過量表達魯氏酵母耐鹽基因GPD1對釀酒酵母的影響
2012-05-08 11:51:14侯麗華于雁飛王春玲
天津科技大學學報 2012年6期
侯麗華,于雁飛,王 川,王春玲
(食品營養與安全教育部重點實驗室,天津科技大學食品工程與生物技術學院,天津 300457)
魯氏酵母(Zygosaccharomyces rouxii)又稱 S酵母,主要應用于醬油的高鹽稀態發酵過程中,用以改善醬油的風味[1-2].S酵母是與模式生物釀酒酵母相近的半子囊菌酵母.魯氏酵母可在高鹽的環境中生長,比如,在醬油高鹽稀態發酵工藝的后期鹽水發酵中含鹽量高達17%,魯氏酵母依然能夠生長及發酵[3].但是在此環境中包括釀酒酵母在內的大多數酵母都不能生長.
目前,一些關于魯氏酵母的耐鹽、耐滲的基因,如 ZrPMA1、ZrSOD2、ZrSOD22、ZrGPD1、ZrHOG1和 ZrHOG2等已經被克隆和測序[4-6].其中,GPD1即 3–磷酸甘油脫氫酶(glycerol-3-phosphate dehydrogenase)的編碼基因,其編碼的酶可使磷酸二羥丙酮生成 3–磷酸甘油,從而增加甘油含量,在酵母耐鹽的滲透調節中發揮舉足輕重的作用[7].因此,本文對魯氏酵母的耐鹽基因 GPD1進行研究,使其在模式酵母即釀酒酵母實驗室菌株W303中過量表達,研究其對釀酒酵母耐鹽性、甘油產量等的影響,為構建耐鹽菌株奠定理論基礎,為闡述酵母菌株的耐鹽機制提供更多的理論依據.
1 材料與方法
1.1 材料
1.1.1 菌株、質粒和培養基
魯氏酵母(Zygosaccharomyces rouxii)、釀酒酵母W303(Saccharomyces cerevisiae W303,MATa,leu2-3/112,ura3-1,trp1-1,his3-11/15,ade2-1,can1-100,GAL,SUC2,mal 0)、多拷貝酵母(S. cerevisiae)與大腸桿菌(E. coli)的穿梭質粒 YEplac195(Ampr,URA3),為本實驗室保存.
YPD 培養基(g/L):酵母浸粉 10,葡萄糖 20,蛋白胨 20,121,℃滅菌 20,min.其中,40%葡萄糖溶液115,℃滅菌20,min后,加入培養基中使含量達到2%.
CM-ura培養基(g/L):無氨基酸的酵母氮源(YNB)6.7,腺嘌呤0.05,組氨酸0.1,色氨酸0.1,亮氨酸0.1,精氨酸0.02,天冬氨酸0.1,谷氨酸0.1,異亮氨酸0.03,賴氨酸0.03,甲硫氨酸0.02,苯丙氨酸0.05,絲氨酸0.15,蘇氨酸0.15,酪氨酸0.03,纈氨酸0.15,調節pH為6.5,加入1.5%的瓊脂,121,℃滅菌 15,min.
1.1.2 試劑與儀器
十六烷基三甲基溴化銨(CTAB),北京鼎國生物技術有限公司;瓊脂糖,Gene Tech(上海)有限公司;Easy Pfu DNA Polymerase、Easy Pfu 10×buffer、Ligase 10×Buffer、T4,DNA Ligase,加拿大 Fermentas生物技術公司;2.5,mmol/L dNTPs,北京全式金生物技術公司;HindⅢ內切酶、10×M Buffer、SphI 內切酶、10×H Buffer,日本東洋坊公司;腺嘌呤、組氨酸、色氨酸、亮氨酸等 15種氨基酸、無氨基酸的酵母氮源(YNB),BBI公司.
電熱恒溫水浴鍋,北京市永光明醫療儀器廠;飛鴿牌微量離心機,上海安亭科學儀器廠;My CyclerTM PCR 儀、DYY–6C 型水平電泳儀、凝膠成像儀,美國 Bio-Rad公司;紫外分光光度計,上海精密科學儀器有限公司.
1.2 方法
1.2.1 引物設計
根據魯氏酵母基因數據(http://www.genolevures.org/zyro.html,序號為 ZYRO0A055390g)設計引物,GDP1-up:5′-GCGCATGCGTTGTTGTTGTC ATCACTC-3'(SphI) ;GDP1-dn :5′-GCAAGCTTTT GTCTTTCTAATACAC-3′(HindⅢ).GPD1 目的片段上游引物 5′端引入 SphI酶切位點(GCATGC),下游引物 5′端引入 HindⅢ酶切位點(AAGCTT).引物由上海生工公司合成.
1.2.2 構建質粒
提取魯氏酵母基因組,并以其為模板,以 GDP1-up和 GDP1-dn為引物作 PCR擴增(95,℃模板預變性 3,min;95,℃模板變性,55、58、60,℃引物退火1.5,min,72,℃引物延伸 2,min,共 30個循環;最后72,℃再延伸 10,min).純化 PCR產物,并用 SphI和HindⅢ雙酶切,將酶切產物連入到同樣雙酶切的YEplac195中,得到重組質粒YEplac195-GPD1.
1.2.3 耐鹽性變化實驗
分別取 200,mL滅菌后含鹽量為 0、9%、18%的YPD 培養基各 3瓶,其中分別接種 106,mL–1的菌種2,mL.將上述樣品置于搖床中 180,r/min、30,℃培養,每隔4,h取樣測A600的值.
1.2.4 抗性實驗
將待測菌株接種到 YPD液體培養基中培養(30,℃,12,h,轉速<200,r/min);測 A600值,計算細胞濃度,取出約含5×106個細胞的菌液加入到1,mL新鮮培養液中活化 2,h;測 A600值,計算細胞濃度,取出約含2×106個細胞的菌液,10,000,r/min離心1,min,棄上清液;加入20,μL無菌水混勻,終濃度為1×106,μL-1.取 2,μL 加入到 18,μL 無菌水中稀釋 10倍.同樣方法按濃度遞增順序分別得到1×106、1×105、1×104、1×103、1×102,μL-1的菌液.每個稀釋度取 3,μL菌液滴在所需平板上,超凈臺晾干后,30,℃倒置培養.
1.2.5 甘油產量的測定
Cu(OH)2懸濁液的配制:取 5支試管.各注入1,mL 0.05,g/mL 的 CuSO4溶液和 3.5,mL 0.05,g/mL的NaOH溶液,振蕩,生成Cu(OH)2懸濁液.
甘油銅溶液的配制:向上述 Cu(OH)2懸濁液的試管中分別注入 6‰、7‰、8‰、9‰、10‰的甘油標準品溶液 0.5,mL,充分振蕩,即生成甘油銅溶液,離心分離,取上清液備用;以超純水為空白,630,nm 為測定波長.以 Cu(OH)2為橫坐標,630,nm 下的吸光度為縱坐標繪制標準曲線.樣品如上操作,將測得的值代入公式計算即可得出甘油產量[8].
2 結果與討論
2.1 工程菌株的構建
將構建的質粒 YEplac195-GPD1(見圖 1)轉入大腸桿菌感受態細胞(TOP10)中,提取質粒后,用 SphI與 HindⅢ雙酶切驗證,瓊脂糖凝膠電泳檢測(見圖2).然后將質粒 YEplac195-GPD1和空質粒 YEplac 195分別轉入釀酒酵母實驗室菌株W303中[9],得到工程菌株WYS和對照菌株WY.
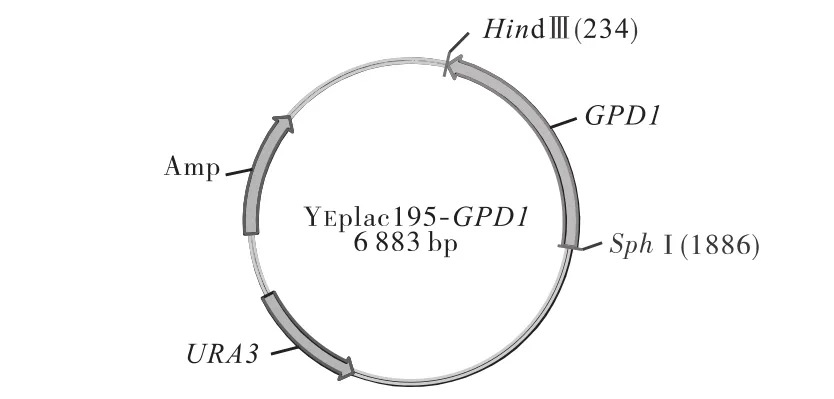
圖1 質粒YEplac195-GPD1結構示意圖Fig.1 Schematic view of the plasmid YEplac195-GPD1

圖2 重組表達質粒的SphI 和HindⅢ雙酶切鑒定Fig.2 Analysis of recombined plasmid digested by SphI and HindⅢ
2.2 耐鹽性變化
對工程菌株 WYS和對照菌株 WY在含鹽量為0、9%、18%的 YPD 培養基中的生長情況進行研究,實驗結果如圖3所示.
因為在一定濃度下,酵母細胞在 600,nm下的吸光度與細胞個數有對應關系,所以本文中通過測定不同菌株在600,nm波長下的吸光度來反映菌株的生長狀況.由圖 3可知,隨著鹽的加入,菌株 WYS和對照菌株 WY到達對數期的時間變長.這說明鹽的加入,使得細胞內外滲透壓發生變化,從而影響細胞增殖,導致到達對數期的時間延長且延緩對數期細胞的增殖.但是,在每個含鹽量下,WYS與對照菌株 WY相比,對細胞的抗滲透壓耐鹽作用仍舊可以明顯表現出來.
由上述分析可知,重組后的質粒 YEplac195-GPD1明顯比空質粒 YEplac195促進了酵母細胞的生長,不僅表現在對數生長期的提前,而且平穩期的細胞數量也有所增加,這說明基因 GPD1對于酵母細胞耐鹽性的重要作用.
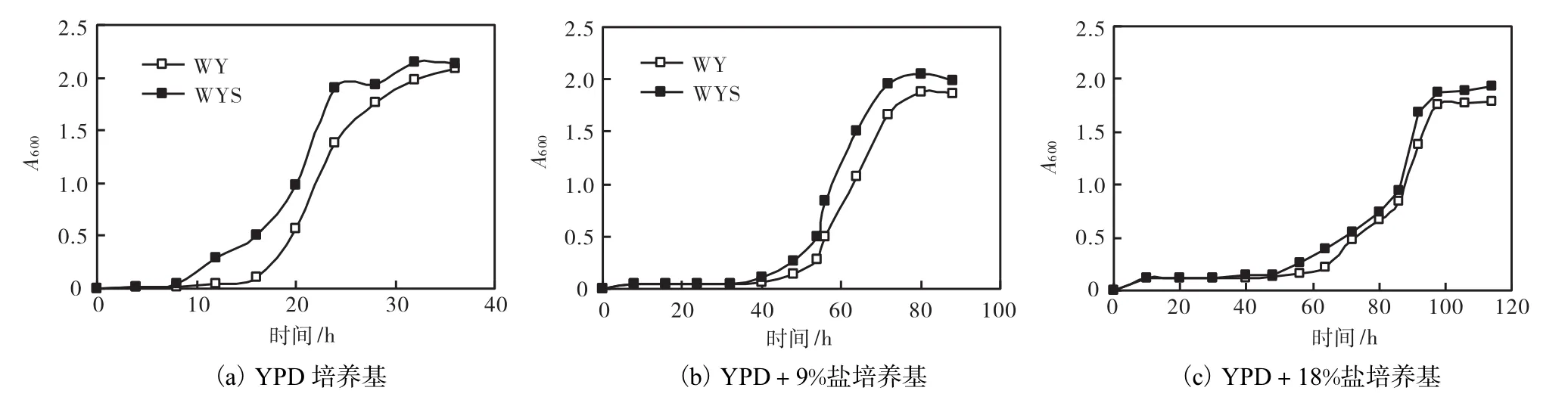
圖3 菌株在不同含鹽量YPD培養基中的生長情況Fig.3 Growth of engineered strains in YPD culture media with different salt concentration
2.3 抗性變化
分別研究菌株 WYS和 WY 對 LiCl、鹽、山梨醇、乙醇和高溫的抗性,結果如圖 4所示,從左到右菌體的濃度依此為 1×106、1×105、1×104、1×103、1×102,μL-1.
由圖 4可見,在 LiCl、NaCl、乙醇不同程度刺激而產生的滲透壓力下[2],WYS生長趨勢明顯優于WY,表現出了較好的耐性;對比圖 4(a)和圖 4(e)發現,(a)組菌落比(e)組菌落大而圓,這說明隨著鹽的加入,導致了細胞內水分的外流,細胞發生了質壁分離現象.而1,mol/L的山梨醇濃度對于該系列菌株來說并無太大影響.42,℃培養對兩株菌株的影響基本相同,只有最高濃度才能生長,但是 WYS的生長優于WY,表明過量表達GPD1對細胞耐受熱具有明顯的作用.

圖4 工程菌株的抗性比較Fig.4 Tolerance comparison of engineered strains
2.4 甘油產量變化
按照 1.2.5方法得到標準曲線 y=0.044,3x+0.001,3,R2=0.999,6.將工程菌株WYS和WY 分別培養在含鹽量為0、9%、18%的YPD培養基中,30,℃培養.當培養至對數后期,測定細胞內外甘油產量,如圖5所示.
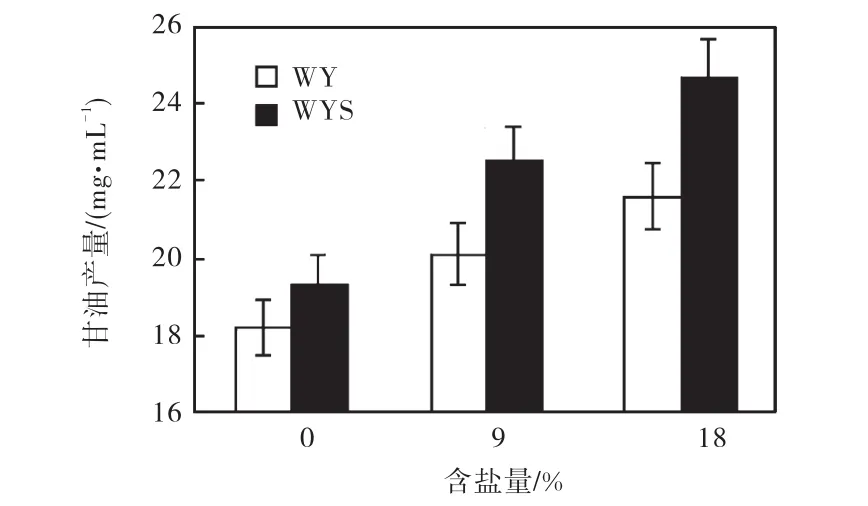
圖5 甘油產量變化Fig.5 Changes of glycerol content
由圖 5可知,隨著含鹽量的增加,WYS和 WY菌株的甘油產量都依次增加,但增加的程度并不一致,這表明它們對環境的應答并不完全一樣.對于同一菌株,隨著含鹽量的增加,其甘油產量也隨之增加;對于同一個含鹽量,WYS菌株的甘油產量高于WY,結合圖 4(e)結果,這說明 WYS中過量表達GPD1使其更能適應外界滲透壓的變化.
3 結 語
基因 GPD1只是耐鹽通路中的一個小分支,要進一步闡明酵母菌株的耐鹽機制,將需要研究此通路中的其他關鍵基因,如FPS1(編碼甘油由胞內向胞外滲透的通道蛋白)、PMA1(編碼質膜 H+-ATPase)、PDC1(編碼丙酮酸脫羧酶)、ALD6(編碼胞質乙醛脫氫酶)等.還需建立起一個立足點較多的基因功能網絡,結合酵母的代謝網絡,對醬油添加酵母的耐鹽機制進行研究.此外,在本研究中只是對過量表達魯氏酵母耐鹽基因 GPD1對釀酒酵母 W303的影響進行了分析,并沒有研究自身過表達 GPD1對耐鹽家族成員魯氏酵母的影響.因此在后續的研究中可以利用質粒 pZEU[10]在魯氏酵母△ura3缺陷型菌株中過量表達耐鹽基因GPD1.
[1] Murooka Y,Yamshita M. Traditional healthful fermented products of Japan[J]. Journal of Industrial Microbiology& Biotechnology,2008,35(8):791-798.
[2] Pribylova L,Montigny J,Sychrova H. Osmoresistant yeast Zygosaccharomyces rouxii:The two most studied wild-type strains(ATCC 2623 and ATCC 42981)differ in osmotolerance and glycerol metabolism[J]. Yeast,2007,24(3):171-180.
[3] Sluis C,Tramper J,Wijffels R H. Enhancing and accelerating flavour formation by salt-tolerant yeasts in Japanese soy-sauce processes[J]. Trends in Food Science &Technology,2001,12(9):322-327.
[4] Pribylova L,Papouskova K,Sychrova H. The salt tolerant yeast Zygosaccharomyces rouxii possesses two plasma-membrane Na+/H+-antiporters(ZrNha1p and Zr Sod2-22p)playing different roles in cation homeostasis and cell physiology[J]. Fungal Genetics and Biology,2008,45(10):1439-1447.
[5] Iwaki T,Higashida Y,Tsuji H,et al. Characterization of a second gene(ZSOD22)of Na+/H+antiporter from salttolerant yeast Zygosaccharomyces rouxii and functional expression of ZSOD2 and ZSOD22 in Saccharomyces cerevisiae[J]. Yeast,1998,14(13):1167–1174.
[6] Iwaki T,Kurono S,Yokose Y,et al. Cloning of glycerol-3-phosphate dehydrogenase genes(ZrGPD1 and Zr GPD2)and glycerol dehydrogenase genes(ZrGCY1 and ZrGCY2)from the salt-tolerant yeast Zygosaccharomyces rouxii[J]. Yeast,2001,18(8):737-744.
[7] Hohmann S. Osmotic stress signaling and osmoadaptation in yeasts[J]. Microbiology and Molecular Biology Reviews,2002,66(2):300-372.
[8] André L,Nilsson A,Adler L. The role of glycerol in osmotolerance of the yeast Debaryomyces hansenii[J].Journal of General Microbiology,1988,134:669-677.
[9] Schiestl R H,Gietz R D. High efficiency transformation of intact yeast cells using single stranded nucleic acids as carrier[J]. Current Genetics,1989,16(5/6):339-346.
[10] Pribylova L,Straub M L,Sychrova H,et al. Characterisation of Zygosaccharomyces rouxii centromeres and construction of first Z. rouxii centromeric vectors[J].Chromosome Research,2007,15(4):439-445.

- 天津科技大學學報的其它文章
- 枯草芽孢桿菌工程菌產耐酸性高溫α-淀粉酶發酵條件的優化