PPO處理對水曲柳合子胚子葉外植體褐化和體胚發生的影響1)
2012-07-02 00:03:54沈海龍李玉花
東北林業大學學報 2012年11期
孫 倩 楊 玲 沈海龍 李玉花 黃 劍
(林木遺傳育種國家重點實驗室(東北林業大學),哈爾濱,150040)
水曲柳(Fraxinus mandshurica Rupr.)是木犀科白蠟樹屬大喬木樹種,主要分布于我國東北,是著名的東北“三大硬闊”之一的優質用材樹種。在水曲柳體胚發生研究中發現,利用合子胚為外植體可以誘導產生水曲柳體胚[1-3],但產生體胚過程中伴隨著外植體的褐化現象,發生體胚的外植體基本上都處于褐化狀態,而且褐化外植體上誘導出的體胚長勢良好[4]。進一步研究證實,水曲柳體胚發生與外植體褐化的伴隨關系較固定,誘導產生的褐化有利于其體胚發生[5-6]。褐化是指外植體或培養材料接種后,在組織培養過程中由于切割造成機械損傷,傷口處分泌出酚類化合物,在有氧的條件下,切面細胞中的酚類物質由多酚氧化酶催化氧化為醌,醌再通過非酶促反應產生有色物質而導致組織褐變,變成棕褐色或暗褐色,并逐漸擴散到培養基中,抑制細胞內其它酶的活性,影響細胞的正常代謝,毒害整個組織,甚至導致組織死亡;尤其是木本植物組織的外植體,褐變死亡是其培養難度較大的主要因素[7-9],因而大部分組織培養過程都需要抑制外植體的褐化。李官德等[10]的研究提到,棉花的褐化且生長緩慢的愈傷組織能分化出體胚,并再生成植株。藍桉體胚發生的研究中發現,在培養基中添加抗氧化劑并不能顯著降低外植體的褐化程度,但使用降低褐化程度的抗氧化劑則可抑制體胚的發生,同時降低了多酚的積聚[11],認為多酚的積聚在體胚發生中具有一定作用,只是這種作用尚不為人知。有研究發現,水曲柳體胚發生過程中的外植體褐化比棉花[10]和藍桉[11]的表現更加明顯,因此在由合子胚子葉為外植體誘導水曲柳體胚產生的過程中伴隨著外植體的褐化,被視為一個很普遍的現象。但這種現象與絕大多數植物組織培養中外植體褐化的效應相反。因此本研究試圖揭示水曲柳體胚發生中外植體褐化與體胚發生的關系,并根據已有的研究從DNA甲基化修飾、細胞程序性死亡(PCD)和生理生化檢測等多個角度探索這個現象的機理和應用技術。
多酚氧化酶(polyphenol oxidase,PPO)在植物細胞中以游離態和束縛態兩種形式存在[12],是褐化過程中的主要酶類。因PPO的專一性,不會對酚類以外的物質作用,性質相對柔和,能夠引起植物組織的褐化,因此,設想利用在培養基中加入PPO來誘導外植體發生褐化,從而促進體胚發生。本研究通過在液體培養基中加入不同質量濃度的PPO,利用其脅迫作用促使外植體褐化,探究PPO對水曲柳體胚發生和外植體褐化及體胚發生的影響,為揭示水曲柳體胚發生過程中外植體褐化與體胚發生的關系提供依據和研究借鑒。
1 材料與方法
1.1 材料采集與處理
水曲柳(Fraxinus mandshurica Rupr.)未成熟種子和成熟種子分別于2010年7月下旬和9月下旬取自東北林業大學哈爾濱實驗林場內生長良好的40~60年生水曲柳母樹;各母樹均為自由授粉,母樹之間距離大于50 m;各母樹采集相同數目種子后充分混合再取樣用于試驗。
未成熟外植體處理:剝除外種皮后用自來水沖洗24 h,用75%乙醇浸泡1 min后,再用2%次氯酸鈉溶液消毒15 min,在超凈工作臺上用無菌水沖洗5次;剝取出單片子葉,并使子葉近軸面接觸培養基接種。
成熟外植體處理:剝除外種皮后用自來水浸泡48 h(定時換水并用攪拌器攪拌)之后再用自來水沖洗12 h;用75%乙醇浸泡1 min后,再用10%的次氯酸鈉消毒20 min,在超凈工作臺上用無菌水沖洗5次;在超凈工作臺上剝取出單片子葉,并使子葉近軸面接觸培養基接種。
1.2 體胚誘導培養
體胚誘導培養基為雙培養基(固體培養基+液體培養基,即用滅菌藥匙在固體培養基表面挖出直徑約1 cm的凹面,然后在凹面內加入0.1 mL的液體培養基)。固體和液體培養基均為MS 1/2(MS基本培養基中所有成分均減半)附加1.5 mg·L-1NAA、0.5 mg·L-1BA、70 g·L-1蔗糖、400 mg·L-1水解酪蛋白。固體培養基加7.5 g·L-1瓊脂固化。培養基的pH值在高壓滅菌前調至5.8。接種后用封口膜將培養皿封口,于25℃下暗培養。培養室的相對濕度為40%~60%。
1.3 PPO 處理
多酚氧化酶(PPO)處理:用液體培養基配制不同處理質量濃度的PPO溶液。其中:未成熟外植體分別用 1、15、25、50、100、150 mg·L-1的 PPO 浸泡;成熟外植體分別用 15、25、50、100、150 mg·L-1的PPO浸泡。處理方法:用過濾滅菌器(孔徑0.22 μm)將PPO溶液過濾后加到固體培養基上的凹面內,每凹面內0.1 mL。以不添加PPO溶液的液體培養基為對照。每個處理質量濃度重復5次,每個重復接種10片子葉外植體。每天用顯微鏡觀察并在培養的第5、10、30、60天時統計外植體生長、褐化情況和體胚發生情況。第10天褐化外植體的統計標準:肉眼能夠觀察到外植體表面1/4以上褐化,即視為外植體褐化(30 d時外植體均已基本褐化,60 d時均深度褐化)。
1.4 外植體PPO、POD和CAT的活性測定
利用前期試驗確定的褐化和體胚發生均最佳的50 mg·L-1的PPO處理進行重復培養試驗,對培養15 d時的外植體提取樣品,用于PPO、POD和CAT酶活性測定。
PPO活性的測定:取0.25 g的試驗材料及0.4 g的 PVP,冰浴研磨,并用 2.5 mL、0.1 mol·L-1、pH為6.8的檸檬酸磷酸緩沖溶液稀釋,在4℃、10000 r·min-1的條件下離心15 min,取上清液用于酶活測定。在4 mL的酶活測定體系中含有:2.9 mL、pH為6.8 的檸檬酸磷酸緩沖溶液,1 mL、0.1 mol·L-1鄰苯二酚及0.1 mL的酶液。測定398 nm處的OD值。
微博傳播的碎片化——以“占領華爾街”為例………………………………………………………………………………………燕道成(4.31)
POD活性的測定:取0.3 g試驗材料并加入5 mL、0.1 mol·L-1、pH 為 7.8 的磷酸緩沖溶液進行冰浴研磨,后置于4℃、10 000 r·min-1條件下離心20 min,取上清液用于酶活測定。在4 mL的反應體系中含有:1 mL、pH 為7.0的磷酸緩沖溶液,0.95 mL、0.2% 的愈創木粉,2 mL、0.2%H2O2及 0.05 mL酶液。在470 nm下記錄OD值。
CAT活性的測定:取0.3 g試驗材料并加入5 mL、0.1 mol/L、pH 為7.8 的磷酸緩沖溶液進行冰浴研磨,后置于4℃、10 000 r·min-1條件下離心 20 min,取上清液用于酶活測定。在3 mL的反應體系中含有:1 mL 0.2%的 H2O2,1.9 mL、pH 為 7.0 的磷酸緩沖溶液及0.1 mL酶液。在240 nm下記錄OD值。
1.5 數據處理方法
數據均為各重復處理的平均數±標準差。標準差由每個試驗的5個重復處理計算得出。采用單因子方差分析評價試驗中各影響因子的作用,并在P=0.05水平上進行差異顯著性檢驗。平均數間統計學差異用鄧肯多重比較法在P=0.01或0.05顯著性水平上檢驗。計算公式為:
外植體生長率=有生長現象的外植體個數×100/接種存活的外植體總數%;
第10天外植體褐化率=培養第10天褐化的外植體個數×100/接種存活的外植體總數%;
第10天褐化外植體體胚發生率=在培養第10天時褐化且在培養第60天時發生體胚的外植體數量×100/具有生理活性的外植體總數%;
第10天未褐化外植體體胚發生率=在培養第10天時褐化且在培養第60天時發生體胚的外植體數量×100/具有生理活性的外植體總數%;
體胚發生率=培養第60天時產生體胚的外植體個數×100/具有生理活性的外植體總數%;
第10天褐化外植體體胚比重=在培養第10天時褐化且在培養第60天時發生體胚的外植體數量×100/培養第60天時的發生體胚的外植體數量%;
第10天褐化外植體體胚比率=在培養第10天時褐化且在培養第60天時發生體胚的外植體數量×100/培養第10天時褐化的外植體數量%。
2 結果與分析
2.1 水曲柳體胚誘導培養中外植體的變化
在體胚誘導培養過程中,未成熟外植體(圖1A)從接種后3 d開始略微增大,外植體顏色從切口處和邊緣向內逐漸由乳白變成淡黃色,隨著外植體的生長,其表面略有褐色斑(圖1B),10 d以后外植體表面部分變為褐色且上面有大量的愈傷組織產生;培養20 d后,在褐化部位愈傷組織的量明顯增加,在褐變部位有少量體胚產生,多為半透明狀球型胚(圖1C);培養30 d后,體胚數量明顯增加,體胚顏色由半透明變為乳白色,此時體胚形態多為心型胚(圖1D)和魚雷型胚(圖1E),及少數體胚簇(圖1F)。40 d后可以觀察到子葉型胚(圖1G)。長體胚的外植體的褐化程度比沒長體胚的外植體的深,且長有體胚的外植體都是褐化的。這些長有體胚的褐化外植體,在用鑷子夾取時,能夠明顯地感受到該外植體具有活細胞脆性和彈性,這與一般意義上的外植體褐變致死后的松軟或干硬的失活狀態完全不同。
成熟外植體接種后經過12 d靜止期后開始生長,第15天開始略微增大,隨后外植體顏色等外觀變化與未成熟外植體相似。
2.2 PPO處理對水曲柳未成熟外植體褐化與體胚發生的影響
PPO處理的水曲柳未成熟外植體從接種后3 d開始略微增大,外植體顏色從切口處和邊緣向內逐漸由乳白變成淡黃色(圖1H),褐變比對照早,且體胚的產生都伴有外植體的褐化。添加外源PPO對水曲柳未成熟外植體的生長影響不大(未顯示數據),其中只有質量濃度為50 mg·L-1的PPO顯著抑制了培養初期外植體的生長(P<0.05)。
由于培養30 d后外植體均已基本褐化,培養60 d后外植體均已深度褐化,所以取培養10 d的外植體褐化狀況進行數據統計,結果見表1。除了100 mg·L-1質量濃度的處理褐化值偏低(等于對照的)外,培養基中添加PPO均促進水曲柳外植體的褐化,總體上比對照的褐化率高出13.4%。其中:PPO質量濃度為50 mg·L-1時外植體褐化率最大,高于其它處理及對照,培養第10天時其褐化率達到50%,比對照高出26.7%,差異達顯著性程度(P<0.05)。相應地,培養基中添加質量濃度為50 mg·L-1PPO處理的體胚發生率也最高,高于其它處理和對照,達到40.0%,比對照高出16.7%,差異顯著(P<0.05)。各PPO處理的體胚發生率雖然差異較大,有的甚至低于對照,但從培養第10天的褐化外植體中發生體胚外植體占全部發生體胚外植體的比重來看,PPO加快褐化進程的同時也明顯促進體胚發生,因為所有PPO處理在第10天褐化外植體上發生的比重均高于對照,總體上高出18.3%。其中:褐化率和體胚發生率最高的50 mg·L-1PPO處理的比重達到58.3%,高出對照29.7%;第10天褐化外植體體胚比率數據也說明了50 mg·L-1PPO處理的促進效果,其培養第10天時褐化的外植體中在第60天時發生體胚的外植體數量占到了培養第10天時褐化的外植體數量的46.7%,高出對照18.1%。除了25 mg·L-1的PPO處理外,其余處理的培養第10天時褐化外植體體胚發生率均高于對照。其中:50 mg·L-1的PPO處理高于其他處理和對照,并與對照的差異達到顯著性程度(P<0.05)。而各處理及對照的10 d未褐化外植體體胚發生率差異很小,未達差異顯著水平(P>0.05),說明PPO處理在加快外植體褐化進程的同時對其體胚發生也具有促進作用。
2.3 PPO處理對成熟外植體褐化與體胚發生的影響
PPO處理的水曲柳成熟外植體的變化與未成熟外植體一樣,但需要經過12 d左右的靜止期后狀態 才能發生改變。
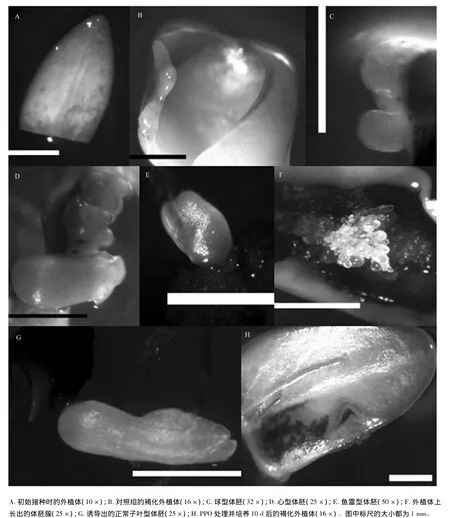
圖1 PPO處理條件下水曲柳未成熟外植體和體胚發生狀態
表2結果顯示,添加一定質量濃度的PPO會促進水曲柳成熟外植體的生長(培養第15天),但不同質量濃度的促進效果不同。其中:50 mg·L-1的PPO處理對外植體的生長抑制作用比較明顯,但差異未達顯著水平(P>0.05)。添加一定質量濃度的PPO會加快水曲柳成熟外植體的褐化進程。其中:50 mg·L-1的PPO處理培養第15天時褐化率達到40.0%,與對照差異顯著(P <0.05);50.0 mg·L-1的PPO處理培養第60天時體胚發生率雖然高于其他處理和對照,但差異未達顯著水平(P>0.05)。
2.4 水曲柳未成熟外植體PPO、POD和CAT的活性比較
選取在褐化及體胚發生中現象最為明顯的PPO質量濃度為50.0 mg·L-1處理的未成熟水曲柳外植體,與同一時間(培養第15天)培養的對照同時進行PPO、POD和CAT活性測定及比較(此時外植體的生長狀態處于體胚發生前期),結果如表3。表3結果顯示:在體胚發生前期,經PPO處理的外植體內的POD活性顯著低于對照;CAT活性低于對照,但差異不顯著;PPO活性高于對照,但差異不顯著。

表1 不同質量濃度PPO處理下水曲柳未成熟外植體褐化與體胚發生狀況 %
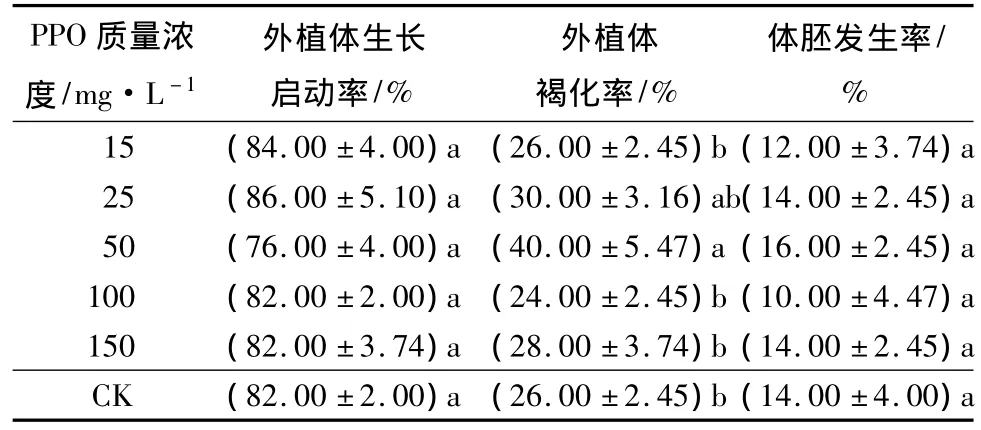
表2 PPO處理對水曲柳成熟外植體生長、褐化和體胚發生的影響
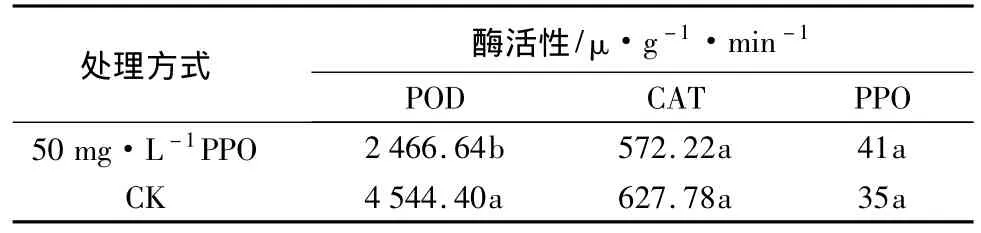
表3 體胚發生前期水曲柳未成熟外植體PPO、POD和CAT活性
3 結論與討論
3.1 水曲柳體胚發生過程中外植體的褐化現象
水曲柳體胚發生過程中伴隨著外植體的褐化現象,發生體胚的外植體基本上都處于褐化狀態,而且褐化外植體上誘導出的體胚長勢良好[3-4],體胚發生與外植體褐化的伴隨關系較固定。誘導產生褐化有利于外植體的體胚發生[5-6]。本研究進一步證明褐化進程不一致:PPO處理的外植體褐化進程快,體胚發生率高;對照褐化進程慢,體胚發生率低。但是在培養30 d后發生體胚的外植體基本上都是褐化的,而且試驗中還發現,這些發生體胚的外植體具有活細胞的脆性和彈性。試驗結果與一般意義上組織培養中的褐變表現不一樣:植物組織培養中所說的褐化,是指在外植體誘導初分化或再分化過程中自身組織從表面向培養基釋放褐色物質,以至培養基逐漸變成褐色,外植體也隨之進一步變褐而死亡的現象;褐化產物不僅使外植體、細胞、培養基褐變,而且對許多酶有抑制作用,影響培養材料的生長與分化,嚴重時甚至導致材料死亡[7-9]。深入解析水曲柳中這種特殊現象的產生機理,不僅可能為解析樹木體胚發生機理找到新的標記和明顯的切入點,也可能為解析植物組織培養中褐變現象的機理找到新的途徑。因此,這種現象值得從形態解剖學、生理學、細胞生物學和分子遺傳學等多種角度深入研究。
3.2 PPO處理可以促進水曲柳外植體褐化進程并提高體胚發生率
本研究結果說明:50 mg·L-1的PPO處理,可以顯著促進水曲柳未成熟外植體的褐化進程(培養第10天外植體褐化率顯著高于對照和其他PPO質量濃度處理),相應地顯著提高了其體胚發生率(總的體胚發生率和培養10 d時褐化外植體的體胚發生率均顯著高于對照和其他PPO質量濃度處理)。總體上,經PPO處理的外植體,其培養10 d時外植體的褐化率、總的體胚發生率、培養10 d褐化外植體體胚比重及比率、褐化外植體體胚發生率,均與對照有顯著差異,只有培養10 d的未褐化外植體體胚發生率與對照差異不顯著。對于水曲柳成熟外植體,50 mg·L-1的PPO處理也顯著促進了外植體的褐化進程,雖然體胚發生率高于其他處理和對照,但差異未達顯著水平。在培養基中添加不同質量濃度的5-氮胞苷的試驗發現[6]:抑制外植體褐化的處理質量濃度同樣抑制了體胚發生;而對外植體褐化抑制程度最低的質量濃度處理,其體胚發生率也最高;不同質量濃度對外植體褐化的作用趨勢與對體胚發生率的影響趨勢相同;在不同時間加入5-氮胞苷,顯著抑制外植體褐化的同時也明顯地抑制了體胚的發生。這與藍桉體胚發生中“使用降低褐化程度的抗氧化劑則抑制了體胚的發生”的結果一致[11],與本研究結果的趨勢相反,但都說明了同一個問題:即促進褐化則提高體胚發生率,抑制褐化則降低體胚發生率。此外,李官德等[10]的研究提到:棉花的褐化且生長緩慢的愈傷組織能分化出體胚,并再生成植株,這與本研究的結果一致:50 mg·L-1的PPO處理顯著抑制了水曲柳未成熟和成熟外植體的生長,但其外植體褐化率和體胚發生率均最高。
3.3 PPO處理促進褐化進程并進而提高體胚發生率的可能機理
本研究發現,在體胚發生前期,經PPO處理的外植體內的POD活性顯著低于對照,雖然差異不顯著,但CAT活性低于對照,PPO活性高于對照。前期研究[5]發現,褐化的外植體中的POD酶活性均顯著低于未褐化的外植體,而且只要發生愈傷組織或體胚的外植體,其POD酶活性就降低;褐化發生愈傷組織和褐化發生體胚的外植體中CAT酶活性極低。POD是植物防御中的第一道防線,POD和PPO共同氧化酚成醌,醌轉變成縮合型鞣質,最后形成褐色[13]。當植物體受到機械損傷時,受傷部位會產生系統素,經過十八烷酸途徑最后合成茉莉酸,再進一步合成寡聚半乳糖醛酸,通過外用誘導物的激發和活性氧的迸發作用,使H2O2產生并作為第二信使誘導防御基因合成相關防御蛋白,從而提高PPO活性;PPO再與酚類物質作用,出現外植體的褐化現象[14-17]。因此,水曲柳外植體褐化的過程可能是:當水曲柳的合子胚被切割后造成了一定的機械損傷,使得細胞產生大量的酚類等物質,傷口處分泌出酚類化合物,切面細胞中的酚類物質由外界的PPO催化,氧化為醌,醌再通過非酶促反應產生有色物質而導致組織褐變;由于加入PPO的外植體一直處在含有PPO和產物醌的脅迫中,因此細胞內的褐化進程加快、褐化程度持續加深,促進胚性細胞周圍細胞發生程序化死亡,從而促進胚性細胞向體細胞胚的轉化速度、提高其轉化率;而由于程序化死亡,POD和CAT的酶的溢出可能受到限制,酶活性降低。然而,對于PPO處理促進外植體褐化進程并提高體胚發生率的機制,本項研究還不能提供確切的證據,需要進一步深入研究。
[1]孔冬梅,譚燕雙,沈海龍.白蠟樹屬植物的組織培養和植株再生[J].植物生理學通訊,2003,39(6):677 -680.
[2]孔冬梅,沈海龍,馮丹丹,等.水曲柳體細胞胚與合子胚發生的細胞學研究[J].林業科學,2006,42(12):130 -133.
[3]Kong Dongmei,Preece J E,Shen Hailong.Somatic embryogenesis in immature cotyledons of Manchurian ash(Fraxinus mandshurica Rupr.)[J].Plant Cell Tiss Organ Cult,2012,108(3):485 -492.
[4]沈海龍,孔冬梅,黃劍,等.水曲柳不定芽和體胚發生中需要解決的幾個關鍵問題[G]//季孔庶.2007中國植物生理學會全國學術會議論文摘要匯編.北京:中國林業出版社,2007:256.
[5]叢建民,沈海龍,李玉花,等.水曲柳體胚發生過程中不同狀態類型外植體的生理生化狀態[J].華南農業大學學報,2012,33(1):48-52.
[6]劉艷,沈海龍,叢建民.5-氮胞苷對水曲柳合子胚外植體狀態及體胚發生的影響[J].東北林業大學學報,2011,39(8):25-27,32.
[7]葉梅,王伯初,段傳人.植物組織培養外植體褐變的研究進展[J].生物技術通訊,2004(4):426 -428.
[8]郭艷,楊海玲.植物組織培養中的褐化現象及解決途徑[J].山西農業科學,2009,37(7):14 -16,31.
[9]朱文祥,王金榮,李珺,等.植物組織培養中外植體褐變研究進展[J].安徽農業科學,2010,38(31):17399 -17401,17415.
[10]李官德,肖麗娟,羅曉麗,等.不同棉花愈傷組織狀態與胚胎發生及其植株再生的關系[J].山西農業科學,2006,34(1):29-31.
[11]Pinto G,Silva S,Park Y S,et al.Factors infiuencing somatic embryogenesis induction in Eucalyptus globulus Labill.:basal medium and anti-browning agents[J].Plant Cell Tiss Organ Cult,2008,95(1):79 -88.
[12]孔俊豪,楊秀芳,涂云飛,等.凍存茶鮮葉多酚氧化酶動力學特性研究[J].保鮮與加工,2011,11(4):9 -11.
[13]Laukkanen H,Haggman H,Kontunen-Soppela1 S,et al.Tissue browning of in vitro cultures of Scots pine:Role of peroxidase and polyphenol oxidase[J].Physiologia Plantarum,1999,106(3):337-343.
[14]段曉娜,陳宗懋,任炳忠,等.植物多酚氧化酶的誘導[J].吉林師范大學學報:自然科學版,2011(2):90-95.
[15]VanDam N M,Horn M,Mares M,et al.Ontogeny constrains systemic protease inhibitor response in Nicotiana attenuata[J].J Chem Ecol,2001,27(3):547 -568.
[16]Miyoshi H,Major I T,Mary E C,et al.A Kunitz trypsin inhibitor gene family from trembling aspen(Populus reemuloides Michx.):cloning, functional expression, and induction by wounding and herbivory[J].Plant Mol Biol,2001,46(3):347 -359.
[17]Mayer A M.Polyphenol oxidases in plants and fungi:Going places?A review[J].Phytochemistry,2006,67(21):2318 -2331.
