to cancer therapy[J].J Biol Chem,2002,277 (51):49870-49876.
[19]謝天培,沈鋒,施樂華.肝癌組織中神經節苷脂譜改變的機理研究[J].上海醫科大學學報,1996,17(3):213-215.
[20]Kim JK,Kim SH,Cho HY,et al.GD3 accumulation in cell surface lipid rafts prior to mitochondrial targeting contributes to amyloid-β-induced apoptosis[J].J Korean Med Sci,2010,25(10): 1492-1498.
[21]Chu SH,Ma YB,Zhang H,et al.Hepatocyte growth factor production is stimulated by gangliosides and TGF-beta isoforms in human glioma cells[J].J Neurooncol,2007,85(1):33-38.
[22]Mesari? M,Decker K.Activation of GD3 synthase by sex steroid hormones in cultured rat hepatocytes[J].Biochem Biophys Res Commun,1990,28,171(3):1188-1191.
[23]Moon SK,Kang SK,Kim CH.Reactive oxygen species mediates disialoganglioside GD3-induced inhibition of ERK1/2 and matrix metalloproteinase-9 expression in vascular smooth muscle cells[J]. FASEB J,2006,20(9):1387-1395.
[24]Lluis JM,Llacuna L,von Montfort C,et al.GD3 synthase overexpression sensitizes hepatocarcinoma cells to hypoxia and reduces tumor growth by suppressing the cSrc/NF-kappaB survival pathway [J].PLoS One,2009,4(11):e8059.
[25]Cheng J,Yu H,Lau K,et al.Multifunctionality of Campylobacter jejuni sialyltransferase CstⅡ:characterization of GD3/GT3oligosaccharide synthase,GD3 oligosaccharide sialidase,and trans-sialidase activities[J].Glycobiology,2008,18(9):686-697.
Advances in the research of GD3 synthase and hepatic carcinoma.
YU Yong.Guangdong Medical College, Zhanjiang 523808,Guangdong,CHINA
The α 2,8-sialyl-transferase,called GD3 synthase,sialic acid-containing glycosphingolipids,has been historically considered to be involved in the proliferation,differentiation,signal transduction and migration of biological behavior in vertebrates.This review summarizes the molecular structure and biological function of GD3 synthase,especially its function in the genesis and progress of hepatic carcinoma.
GD3 synthase;Hepatic carcinoma;NF-KB
R735.7
A
1003—6350(2012)02—122—03
10.3969/j.issn.1003-6350.2012.02.055
2011-08-04)
余勇(1985—),男,湖北省黃岡市人,在讀碩士。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
中成藥(2016年8期)2016-05-17 06:08:14
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
