常溫下CANON反應器中功能微生物的沿程分布
2012-09-04 02:30:14曾輝平
哈爾濱工業大學學報 2012年10期
劉 濤,李 冬,曾輝平,張 杰,
(1.哈爾濱工業大學城市水資源與水環境國家重點實驗室,150090哈爾濱;2.北京工業大學水質科學與水環境恢復工程北京市重點實驗室,100124北京)
由于上向流曝氣生物濾池具有處理效率高、占地面積小、抗沖擊負荷能力強等特點[1-2],目前許多CANON工藝多采用此形式實現.然而,這種反應器作為一種推流式反應器,由于濾池內部自下而上各段NH4+-N負荷及其他水力條件不同,會造成濾層各段微生物的活性及群落結構不同,從而會出現反應器沿程各段對NH4+-N的去除能力不同[3].因此,需要分析內部微生物的沿程分布規律,從而優化工藝運行條件.然而,目前這種類型CANON反應器內部的微生物沿程分布研究還鮮見報道[4-5].由于CANON反應器中的兩種功能細菌為氨氧化細菌(AOB)與厭氧氨氧化菌(ANAMMOX菌)[6],相比于分離、純化等傳統的生物學方法,應用顯微技術及分子生物技術研究兩類功能微生物的形態特征、群落結構及種屬特性,能夠更有效、更準確地獲得微生物方面的信息.掃描電鏡(SEM)雖然不能對微生物種群進行直接定性,但可對系統內微生物形態特征的變化進行直觀的評價.此外,在CANON工藝中,NH4+-N在氨單加氧酶(AMO)的催化作用下氧化生成羥胺(NH2OH),這是脫氮反應的第一步,也是極為關鍵的一步[7],amoA基因正是編碼氨單加氧酶(AMO)活性位點多肽的基因[8-9].利用amoA基因作為分子標記可以從分子水平研究氨氧化細菌的多樣性及種屬特性[10].對于厭氧氨氧化微生物來說,應用其特異性引物擴增16SrDNA作為標記可以研究其種群結構[11].
本文研究上向流曝氣生物濾池型CANON反應器中功能微生物種群結構及種屬特性的沿程變化規律,以便更合理地設置功能分區、優化設計參數,提高脫氮效果.
1 實驗
1.1 CANON反應器及樣品采集
上向流曝氣生物濾池型CANON反應器如圖1所示.反應器內徑150mm,有效高度700mm,總容積8.15L,有效體積1.8L,柱內裝填火山巖活性生物陶粒濾料.以NaHCO3、(NH4)2SO4、KH2PO4與自來水配制進水氨氮為300mg/L左右原水,從反應器底部進入,由上部出水口排出,在整個過程中,溫度一直保持在16~20℃.

圖1 CANON反應器試驗裝置及工藝流程圖
在反應器連續穩定運行第28天,分別從反應器濾層由上至下100、300、500和700 mm處采集濾料若干,將樣品保存于-20℃冰箱中用以提取基因組DNA.
1.2 掃描電鏡觀察
取不同濾層高度采集的濾料少許,經戊二醛固定、乙醇梯度脫水、乙酸異戊酯置換、臨界點干燥、離子濺射噴金處理后使用日立S-4300型掃描電鏡儀對樣品進行觀察并拍照.每個樣品隨機拍攝5~10張照片.
1.3 基因組DNA的提取
用無菌玻璃棒攪拌濾料以使生物膜脫落,稱取1g濕質量的生物膜,加入2.7mLDNA提取液(100mmol/LTris-HCl,100mmol/LEDTA(乙二胺四乙酸),1.5mol/LNaCl,100mmol/LNa3PO3,1%CTAB(十六烷基三甲基溴化銨),pH8.0),并加入50μL蛋白酶K(30g/L),50μL溶菌酶(20g/L)以及數粒玻璃珠,37℃水浴30min.此后加入1.5mLSDS(十二烷基硫酸鈉)溶液(200g/L),65℃水溶2h,期間每隔20min上下顛倒混勻一次.8000g離心10min,將上清液轉移至新的無菌離心管中,并加入等體積的氯仿/異戊醇(24∶1),混勻,8000g離心10min后將上清液轉移至新的無菌離心管中,并加入0.6倍體積預冷的異丙醇,-20℃過夜保存.12000g離心5min,棄上清,再以同樣的轉速離心3min,棄上清液并將樣品置于通風處徹底晾干,之后用50μL1×TEbuffer(10mmol/LTris-HCl;1mmol/LEDTA,pH8.0)溶解.所提取的基因組DNA結果用0.8%(質量分數)的瓊脂糖凝膠電泳檢測以備PCR用.
1.4 PCR擴增及DGGE電泳
用引物amoA-1F和amoA-2R[12]擴增氨氧化細菌amoA基因.其中amoA-1F的5'端所加GC夾子為DGGE設計;對于ANAMMOX菌的特異性片段的擴增采用巢式PCR方法:第一階段先用細菌的通用引物27F/1492R[13]進行16SrDNA序列的PCR擴增,并對PCR產物進行純化回收,之后以第一階段的PCR產物為模板,以ANAMMOX菌的特異性引物對Amx368F/Amx820[14]進行第二階段的PCR擴增,Amx368F的5'端加GC夾子同樣為后續DGGE所設計.PCR反應體系為25μL,其中包含2.5μL10×ExTaqbuffer(Mg2+Plus),dNTP2.0μL,引物各1.0μL,TaKaRaExTaq酶0.625U,模板DNA約1.0ng,用無菌水補齊至25μL.各種引物序列及PCR反應條件見文獻[12-14].PCR擴增產物用1.5%(質量分數)的瓊脂糖凝膠進行電泳檢測.
采用北京天根公司DNA膠回收試劑盒進行PCR產物的純化回收,具體操作按說明書進行.對PCR產物進行DGGE分析:聚丙烯酰胺質量分數8%,變性梯度為30%~60%,電壓120V,電泳時間5h,PCR產物上樣量約500ng,電泳在Dcode UniversalMutation Detection System儀器上進行.電泳結束后用Bassam等[15]的方法對凝膠進行銀染,并對凝膠拍照.
1.5 基因文庫的構建、測序及系統發育分析
切取DGGE圖譜中的目的條帶溶于50μLTE(pH8.0)溶液中,4℃過夜,以此為模板,以不含GC夾的引物進行PCR擴增,并對PCR產物進行純化.按照pMD19-Tplasmidvectorsystem說明書進行基因片段與載體的連接后,轉化到大腸桿菌DH5α感受態細胞中,通過藍白斑法篩選陽性克隆子并進行測序.采用BLAST對測序結果和基因庫中已知序列進行相似性分析,并利用MEGA4.0軟件,采用鄰位相連法(Neighbor-Joining)構建系統發育樹,自舉值為1000.
2 結果與討論
2.1 穩定運行時的脫氮效果
反應器在常溫、進水氨氮為300 mg/L條件下,通過調節曝氣及水力停留時間實現了CANON的穩定運行,并連續穩定運行約30 d.取樣時反應器運行工況為:進水氨氮質量濃度300 mg/L,溫度18℃,曝氣量4.5 L/min,氨氮去除率達83%,氨氮去除負荷為1.4kg/(m3·d),總氮去除率75%,總氮去除負荷1.1kg/(m3·d),出水硝氮濃度維持在20~35mg/L.盡管得到的總氮去除負荷略低于Sliekers以及Chuang等的研究[16-17],但依然屬于較高的去除負荷,因此,分析在該運行工況下的微生物沿程分布特點更具有代表性.
2.2 電鏡(SEM)照片
由于氨氧化細菌和厭氧氨氧化細菌形態多樣,一般難以通過形態來區分和鑒定其種屬.污水處理廠經常出現的氨氧化細菌主要是亞硝化球菌屬(Nitrosococcus)和亞硝化單胞菌屬(Nitrosomonas),其形態分別呈球狀和短桿狀,而亞硝酸鹽氧化菌主要是硝化螺菌屬(Nitrospira)和硝化桿菌屬(Nitrobacter),形態分別呈螺旋狀和桿狀[18].過去曾報導厭氧氨氧化菌為規則或者不規則的球形和橢球形,單生或成簇聚生[19],直徑約0.8~1.1 μm.因此,圖2中的球菌和橢球菌可能為亞硝化球菌屬(Nitrosococcus)和厭氧氨氧化細菌,短桿菌可能為亞硝化單胞菌屬(Nitrosomonas),而長桿菌可能為硝化桿菌屬(Nitrobacter).
由圖2可見,顯微鏡下可檢測到長桿菌、短桿菌、球菌和橢球形的菌,其中直徑0.2~1.0μm的橢球形和球形菌為優勢菌,幾乎未檢測到螺旋狀細菌,而長桿菌所占的比例也很低,說明反應器中亞硝酸鹽氧化菌含量很低,這也保證了反應器中的亞氮幾乎不會被硝酸鹽氧化菌利用,從而得到有效積累,為后續的厭氧氨氧化創造條件.圖2(a)中微生物數量較少,多數為單生,并未成簇聚生;在濾層300mm處,細菌數量增加,且出現了聚集生長的趨勢(圖2(b));隨著濾層深度的增加,微生物數量明顯增多(圖2(c)、(d)),且成簇聚生,其中以橢球形和球形菌為主,但也存在數量可觀的短桿菌.這些細菌可能為氨氧化細菌和厭氧氨氧化菌,其種屬特性將通過接下來的分子生物學技術進一步驗證.

圖2 不同濾層高度樣品的電鏡照片
2.3 PCR-DGGE圖譜及系統發育分析
由于PCR-DGGE圖譜中的每一條帶代表一個可能的細菌類群或可操作分類單位(OTU),條帶的數量和信號強度與生物多樣性和生物數量密切相關,基于PCR-DGGE圖譜可以確定不同取樣處微生物的種類和數量關系.
不同高度采集樣品的DGGE分析見圖3.氨氧化細菌在100和300mm處的可見條帶只有3條,而在500和700mm處條帶數量明顯增多,條帶信號明顯增強,說明氨氧化細菌在濾層500mm處以下的種類和數量明顯增多,這與掃描電鏡試驗的結果一致.其原因可能為:由于反應器采用上向流曝氣,而氨氧化細菌是一種嚴格的好氧細菌,在反應器底部曝氣量充足,有利于其生長.而富含氨氮基質的進水也是從反應器底部進入,氨氮會率先供給下部的氨氧化細菌.隨著反應器內溶解氧和氨氮被利用,在反應器上部的氨氧化細菌受到抑制,從而造成種類和數量的下降.此外,水力沖刷作用導致上部生物膜在一定程度上的破壞也是造成這種現象的原因之一.Purkhold等研究表明:污水處理系統中氨氧化細菌的多樣性程度越高,對復雜環境的適應能力就越強,其抗沖擊能力也就越強;反之,如果系統中只含有單一的氨氧化細菌,其抗干擾能力就較差[10].在本實驗所用的CANON反應器中,500 mm以下處的氨氧化細菌的多樣性程度很高,具有較強的抗沖擊負荷.然而在300mm以上的位置,氨氧化細菌多樣性很低,抗干擾能力較差.因此,為了提高濾層上部區域的抗沖擊負荷,需要采取一定措施提高氨氧化細菌的多樣性程度,其中最直接的做法就是對濾料進行重新排布.考慮到火山巖生物陶粒濾料填充的CANON反應器內已經形成穩定的氣道,重新排布濾料可能會破壞系統內的好氧/厭氧區域分布,進而破壞功能微生物的穩定性,影響系統的脫氮性能,可改用便于排布的軟性填料,比如海綿、無紡布等.但是它們對細菌的持留能力可能低于火山巖生物陶粒濾料,從而使CANON的啟動時間延長,具體解決方案還需進一步研究.
對于ANAMMOX菌來說,4個取樣點的DGGE圖譜基本一致,而且只有兩條可見條帶,說明ANAMMOX菌的群落結構在整個反應器中基本一致,幾乎不隨濾層高度的變化而變化.此外,條帶6的信號沿濾層自上而下有逐漸增強的趨勢,也說明ANAMMOX菌在反應器下方的數量要略多于上方.原因可能在于反應器上部氨氧化細菌種類和數量的減少,不能為ANAMMOX菌很好地創造厭氧環境,也不能提供ANAMMOX菌代謝所需足夠的亞硝酸鹽.此外,水力沖刷作用導致上部生物膜一定程度的破壞也是造成這種現象的原因之一.值得注意的是,通過條帶信號的強弱只能粗略推測細菌數量的多少,要想更精確地檢測細菌數量,還需要通過熒光定量PCR等其他檢測手段.
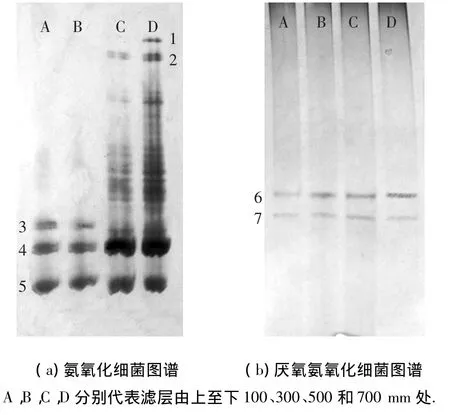
圖3 PCR產物DGGE圖譜沿程分布
對DGGE圖譜上的7條主要條帶進行切割、DNA洗脫、回收、重新擴增,構建基因克隆文庫,經測序所得的DNA序列提交至GenBank,得到的GenBank序列號為JN367453-JN367457以及JQ753318.對測序結果和基因庫中已知序列進行相似性對比分析,結果見表1.由于條帶6和7之間相似度達98%,可以歸并為一個可操作分類單位(OTU),它們與Candidatus Kuenenia stuttgariensis相似度達98%.這些已知細菌的形態多為球形、橢球形和短桿狀,與前文掃描電鏡結果一致.

表1 7個條帶所代表的基因序列對比結果
基于amoA基因序列構建系統發育樹(圖4),樹圖的外源基因為4種常見的氨氧化微生物,即亞硝化單胞菌屬(Nitrosomonas)、亞硝化螺菌屬(Nitrosospira)、亞硝化球菌屬(Nitrosococcus)、亞硝化葉菌屬(Nitrosolobus)以及一些未培養的氨氧化細菌.從圖4可知,條帶3與亞硝化球菌屬(Nitrosococcus)處于一個分枝上,其余4個條帶均與亞硝化單胞菌屬(Nitrosomonas)的親緣關系較近,與亞硝化葉菌(Nitrosolobus)、亞硝化螺菌(Nitrosospira)的遺傳距離較遠.由于所研究的反應器進水氨氮濃度為300 mg/L,屬于較高的氨氮環境,在該條件中檢測到亞硝化球菌(Nitrosococcus)的存在,這與前人報道的亞硝化球菌屬(Nitrosococcus)在高氨氮環境中作為氨氧化細菌的結果吻合[20].此外,系統中還存在亞硝化單胞菌屬(Nitrosomonas),它是許多水生態系統中最常見的氨氧化菌類型[7].
從圖3(a)不同泳道的條帶變化情況來看,亞硝化球菌(Nitrosococcus)僅出現在濾層100及300mm處,而與亞硝化單胞菌屬(Nitrosomonas)相關的條帶1、條帶2所代表的氨氧化細菌僅在濾層下方出現.如何根據這些細菌的空間分布特點改進工藝條件以達到更好的脫氮效果,還有待于進一步的研究.
基于ANAMMOX菌16SrDNA序列構建的系統發育樹見圖5.樹圖的外源基因選取了常見的幾種ANAMMOX菌.可以看出,條帶6、7與Candidatus Kuenenia stuttgariensis遺傳距離最近,這與之前報道的在一個特定的環境條件中,只可能有一種ANAMMOX菌會成為優勢菌種的結果相一致[21].Candidatus Kuenenia stuttgariensis屬于浮霉菌屬,是污水處理系統中除Candidatus Brocadia anammoxidans之外的典型的具有厭氧氨氧化活性的微生物[22-24].存在于本系統中的ANAMMOX菌是CandidatusKueneniastuttgariensis而非Candidatus Brocadia anammoxidans,其原因在于本實驗所用污水是用自來水加一定的無機鹽配制而成,具有較高的鹽離子濃度,而Candidatus Kuenenia stuttgariensis被認為是一種存在于淡水環境中,并對較高的鹽離子濃度具有一定耐受性的ANAMMOX菌[21],因此,Candidatus Kuenenia stuttgariensis成為系統中的優勢ANAMMOX菌.

圖4 基于amoA基因序列的系統發育樹

圖5 基于ANAMMOX菌16S rDNA序列的系統發育樹
3 結論
1)上向流曝氣生物濾池型CANON反應器中,濾層下方的氨氧化細菌的種類和數量遠高于濾層上部;厭氧氨氧化菌的多樣性幾乎不隨濾層高度發生變化,其數量沿著濾層自上而下有逐漸增強的趨勢.
2)反應器中微生物形態多樣,易成簇生長,其中以直徑0.2~1.0 μm的球形及橢球形菌為主.
3)DNA測序結果表明,亞硝化球菌屬(Nitrosococcus)和亞硝化單胞菌(Nitrosomonas)是反應器中的主要氨氧化細菌,而厭氧氨氧化菌與Candidatus Kuenenia stuttgariensis的相似度高達98%.
[1]RYH H D,LEE S I.Comparison of 4-stage biological aerated filter(BAF)with MLE process in nitrogen removal from low carbon-to-nitrogen wastewater[J].Environmental Engineering Science,2009,26(1):163 -170.
[2]CHANG W S,TRAN H T,PARK D H,et al.Ammonium nitrogen removal characteristics of zeolite media in a biological aerated filter(BAF)for the treatment of textile wastewater[J].Journal of Industrial and Engineering Chemistry,2009,15(4):524-528.
[3]張文藝,夏紹鳳,翟建平,等.曝氣生物濾池反應器的沿程生化特性研究[J].中國給水排水,2006,22(15):71-74.
[4]CHUDOBA J,CECH J S,CHUDOBA P.The effect of aeration tank configuration on nitrification kinetics[J].Journal Water Pollution Control Federation,1985,57(11):1078-1083.
[5]AZIMI A A,HORAN N J.The influence of reactor mixing characteristics on the rate of nitrification in the activated-sludge process[J].Water Research,1991,25(4):419-423.
[6]王盼盼,陳建中.CANON工藝中的微生物及其相互關系[J].環境科學與管理,2007,32(8):97-100.
[7]TAO L,DONG L,JIE Z.Phylogenetic and microbial community analysis based on amoA gene and 16SrDNA in nitrosification biofilm reactor[J].Environmental Biotechnology and Materials Engineering,2011,183:1051-1056.
[8]KLOTZ M G,ALZERRECA J,NORTON J M.A gene encoding a membrane protein exists upstream of the amoA/amoB genes in ammonia oxidizing bacteria:a third member of the amo operon[J].Fems Microbiology Letters,1997,150(1):65 -73.
[9]MCTAVISH H,FUCHS J A,HOOPER A B.Sequence of the gene coding for Ammonia Monooxygenase in Nitrosomonas-Europaea[J].Journal of Bacteriology,1993,175(8):2436-2444.
[10]PURKHOLD U,POMMERENING R A,JURETSCHK O,et al.Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis:implications for molecular diversity surveys[J].Applied and Environmental Microbiology,2000,66(12):5368-5382.
[11]張亞平,阮曉紅.淺水湖泊(陽澄湖)沉積物氨氧化菌的分子證據[J].環境科學學報,2012,32(1):182-189.
[12]LING C,ZHENHUAN M.Detecting and diversity analysis of amoA gene from ammonia-oxidizing bacteria in a nitrifying pool[J].Journal of Zhejiang University(Science Edition),2004,31(5):565 -9.
[13]樊景鳳.北戴河近岸沉積物中微生物16S rDNA的PCR-RFLP分析[J].海洋環境科學,2008,27(5):409-413.
[14]葉磊,祝貴兵,倫中財,等.應用分子生物學與同位素示蹤技術研究厭氧氨氧化菌活性及功效[J].環境科學學報,2011,31(6):1206 -1211.
[15]BASSAM B J,CAETANOANOLLES G,GRESSHOFF P M.Fast and sensitive silver staining of DNA in polyacrylamide gels[J].Analytical Biochemistry,1991,196(1):80-83.
[16]SLIKERS A O,THIRD K A,ABMA W,et al.CANON and Anammox in a gas-lift reactor[J].Fems Microbiology Letters,2003,218(2):339-344.
[17]CHUANG H P,OHASHI A,IMACHI H,et al.Effective partial nitrification to nitrite by down-flow hanging sponge reactor under limited oxygen condition[J].Water Research,2007,41(2):295-302.
[18]郭建華,王淑瑩,鄭雅楠,等.實時控制實現短程硝化過程中種群結構的演變[J].哈爾濱工業大學學報,2010,42(8):1259-1263.
[19]秦玉潔,周少奇,朱明石.厭氧氨氧化反應器微生態的研究[J].環境科學,2008,29(6):1638-1643.
[20]段莎麗,孫亞琴,伍陽,等.兩株耐堿性亞硝化細菌的初步鑒定和特性研究[J].農業環境科學學報,2007(S2):406-409.
[21]HU B L,ZHENG P,TANG C J,et al.Identification and quantification of anammox bacteria in eight nitrogen removal reactors[J].Water Research,2010,44(17):5014-5020.
[22]STROUS M,FUERST J A,KRAMER E H M,et al.Missing lithotroph identified as new planctomycete[J].Nature,1999,400(6743):446-449.
[23]JETTEN M S M,WAGNER M,FUERST J,et al.Microbiology and application of the anaerobic ammonium oxidation('anammox')process[J].Current Opinion in Biotechnology,2001,12(3):283-288.
[24]YANG Q X,JIA Z J,LIU R Y,et al.Molecular diversity and anammox activity of novel planctomycete-like bacteria in the wastewater treatment system of a fullscale alcohol manufacturing plant[J].Process Biochemistry,2007,42(2):180 -18722.
