長白山闊葉紅松林及楊樺林下草本季節動態及環境解釋1)
2012-06-28 09:12:06夏富才
東北林業大學學報 2012年11期
關鍵詞:物種
夏富才
(北華大學,吉林,132013)
趙秀海 潘春芳 范 娟 汪金松 倪瑞強 何懷江 陳貝貝 丁勝建 姜 俊
(北京林業大學)
溫帶森林草本層生物量雖然僅占整個森林生物量的1%左右,卻包括林內90%甚至更多的物種并且能夠為森林提供20%的富含養分的枯落物。作為森林中有明顯競爭作用的一個層片,草本層可以在林冠干擾后決定森林發展的方向,并且可以和林冠層喬木形成紐帶關系[1]。在國外,一些綜述性文章[2-5]概括了有關草本層的生態學特征。土壤因子、立地類型、林分結構和光照環境被認為是影響草本層的主要因素[6-8]。在國內,長白山闊葉紅松林下的草本層片也是生態學家關注的熱點。劉琪景[9]、邵 國 凡[10]、趙 秀 海[11]、于 鳳 英[12]、夏 富才[13-14]等對早春草本層的群落特征進行了詳細的研究,郝占慶等[15]于20世紀90年代最早研究了長白山闊葉紅松林下草本植物多樣性的季節動態,發現闊葉紅松林下的草本植物1 a之內,水平和垂直結構變化較大;為了深入了解長白山闊葉紅松林下草本層的季節動態,李步杭[16]、常新華[17]、夏富才[18]等隨后對闊葉紅松林下大面積固定樣地中的草本層多樣性的季節動態做了進一步研究,研究表明坡向是影響草本層物種豐富度和多度的主要因素,1 a內草本植物的垂直結構有明顯的分層現象,次生林下的草本多樣性高于原始闊葉紅松林。然而,郝占慶[15]、李步杭[16]等只是集中研究了原始闊葉紅松林下的草本層多樣性季節動態,常新華[17]、夏富才[18]雖然將不同演替階段林分下的草本多樣性與原始闊葉紅松林下的草本多樣性進行了對比,可是在他們的研究中,次生楊樺林的面積是5.2 hm2,樣方數為460個,而闊葉紅松林的面積是1.0 hm2,樣方數為100個,兩個樣地中取樣面積和樣方數不同,由此計算出來的多樣性指標沒有可比性。郝占慶[19]在21世紀初還研究了長白山海拔700~2 600 m草本的分布與環境因子的相關性,發現海拔是影響草本分布的主導因子,林冠郁閉度、土壤有機質和速效N、P、K均對草本層有較大影響。但目前仍不清楚闊葉紅松林下的環境因子在不同季節對草本植物多樣性的影響如何,文中在以往研究的基礎上,結合草本調查和環境因子調查,旨在探討闊葉紅松林下草本層多樣性隨季節變化的特征及其可預測性。
1 研究地區概況
本研究的次生楊樺林樣地位于吉林省白河林業局光明林場(42°20.907'N,128°07.988'E),海拔 899 m;闊葉紅松林樣地位于長白山自然保護區內(42°20.211'N,128°05.705'E),海拔 784 m。研究區域立地類型為平緩的坡地,屬典型的溫帶大陸性山地氣候[20],受東面海洋季風的影響,夏季溫暖多雨,冬季寒冷干燥,氣候溫和,年平均溫度為3.6℃,年平均降水量為700 mm,主要集中在7—9月份。土壤為暗棕色森林土[21]。
次生楊樺林的蓄積量為169.16 m3·hm-2,胸徑≥1 cm的木本植物的密度為4 058株·hm-2。上層喬木樹種主要有紅松(Pinus koraiensis)、蒙古櫟(Quercus mongolica)、紫椴 (Tilia amurensis)、水曲柳(Fraxinus mandshurica)、臭冷杉(Abies nephrolepis)、白樺(Betula platyphylla)、山楊(Populus davidiana)等,中層喬木以色木槭(Acer mono)為主,林下層主要是各種萌蘗生長的槭屬類植物。闊葉紅松林的蓄積量為331.7 m3·hm-2,胸徑≥1 cm個體的密度為1 801株·hm-2。上層喬木樹種主要有紅松、蒙古櫟、紫椴、水曲柳,中層喬木以色木槭為主,林下層以簇毛槭(A.barbinerve)、毛榛(Corylus mandshurica)和暴馬丁香(Syringa reticulata)為主。
2 研究方法
2.1 樣地設置及數據收集
本研究的次生楊樺林樣地面積為5.2 hm2(200 m×260 m),闊葉紅松林樣地面積僅為1.0 hm2(100 m×100 m),于2005年7—9月份,用全站儀分別將次生楊樺林樣地和闊葉紅松林樣地劃分為20 m×20 m的亞樣方,在每個亞樣方中用木樁均勻布置4個1.0 m2(1.0 m ×1.0 m)的草本樣方。為保證次生楊樺林和闊葉紅松林具有相同的抽樣面積和樣本數,在次生楊樺林樣地中只調查樣地中最有代表性的連續的1.0 hm2(100 m×100 m)面積中的100個草本樣方。于2009年5—9月份的月初調查2個林分下的各100個草本樣方,記錄物種、株數、高度、蓋度(長軸、短軸的長度),以及開花個體數和結果個體數。
文中調查的環境因子包括土壤養分、土壤水分、葉面積指數、總光量子密度、土壤質地以及林隙狀況。具體調查方法如下:在固定樣地內按照系統布點的方法布置土壤采樣點,2006年8月份在每個20 m×20 m樣方的中線上均勻布置2個采樣點,在每個土壤采樣點的表層(0~20 cm)各取1個500 g的土壤樣品,帶回實驗室進行分析。
室內土壤樣品分析方法依據《土壤農業化學分析方法》[22]的操作步驟進行:采用銨態氮靛酚藍比色法測土壤中速效氮的含量;采用鹽酸氟化銨法測定土壤中速效磷含量;采用乙酸銨提取法測定土壤中速效鉀含量;采用高溫外熱重鉻酸鉀氧化密度法測定土壤中有機質含量;用pH計測定土壤pH值。計算2個樣品的平均值作為每個樣方的值。用手持土壤水分測定儀(HH2 DelLa-TDevices Moisture Meter,UK)測定土壤水分。
為評價林分光照情況,在2007年8月份,使用冠層分析儀(WinSCANOPY,Canada)在每個樣方中距離地面1 m處拍攝2張半球狀照片,用對應的WinSCANOPY和XLScanopy軟件處理照片,計算林下包括直射光和散射光在內的總光量子密度(PPFD),并依據 Bonhomme et al.[23]的方法測算冠層結構指標:葉面積指數(LA,I)[24]。調查每個草本樣方的林冠層狀況,如20 m×20 m中的4個1 m2的草本樣方全部在林窗下,則記為1;全部在林冠下,則記為0;其中1個在林窗下,其余3個在林冠下,記為0.75,依此類推。研究樣地地勢平坦,坡度小于5°,故文中未考慮地形對草本植物分布的影響。
2.2 數據分析方法
Kudo et al.[25]按照繁殖時間將溫帶森林下的草本植物劃分為3類:早春短命植物、早夏植物和晚夏植物。早春短命植物在積雪融化之時出土,最晚在50%冠層郁閉之前開花,冠層郁閉之后地上部分枯萎,進入休眠期,一般出現在從4月初到5月中旬;早夏植物在冠層郁閉的過程中(5月中旬到6月初)開花,在冠層郁閉之后結果;晚夏植物在冠層郁閉之后(6月下旬之后)才開始生長,在夏季開花結果。同屬溫帶地區的長白山闊葉紅松林林下草本植物也完全符合這一規律,即該林下草本層片也存在明顯的季節特征。按照這一劃分標準,將闊葉紅松林及其次生楊樺林下的草本劃分為以上3種不同的物候類型。
用Jaccard指數(J)分析同一林分不同月份間以及不同林分同一月份間的物種相似度。
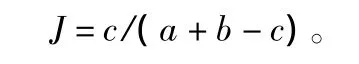
式中:a為群落A中的物種數;b為群落B中的物種數;c為群落A和群落B之間的共有物種數[26]。
用物種豐富度(S,每個樣方中的物種數)、物種多度(A,每個樣方中的總個體數)、香濃多樣性指數[27](H,H=- ∑pilnpi,pi表示第 i個物種的個體數占所有物種個體數的比例)和均勻度指數(E,E=eH/S)[28]表示草本植物的多樣性格局。用999次隨機置換的雙因素方差分析檢驗次生楊樺林和闊葉紅松林樣地之間,以及調查季節(5—9月份)的多樣性差異。方差分析前將數據經過對數轉換,以使數據服從正態分布和滿足方差齊性。
稀疏曲線是通過對N個個體或樣本進行多次可放回完全隨機抽樣,然后以個體數或樣本數序列(1,2,…,N)為橫坐標,以物種豐富度為縱坐標繪制的曲線。所調查的樣本一般不通過重復抽樣產生[29]。參考稀疏曲線的概念,假設樣地內所調查的100個草本樣方隨機分布,逐次抽取1,2,…N,…100個1 m2的草本樣方,用可放回抽樣的方法重復抽取999次。將每次抽到的N個草本樣方合并為1個大樣本統計物種豐富度,將999次重復抽樣得到的999個大樣本的物種豐富度計算平均值,作為N個樣本時候的物種豐富度繪制稀疏曲線。用冪函數、對數函數和邏輯斯蒂函數分別擬合稀疏曲線。用殘差平方和檢驗擬合效果。檢驗是否顯著偏離真值的最簡單的辦法就是檢驗真值是否落在模型參數的95%置信區間內[29]。如果相關系數的置信區間沒有重疊,就認為存在顯著差異。文中所有的顯著性和置信區間都在α=0.05水平下計算。以上過程通過R2.10.0軟件編程實現。
999次隨機置換的雙因素方差分析用anova.2way.R代碼實現。
為了分析環境因子與草本植物多樣性的關系,環境因子與各多樣性指標的相關性用corPerm2程序在R2.10.0軟件下執行999次隨機置換檢驗,將相關性顯著的環境因子作為解釋變量,將草本植物的多樣性格局作為響應變量,用 rdaTest軟件包在R2.10.0軟件下做999次隨機置換的典型相關(RDA,redundancy analysis)檢驗。
3 結果與分析
3.1 物種的基本特征
在次生楊樺林中調查到84個物種,隸屬于35科,64屬;在闊葉紅松林中共調查到76個物種,隸屬于31科,57屬。其中,闊葉紅松林蕨類植物5種,次生楊樺林蕨類植物9種。2個林型草本層共有被子植物52種,只在次生楊樺林中草本層出現的被子植物19種,只在闊葉紅松林草本層出現的被子植物24種。次生楊樺林中的草本植物包括8種早春短命植物、36種早夏植物和31種晚夏植物;闊葉紅松林中的草本植物包括13種早春短命植物,37種早夏植物和21種晚夏植物。
3.2 物種多樣性格局
Jaccard相似度指數表明,在次生楊樺林和闊葉紅松林中,5月份草本植物的物種與其他4個月份的相似度最低;次生楊樺林中,6月份和7月份的相似度最高,其次是7月份和8月份的;闊葉紅松林中,7月份和8月份的相似度最高,其次是6月份和7月份的(表1)。相比于同一樣地各月份之間的相似度指數,次生楊樺林和闊葉紅松林之間的草本物種相似度在各月份均較低,從低到高依次為:0.337(8月份);0.370(9 月份);0.385(5 月份);0.422(7月份);0.429(6月份)。
999次隨機置換的雙因素方差分析表明,不同季節之間的物種多度和物種豐富度在0.001水平上存在顯著差異,香濃指數在0.05水平上存在顯著差異;2塊樣地之間的物種多度和香濃指數在0.001水平上存在顯著差異,物種豐富度在0.05水平上存在顯著差異(表2),這表明不僅同一樣地各月份之間的物種多樣性變化較大,同一月份2塊樣地之間也存在極顯著差異。而季節之間和樣地之間的均勻度指數的差異均不顯著。
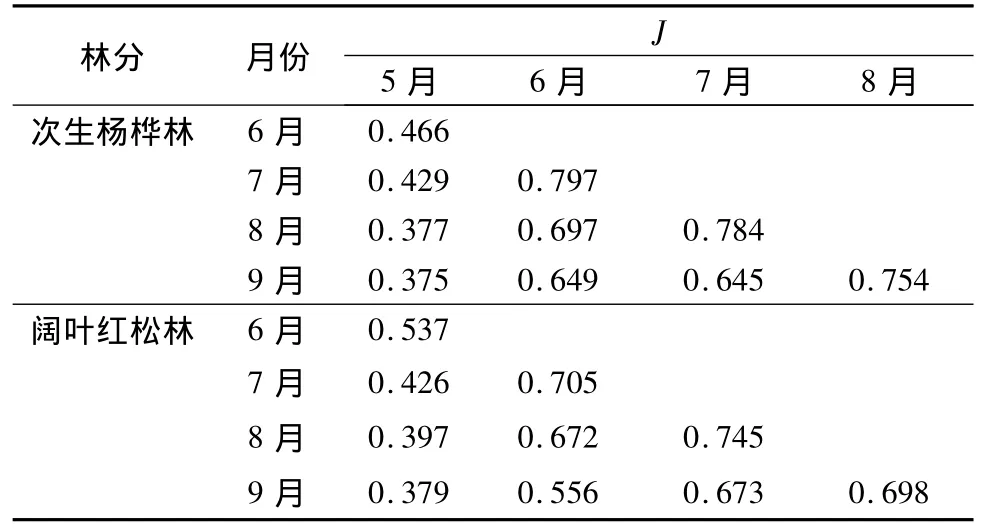
表1 次生楊樺林各月份之間的Jaccard相似指數
5月份闊葉紅松林中的物種豐富度最高,其他月份次生楊樺林中的物種豐富度最高。從5—9月份,闊葉紅松林中的香濃指數和均勻度指數始終高于次生楊樺林,香濃指數增幅為11.02% ~41.65%,均勻度指數增幅為12.25% ~55.48%,這說明闊葉紅松林下草本層生物多樣性水平相對較高 (表3)。進一步研究發現,只在次生楊樺林出現的物種有17種,主要是北野豌豆(Vicia amoena)、木賊(Equisetum hiemale)、單穗升麻(Cimicifuga simplex)、草地風毛菊(Saussurea amara)、魁蒿(Artemisia princeps)等喜陽植物,只在闊葉紅松林出現的物種有23種,主要是早春短命植物側金盞(Adonis amurensis)、反萼銀蓮花(Anemone reflexa)、單花韭(Allium monanthum)、朝鮮頂冰花(Gagea lutea)、鮮黃連(Jeffersonia dubia)、紫花變豆菜(Sanicula rubriflora)、荷青花(Hylomecon japonica)、東北擬扁果草(Isopyrum manshuricum)、延胡索(Corydalis spp.)和輪葉百合(Lilium distichum)、興安鹿藥(Smilacina davurica)、類葉牡丹(Caulophyllum robustum)等耐陰植物。
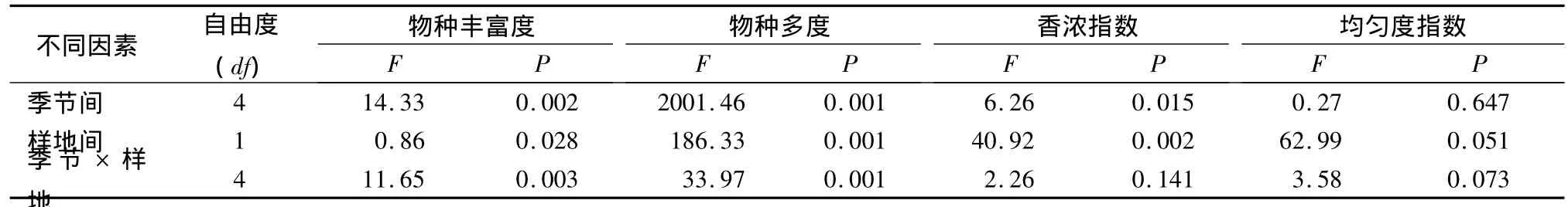
表2 對物種豐富度、物種多度、香濃指數和均勻度指數進行999次隨機置換的雙因素方差分析
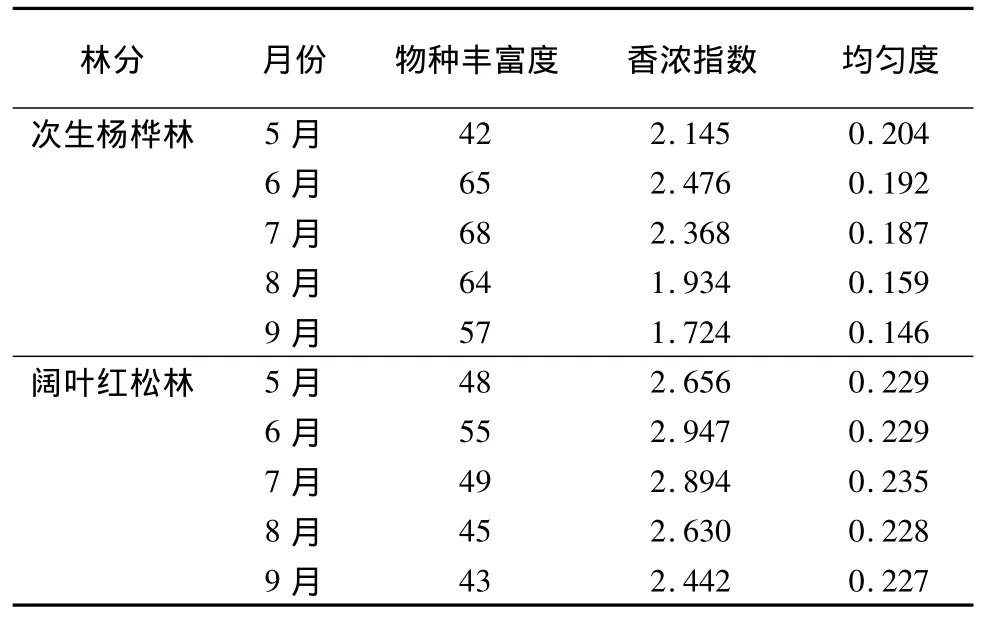
表3 物種豐富度、香濃指數和均勻度指數
稀疏曲線描述草本樣方的數量與物種數的關系,隨著樣本數量的增多,曲線的斜率逐漸減小,但仍然是遞增趨勢。當樣本數量增加到100個時,次生楊樺林中5—9月份的物種數分別為42、65、68、64、57 種,闊葉紅松林中分別為 48、55、49、45、43種。假設所有草本樣方隨機分布在調查樣地內,999次隨機抽樣得到的稀疏曲線(圖1)和表3的結果一致:在5月份,闊葉紅松林中的物種豐富度高于次生楊樺林,從6月份開始,次生楊樺林中的物種豐富度更高。相比于對數模型和冪指數模型,邏輯斯蒂模型能夠較好地擬合實際的稀疏曲線,對次生楊樺林各月份稀疏曲線的擬合效果比闊葉紅松林的好(表4)。由表1和表3可以看出,從7月份到8月份,草本植物的各項指標變化較小,該時期屬于夏季草本植物的生長期,故在本研究中用7月份的調查結果代表夏季中期草本植物的多樣性。
以上分析可知,原始林與次生楊樺林各個月份之間物種數變化趨勢均經歷了由低到高,再由高到低的變化。早春階段同一林型不同月份之間的物種相似度在5月份最低,夏季最高;2種林分各個月份之間的相似度均較低,這是因為在次生楊樺林內喜陽植物相對較多,而原始林內耐陰植物相對較多。除了5月份,次生楊樺林的物種豐富度均高于原始林。在闊葉紅松林區,不同月份之間以及不同林型之間,草本層物種多樣性變化較大。
3.3 物種多樣性與環境因子的相關性
因5月份林分沒有郁閉,所以不分析5月份葉面積指數和總光量子密度對草本植物多樣性的影響;999次隨機置換的相關性檢驗表明,次生楊樺林中6、7、9月份的物種多樣性與總光量子密度顯著正相關,6月和7月的草本植物蓋度還與葉面積指數顯著正相關,7月和9月的蓋度和總光量子密度正相關。9月份總光量子密度對物種多樣性變異的可解釋量高達33.7%。土壤pH值和速效磷對9月份的草本蓋度和多樣性有一定影響(表5)。
闊葉紅松林中各月份的草本蓋度與林隙大小顯著正相關,而物種多樣性與林隙大小呈顯著負相關,這說明在光照充足的林隙下,草本植物蓋度較大,優勢種的強勢生長導致林隙草本植物的種類較少。春、夏季節(5、6、7月份)的物種多樣性與土壤pH值呈極顯著負相關。除此之外,春季和初夏(5、6月份)的草本蓋度、多樣性和物種數還受土壤有機質含量、土壤水分的影響,夏秋季節(7、9月份)的草本蓋度及物種數受光照環境的影響。土壤水分和林隙對5月份草本蓋度格局的可解釋量高達57.72%。環境因子對春夏季節草本植物的影響極大。6月份林冠郁閉,光照環境發生劇烈變化,環境對草本植物的影響最大,分別可以解釋物種多樣性格局的59.34%和物種豐富度格局的55.96%。
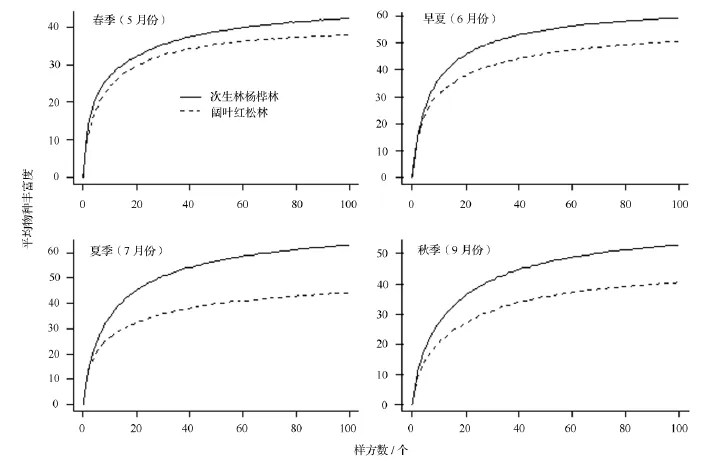
圖1 由1.0 hm2樣地中的100個1 m2草本樣方得到的稀疏曲線
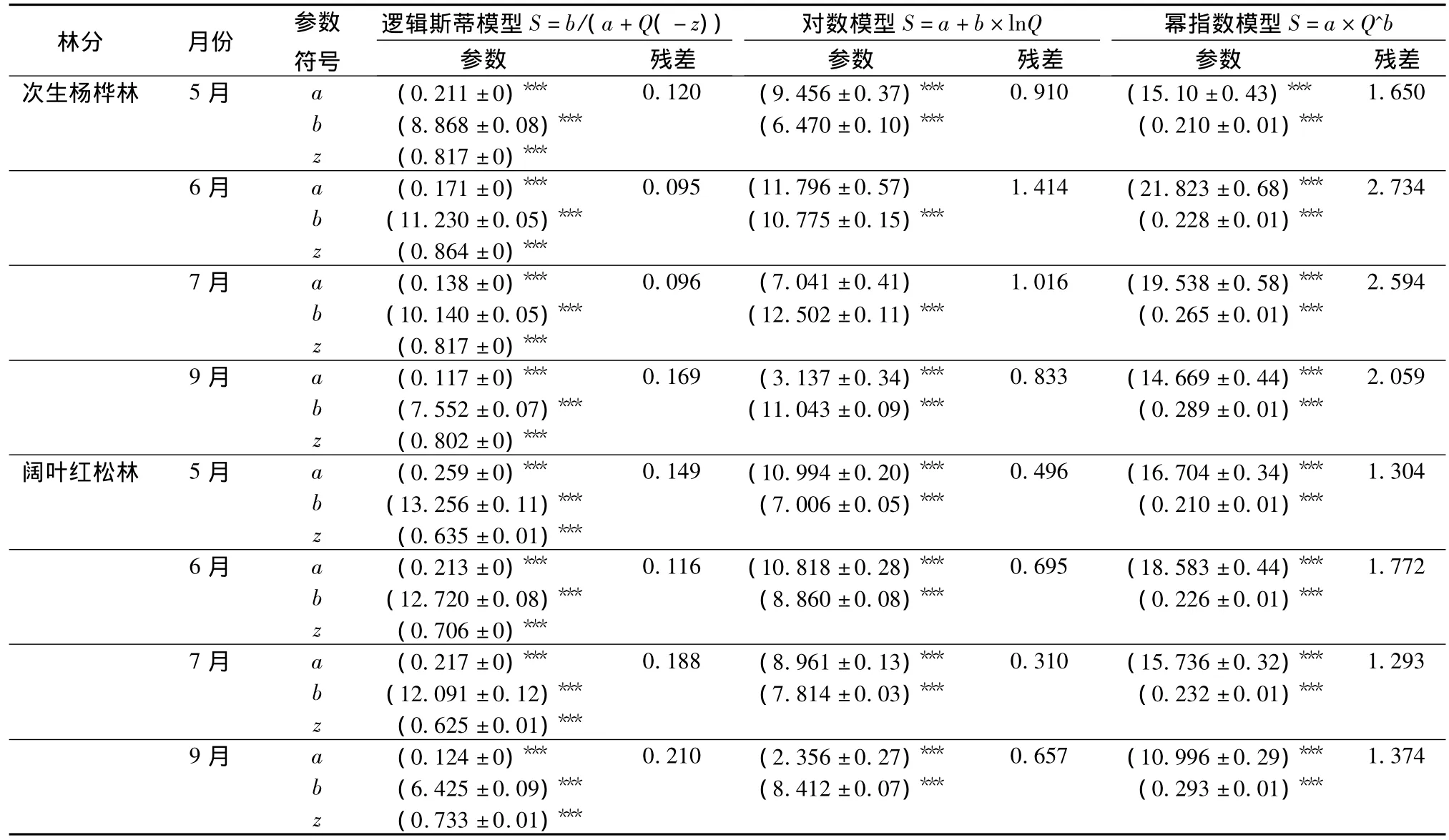
表4 3種物種累計曲線模型的比較
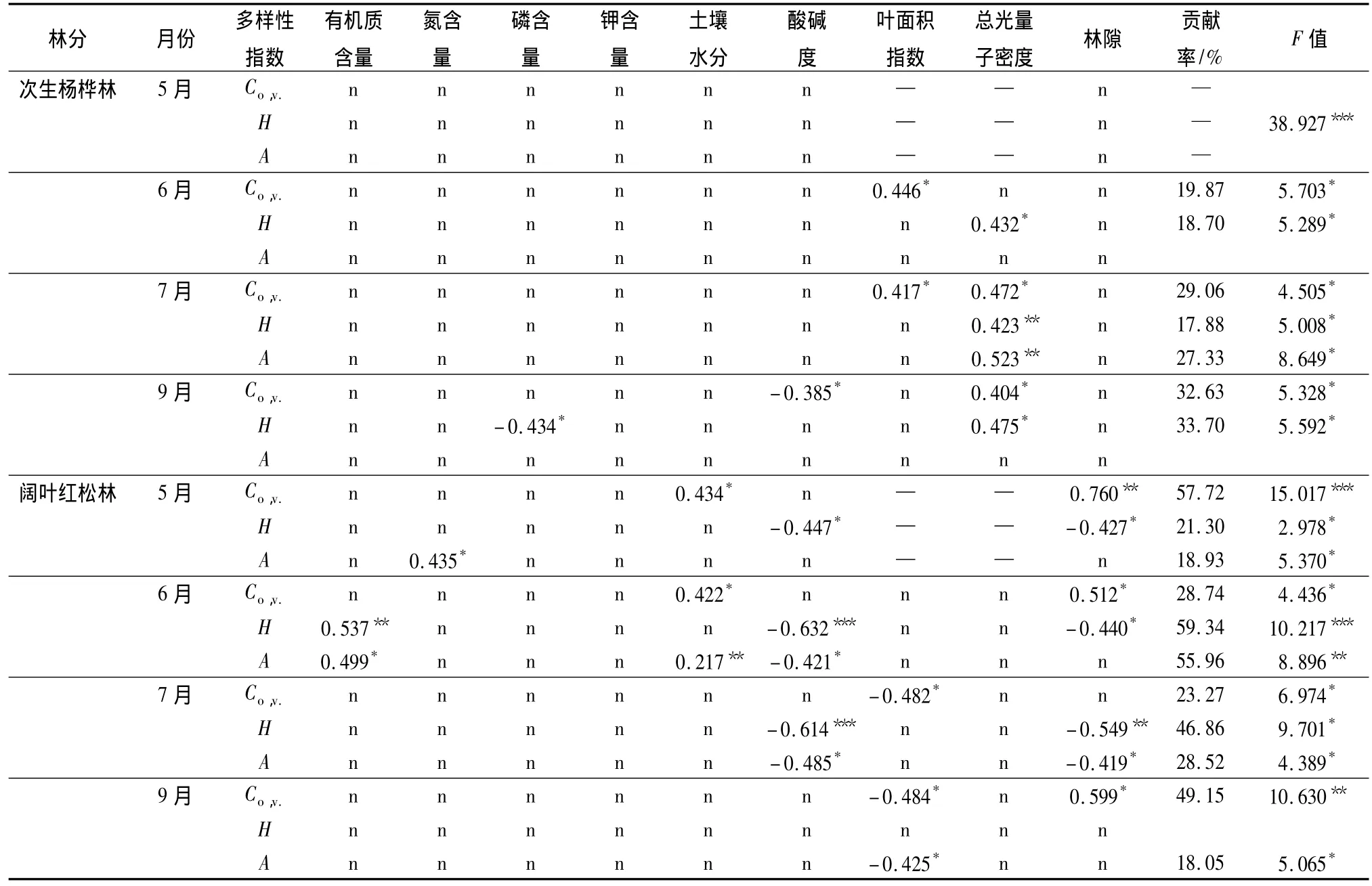
表5 次生林及闊葉紅松林下草本蓋度、香濃指數和物種豐富度與環境因子的關系
4 結論與討論
闊葉紅松林下的草本植物存在明顯的季節性差異,早春和夏、秋季之間的差異最大。這主要是因為5月份早春短命植物極占優勢,作為溫帶森林下一個特殊的森林層片,早春短命植物在喬灌木樹種處于休眠狀態時就開始生長,林分郁閉前完成開花、結果等各個生活史階段,林分郁閉后結束生長,進入休眠期[29]。
6月初冠層郁閉時很多早春短命植物的地上部分已經枯萎,林下以在林冠郁閉時開花、林冠郁閉之后結果的早夏植物和在林冠郁閉之后開花、結果的晚夏植物為主。早夏植物和晚夏植物生長到夏末秋初才逐漸枯萎。因此,早春和夏、秋季節的草本植物差異最大。
5月份闊葉紅松林中的物種數高于次生楊樺林,夏、秋季節闊葉紅松林的物種數少于次生楊樺林。闊葉紅松林中的多樣性指數始終較高。這主要是因為5月份氣溫較低,闊葉紅松林中早春短命植物較占優勢,而在受過采伐干擾的次生楊樺林中,這些短命植物較少,喜陽植物尚未全部萌發,故物種數較少。香濃多樣性指數是將物種數與物種多度相結合計算的指標,6—9月份次生楊樺林中的物種數更多,而多樣性指數更低,說明在次生楊樺林內物種多度的差異較大。這可能是因為干擾之后次生楊樺林中環境的異質性,尤其是光照環境的異質性增大,有些環境敏感物種只在某些微環境下幸存。Aikens et al.[30]認為雖然剛開始一些植物在強度干擾和中度干擾區域不復存在,但是他們能夠在相對短的時間里重新恢復優勢。然而,那些對干擾很敏感的當地稀有物種將可能永遠消失。因此,林業經營中應該盡量減少對地表層的干擾,并通過保留木產生半遮陰的更新環境,盡可能減少物種的喪失。
相比于冪函數模型和對數函數模型,邏輯斯蒂模型能較好地擬合闊葉紅松林及其次生楊樺林下的稀疏曲線,即樣方數與物種數的關系。但由于稀疏曲線理論的前提假設是所有樣方完全隨機分布,而由于環境異致性和種子散播等原因,草本植物在林內的分布并不是完全隨機的,邏輯斯蒂模型只能在一定程度上預測闊葉紅松林中的物種數。
光照是影響次生楊樺林草本多樣性的主要因素,林隙分布和土壤酸堿度是影響闊葉紅松林草本多樣性的主要因素。RDA分析表明,環境因子對闊葉紅松林的草本蓋度、多樣性及物種數變異的可解釋量較高,因此,可以通過這些指標在一定程度上預測闊葉紅松林中草本多樣性的高低,而環境因子對次生楊樺林中草本的影響較弱。這可能是因為采伐干擾后的次生楊樺林內環境條件較復雜,可能還有其他未知因素影響著次生楊樺林下的草本植物多樣性格局。與總光量子密度的正相關關系說明次生楊樺林下的草本植物受光照制約,冠層光照的改變對草本層具有非常大的影響。可是,草本植物與林隙的相關性不顯著,這可能是因為次生楊樺林下小灌木較多,較難辨別林隙。而在闊葉紅松林,草本蓋度與林隙呈顯著正相關,與葉面積指數呈顯著負相關,多樣性與林隙呈負相關。郝占慶等[19]對長白山北坡草本植物與環境因子相關性的研究表明,除了海拔,林冠郁閉度對草本植物的影響也比較明顯。Scheiner[31]認為林隙大小和草本種群大小呈正相關,Valverde et al.[32]認為冠層光照改變后,Primula vulgaris形成非平衡復合種群,而原有的一些種群可能會因為冠層變化導致改變,甚至滅絕。因此,闊葉紅松林中多樣性和蓋度對林隙的不同反應可能是因為在林隙下優勢物種在數量上占有極大的競爭優勢。但也有研究認為,林隙下物種多樣性更高[33]。同時,在闊葉紅松林,總光量子密度與林隙的相關性不顯著,這與 M?lder et al.[34]對海尼西公園山毛櫸林下草本層多樣性的研究結果相同,說明在闊葉紅松林中,光照并不是影響草本層多樣性的主要因子。而真正影響草本層多樣性變化的因素可能是隨著林隙出現而改變的其他環境因子,如局部溫濕度、紫外線強度、pH值和土壤養分等。
[1]Gilliam F S.The ecological significance of the herbaceous layer in temperate forest ecosystems[J].Bioscience,2007,57(10):845-858.
[2]Bierzychudek P.Life histories and demography of shade-tolerant temperate forest herbs:a review[J].New Phytologist,1982,90(4):757-76.
[3]Whigham D E.Ecology of woodland herbs in temperate deciduous forests[J].Annual Review of Ecology Evolution and Systematics,2004,35:583-621.
[4]Roberts M R.Response of the herbaceous layer to natural disturbance in North American forests[J].Canadian Journal of Botanyrevue Canadienne de Botanique,2004,82(9):1273-1282.
[5]Onaindia M,Dominguez I,Albizu I,et al.Vegetation diversity and vertical structure as indicators of forest disturbance[J].Forest Ecology and Management,2004,195(3):341-354.
[6]Svenning J C,Skov F.Mesoscale distribution of understorey plants in temperate forest(Kalo,Denmark):the importance of environment and dispersal[J].Plant Ecology,2002,160(2):169-185.
[7]H?rdtle W,von Oheimb G,Westphal C.The effects of light and soil conditions on the species richness of the ground vegetation of deciduous forests in northern Germany(Schleswig-Holstein)[J].Forest Ecology and Management,2003,182(1-3):327-338.
[8]Kolb A,Diekmann M.Effects of environment,habitat configuration and forest continuity on the distribution of forest plant species[J].Journal of Vegetation Science,2004,15(2):199-208.
[9]劉琪景,趙士洞,代洪才.闊葉紅松林與楊樺林早春階段草本植被幾個數量特征[J].森林生態系統研究,1994(7):28-36.
[10]邵國凡,劉琪景,錢宏.長白山闊葉紅松林早春草本層的群落生態學初步研究[J].森林生態系統研究,1992(6):24-31.
[11]趙秀海,戚繼忠,孫廣仁,等.長白山闊葉紅松林早春草本層植物的數量特征[J].吉林林學院學報,1997,13(1):5-9.
[12]于鳳英,周永斌,安云全,等.長白山闊葉紅松林早春植物群落特征研究[J].沈陽農業大學學報,2003,34(6):430-433.
[13]夏富才,趙秀海,張春雨,等.長白山紅松闊葉林及其次生林早春植物群落特征研究[J].吉林農業大學學報,2008,30(2)166-171.
[14]夏富才,張春雨,趙秀海,等.早春草本植物群落結構及其聚類分析[J].東北師大學報:自然科學版,2008,40(4):109-114.
[15]郝占慶,趙士洞,陶大立.長白山北坡闊葉紅松林草本植物物種多樣性及其季節動態[J].生物多樣性,1994,2(3):125-132.
[16]李步杭,張健,姚曉琳,等.長白山闊葉紅松林草本植物多樣性季節動態及空間分布格局[J].應用生態學報,2008,19(3):467-473.
[17]常新華,夏富才,潘春芳,等.長白山紅松闊葉林及其次生林草本植物多樣性季節動態研究[J].林業資源管理,2008(4):89-92.
[18]夏富才,趙秀海,潘春芳,等.長白山紅松闊葉林草本植物物種多樣性季節動態[J].吉林農業大學學報,2009,31(1):27-31.
[19]郝占慶,郭水良.長白山北坡草本植物分布與環境關系的典范對應分析[J].生態學報,2003,23(10):2000-2008.
[20]李文華,鄧坤梅,李飛.長白山主要生態系統生物量初步研究[J].森林生態系統研究,1981(2):34-50.
[21]許廣山,丁桂芳,張玉華,等.長白山北坡主要森林土壤有機質及其特征的初步研究[J].森林生態系統研究,1980(1):215-220.
[22]中國土壤學會.土壤農業化學分析方法[M].北京:中國農業科學技術出版社,1999.
[23]Bonhomme R,Chartler P.The interpretation and automatic measurement of hemispherical photographs to obtain sunlit folige area and gap frequency[J].Israel Journal of Agricultural Research,1972,22:53-61.
[24]張春雨,趙秀海,趙亞洲.長白山溫帶森林不同演替階段群落特征[J].植物生態學報,2009,33(6):1090-1100.
[25]Kudo G,Ida T Y,Tani T.Linkages between phenology,pollination,photosynthesis,and reproduction in deciduous forest understory plants[J].Ecology,2008,89(2):321-331.
[26]張金屯.數量生態學[M].北京:科學出版社,2004.
[27]Buzas M A,Gibson T G.Species diversity:benthonic foraminifera in western North Atlantic[J].Science,1969,163:72-75.
[28]Gotelli N J,Colwell R K.Quantifying biodiversity:procedures and pitfalls in the measurement and comparison of species richness[J].Ecology Letters,2001,4:379-391.
[29]Wang X G,Hao Z Q,Ye J,et al.Spatial pattern of diversity in an old growth temperate forest in Northeastern China[J].Actaoecologica,2008,33:345-354.
[30]Aikens M L,Ellum D,McKenna J J,et al.The effects of disturbance intensity on temporal and spatial patterns of herb colonization in a southern New England mixed-oak forest[J].Forest E-cology and Management,2007,252(1-3):144-158.
[31]Scheiner S M.Population dynamics of an herbaceous perennial Danthonia spicata during secondary forest succession[J].The A-merican Midland Naturalist,1988,119(2):268-281.
[32]Valverde T,Silvertown J.Variation in the demography of a woodland understorey herb(Primula vulgaris)along the forest regeneration cycle:projection matrix analysis[J].Journal of Ecology,1998,86(4):545-562.
[33]Bianba D,Guo Q,Ci B,et al.Effects of gap in primitive subalpine fir forest on diversity of herb and shrub in Tibet[J].Chinese Journal of Applied Ecology,2004,15(2):191-194.
[34]M?lder A,Bernhardt-R?mermann M,Schmidt W.Herb-layer diversity in deciduous forests:Raised by tree richness or beaten by beech?[J].Forest Ecology and Management,2008,256(3):272-281.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
學生天地(2019年36期)2019-08-25 08:59:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
