枯草芽孢桿菌工程菌產耐酸性高溫α-淀粉酶發酵條件的優化
2012-05-08 11:51:14劉逸寒徐艷靜薄嘉鑫路福平
天津科技大學學報 2012年6期
胡 博,劉逸寒,徐艷靜,薄嘉鑫,路福平
(工業發酵微生物教育部重點實驗室;天津科技大學生物工程學院,天津 300457)
α-淀粉酶作為淀粉酶的一種,是目前最重要、最古老、應用范圍最廣的工業酶制劑之一,其中耐高溫α-淀粉酶憑借其熱穩定上的優勢已經占據了很大的市場[1].耐高溫α-淀粉酶目前被廣泛地應用于味精、啤酒、有機酸、酒精等食品發酵以及淀粉工業[2-3].目前,國內外市場中常用的耐高溫α–淀粉酶的最適 pH范圍為 6~7[4],在酸性條件下酶活明顯降低,甚至失去活性,已經不能滿足酸性條件下淀粉原料深加工工藝的要求,因此對耐酸性α-淀粉酶的需求日益上升.近幾年來國內開展了大量的針對產耐酸性α-淀粉酶菌株發酵條件優化的研究,王淑軍等[5]對 1株產耐熱酸性α-淀粉酶的超嗜熱古菌進行了發酵條件的優化,經優化后α-淀粉酶活力達到了 500,U/mg;劉雅琴等[6]在篩選出的耐酸性α-淀粉酶產生菌的基礎上對其進行發酵培養基與發酵條件的優化,最終α-淀粉酶活力達到 31.4,U/mL,比初始時提高了 0.65倍;莫新迎等[7]通過單因素實驗和響應面分析法對 1株產耐酸性α-淀粉酶的小孢根霉菌培養基組分進行優化,α-淀粉酶活力較優化前提高了 0.15倍,達到259.902,U/g干物質,但其酶活力均不高.
本實驗通過對本課題組前期研究獲得的可高效表達耐酸性高溫α-淀粉酶枯草芽孢桿菌工程菌株pWB-amyd/WB600[8-10]發酵條件進行優化,旨在提高耐酸性高溫α-淀粉酶活力,為以淀粉為原料的深加工工業的發展提供有力支撐.
1 材料與方法
1.1 菌株
產耐酸性高溫α-淀粉酶工程菌株 pWB-amyd/WB600,由本實驗室構建[9]并保藏.以初始發酵培養基及未優化的發酵條件進行發酵培養,初始酶活力為1,890,U/mL.
1.2 培養基
斜面培養基(g/L):胰蛋白胨 10,酵母浸出粉 5,NaCl 10,瓊脂粉 20,卡那霉素 0.03,pH 自然.
種子培養基(g/L):胰蛋白胨 10,酵母浸出粉 5,NaCl 10,卡那霉素 0.03,pH 自然.
初始發酵培養基(g/L):胰蛋白胨 10,酵母浸出粉 5,NaCl 10,pH 自然.
1.3 培養方法
種子的培養:取凍存于–70,℃的工程菌株 pWB-amyd/WB600,將其接種到新鮮斜面培養基,37,℃培養 18,h后,挑取單菌落接入種子培養基,37,℃、200,r/min搖床培養16,h,得到種子液.
初始發酵培養:以 1%接種量接種至裝液量50,mL發酵培養基的 250,mL三角瓶中,在 37,℃、160,r/min培養36,h.
1.4 生長曲線的繪制
配制 400,mL發酵培養基分裝至 250,mL三角瓶,編號為 1—8,每瓶裝液量 50,mL,121,℃滅菌20,min,待用.從新鮮斜面培養基挑取單菌落接入含有發酵培養基的三角瓶中,于 37,℃、200,r/min搖床振蕩培養.每隔一定時間取樣.將菌液適當稀釋至吸光度在0.1~1.0范圍內,用分光光度計于光程1,cm、波長 600,nm條件下測定其吸光度.以時間為橫坐標,吸光度為縱坐標繪制重組菌生長曲線.
1.5 發酵培養基優化
1.5.1 碳源的選擇
選擇 10,g/L的葡萄糖、麥芽糖、蔗糖、可溶性淀粉、糊精、玉米粉、薯干粉作為碳源,氮源為10,g/L的蛋白胨,比較不同碳源對產酶的影響,確定最適碳源.再以10,g/L的蛋白胨為氮源,加入10、20、30、40、50,g/L的最適碳源,確定最適碳源的最優質量濃度.
1.5.2 氮源的選擇
以最適質量濃度的最適碳源為培養基基本組分,分別加入 10,g/L的胰蛋白胨、蛋白胨、酵母膏、牛肉膏、尿素、NH4NO3、NH4Cl,比較不同氮源對產酶的影響,確定最適氮源.再以最適質量濃度的最適碳源為培養基基本組分,加入 10、20、30、40、50,g/L 的最適氮源,確定最適氮源的最優質量濃度.
1.5.3 金屬離子Ca2+對產酶的影響
以最優質量濃度的最適碳源、氮源為培養基基本組分,加入 0、0.2、0.4、0.6、0.8、1.0,g/L 的 Ca2+進行發酵,確定Ca2+的最優質量濃度.
1.5.4 Na2HPO4對產酶的影響
以最優濃度的最適碳源、氮源、Ca2+為培養基基本組分,加入 0、4、8、12、16,g/L 的 Na2HPO4進行發酵,確定Na2HPO4的最優質量濃度.
1.5.5 正交實驗
利用 L9(34)正交實驗篩選出玉米粉、蛋白胨、CaCl2和Na2HPO4的最佳配比.
1.6 發酵條件優化
1.6.1 培養基初始pH對產酶的影響
以上述優化后的培養基配方為基礎,用一定濃度的 NaOH或 HCl溶液調節培養基初始 pH分別為5.0、5.5、6.0、6.5、7.0、7.5、8.0 進行發酵,確定培養基最適初始pH.
1.6.2 接種量對產酶的影響
以原始發酵培養條件為基礎,分別按 1%、2%、3%、4%和 5%的接種量接種到優化后的最優培養基中,在 37,℃、160,r/min培養 36,h,取樣測定酶活力,確定最適接種量.
1.6.3 裝液量對產酶的影響
按優化后所得的最適接種量將種子液分別接種于裝有 20、30、40、50、60、70,mL 發酵培養基的250,mL三角瓶中,在 37,℃、160,r/min培養 36,h,取樣測定酶活力,確定最適裝液量.
1.6.4 搖床轉速對產酶的影響
將種子液接種到發酵培養基中,接種量和裝液量均按照優化后的結果,在 37,℃條件下,搖床轉速分別為 120、160、200、240,r/min 培養 36,h,取樣測定酶活力,確定最適搖床轉速.
1.6.5 發酵溫度對產酶的影響
將種子液接種到發酵培養基中,接種量、裝液量及搖床轉速均按照優化后的結果,在27、32、37、42,℃分別培養36,h,取樣測定酶活力,確定最適發酵溫度.
1.7 酶活檢測
參照QB/T 2306—1997[11].酶活力單位定義:在70,℃、pH 6.0條件下,1,min液化1,mg可溶性淀粉成為糊精所需要的酶量,即為 1個酶活力單位,以U/mL表示.

式中:X,樣品的酶活力(U/mL);c,測試的酶液濃度(U/mL);n,樣品的稀釋倍數;16.67,換算常數.
2 結果與分析
2.1 生長曲線的測定
工程菌株pWB-amyd/WB600生長曲線如圖1所示.由圖 1可知,0~12,h為生長的延滯期;12~18,h為對數生長期;從18,h開始進入穩定期.由于在對數生長中后期,菌體量較大,此時菌體還未發生自溶,因此確定在接種后18,h時收集菌體.
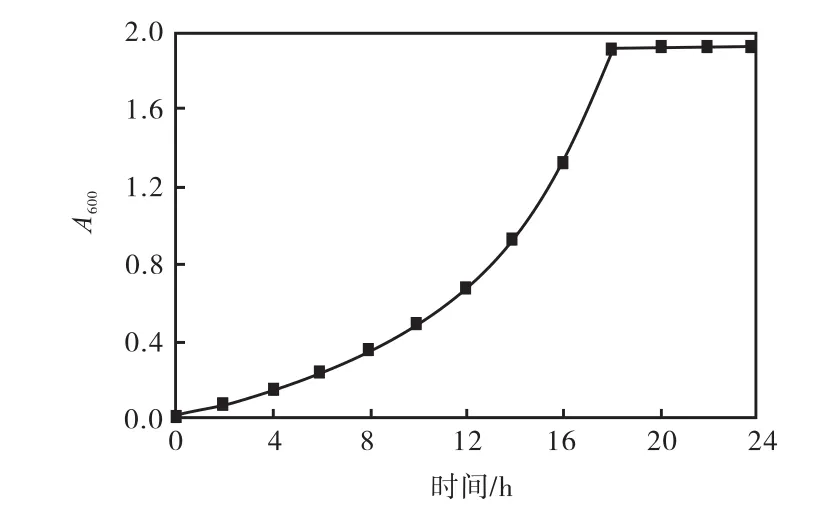
圖1 工程菌株pWB-amyd/WB600生長曲線Fig.1 Growth curve of pWB-amyd/WB600
2.2 發酵培養基的優化
2.2.1 碳源對產酶的影響
初始發酵培養基中無特定的碳源,胰蛋白胨和酵母提取物僅能維持菌體生長至A600為3.0左右[12],為了考察重組菌株對碳源的利用情況,本實驗選擇了幾種常用的碳源進行碳源的優化.由表 1結果可知,玉米粉是產酶的最佳碳源,耐酸性高溫α-淀粉酶活力達到最高值,為 2,090,U/mL,其余依次為可溶性淀粉、糊精、薯干粉、蔗糖、葡萄糖、麥芽糖.玉米粉的最優質量濃度實驗結果表明,隨著玉米粉質量濃度的增加,耐酸性高溫α-淀粉酶活力也隨之增加,當玉米粉質量濃度為 20,g/L時,耐酸性高溫α-淀粉酶活力達到最高,為 2,310,U/mL.當繼續加大玉米粉質量濃度時,其耐酸性高溫α-淀粉酶活力不再隨玉米粉質量濃度的增加而增加,反而隨之下降,所以本實驗選取20,g/L為玉米粉的最適質量濃度.
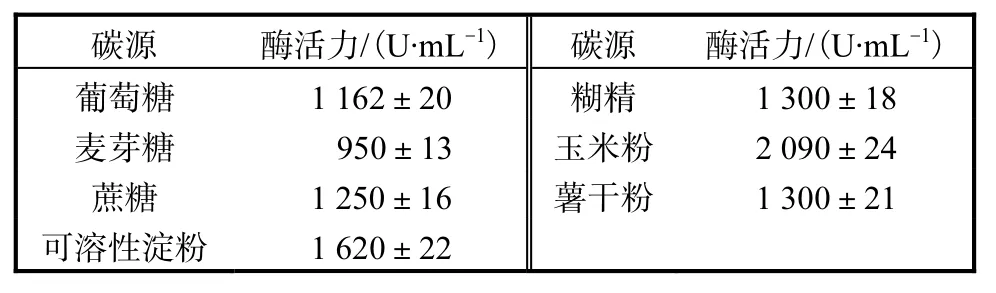
表1 碳源對產酶的影響Tab.1 Effects of carbon resource on the yield of enzyme
2.2.2 氮源對產酶的影響
為了考察重組菌株對碳源的利用情況,實驗中同時對有機氮源和無機氮源進行研究,選擇 10,g/L的胰蛋白胨、蛋白胨、酵母膏、牛肉膏、玉米漿、尿素、NH4NO3、NH4Cl作為氮源,碳源為20,g/L的玉米粉,其他條件相同,結果見表2.
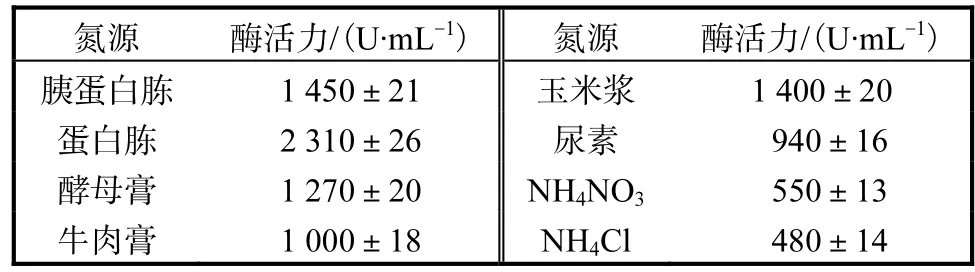
表2 氮源對產酶的影響Tab.2 Effects of nitrogen resource on the yield of enzyme
由表 2可知,重組菌株利用以上氮源培養時,有機氮源對產酶的促進作用明顯優于無機氮源.單一的無機氮源作為培養基中的氮源,菌體生長緩慢,產酶量較低.而有機氮源中,蛋白胨是最佳的氮源,其主要成分是各種氨基酸、肽類等可溶性氮化合物,耐酸性高溫α-淀粉酶活力達到最高值,為2,310,U/mL.蛋白胨最優濃度實驗結果表明,隨著蛋白胨濃度的增加,耐酸性高溫α-淀粉酶活力也隨之增加,當蛋白胨質量濃度為 30,g/L時,耐酸性高溫α-淀粉酶活力達到最高,為2,620,U/mL.所以本實驗選取30,g/L為蛋白胨最適質量濃度.
2.2.3 金屬離子Ca2+質量濃度對產酶的影響
根據 Machius等[13]的研究可知,Ca2+的添加對產地衣芽孢桿菌α-淀粉酶有明顯的促進作用,這是因為產地衣芽孢桿菌α-淀粉酶是一種金屬酶,金屬離子Ca2+與之結合,有利于保持α-淀粉酶的結構穩定性及其活力.故選用不同質量濃度的 Ca2+進行發酵,由圖2中可以看出,隨著Ca2+質量濃度的增加,耐酸性高溫α-淀粉酶活力也隨之增加,當 Ca2+質量濃度為 0.6,g/L時,耐酸性高溫α-淀粉酶活力達到最高,為2,740,U/ mL.當繼續加大Ca2+質量濃度時,其耐酸性高溫α-淀粉酶活力不再隨Ca2+質量濃度的增加而增加,反而隨之下降,所以本實驗選取0.6,g/L為Ca2+最適質量濃度.

圖2 CaCl2質量濃度對產酶的影響Fig.2 Effects of CaCl2concentration on the yield of enzyme
2.2.4 磷酸鹽質量濃度對產酶的影響
磷酸鹽對微生物產酶的作用是非常重要的,磷是細胞合成核酸必需的一種元素,也是加快吸收葡萄糖,促進細胞生長不可或缺的一種物質.所以耐酸性高溫α-淀粉酶活力隨著磷酸鹽質量濃度的增加而增加,當磷酸鹽濃度為8,g/L時,耐酸性高溫α-淀粉酶活力達到最高,為 2,870,U/mL.但是隨著磷酸鹽質量濃度的繼續升高,耐酸性高溫α-淀粉酶活力卻隨之下降,所以本實驗選取 8,g/L為磷酸鹽最適質量濃度(圖 3).
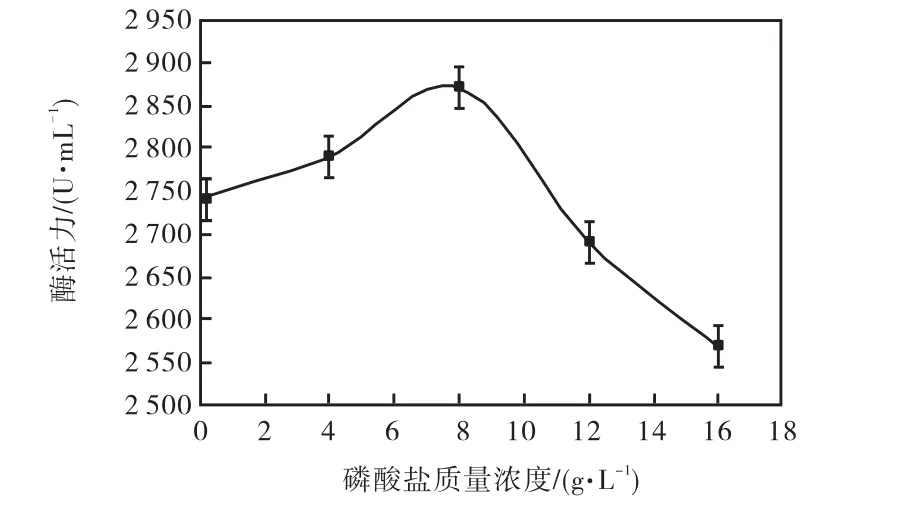
圖3 磷酸鹽質量濃度對產酶的影響Fig.3 Effects of Na2HPO4 concentration on the yield of enzyme
2.2.5 正交實驗
利用 L9(34)正交實驗篩選出玉米粉、蛋白胨、CaCl2和Na2HPO4的最佳配比,結果見表3.
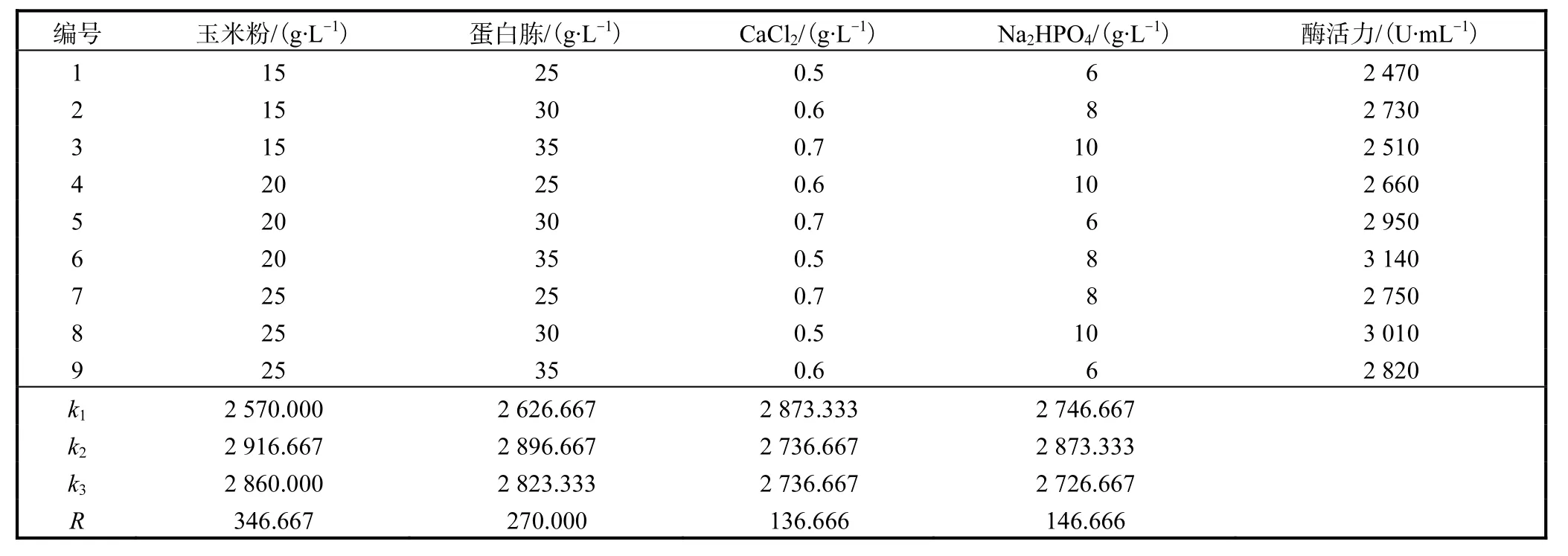
表3 培養基配方正交實驗結果及其極差分析Tab.3 Results and variance analysis of orthogonal experiment of medium components
由表 3可知,在玉米粉、蛋白胨、CaCl2和Na2HPO4這4種培養基組分的不同配比中,耐酸性高溫α-淀粉酶活力最高的組合為(g/L):玉米粉 20,蛋白胨 30,CaCl20.5,Na2HPO48.極差分析結果表明4個因素的影響程度依次為:玉米粉>蛋白胨>Na2HPO4>CaCl2.
由于從理論上優化得到的培養基不在正交實驗表中,經驗證實驗結果表明,耐酸性高溫α-淀粉酶活力達到 3,280,U/mL,此活力大于正交實驗中出現的所有結果,驗證了選取該結果的正確性.因此,確定最適發酵培養基配方為(g/L):玉米粉 20,蛋白胨30,CaCl20.5,Na2HPO48.
2.3 發酵條件
2.3.1 培養基初始pH對產酶的影響
pH是影響微生物正常生理活動的一個重要因素.由圖4結果顯示,發酵培養基初始pH在5.0~8.0之間,都可產生耐酸性高溫α-淀粉酶,初始pH為6.5時,即發酵培養基自然 pH,耐酸性高溫α-淀粉酶活力最高,達到3,280,U/mL.所以本實驗選取pH,6.5為最適初始pH.
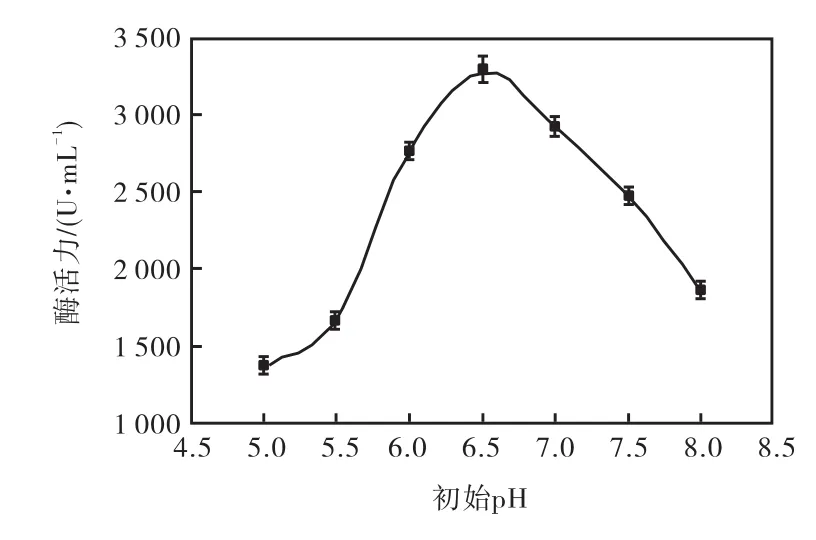
圖4 初始pH對產酶的影響Fig.4 Effects of pH on the yeild of enzyme
2.3.2 接種量對產酶的影響
由圖 5結果顯示,在接種量為 1%時,由于菌體數量過低,減少了蛋白質分泌總量,因此酶活力較低.當以 2%的接種量進行發酵培養時,菌體蛋白分泌較高,酶活力顯著提高,耐酸性高溫α-淀粉酶活力最高,達到 3,450,U/mL.隨著接種量繼續增加,耐酸性高溫α-淀粉酶活力卻隨之下降.根據余曉紅等[14]的研究可知,這是由于培養基中菌體相對密度增大,導致菌體生長時所需培養基成分不足同時溶氧不足,菌體生長狀態不良,反而使酶活力降低造成的,所以本實驗選取2%為最適接種量.
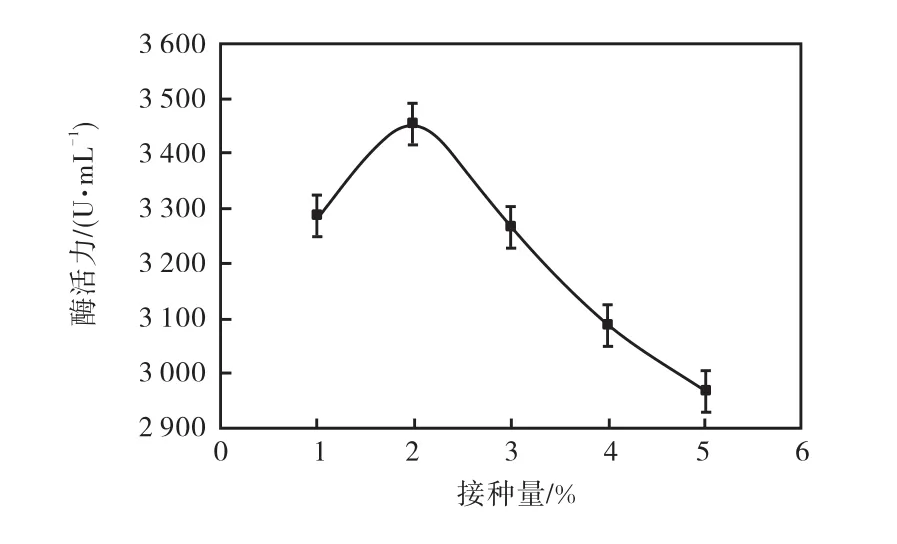
圖5 接種量對產酶的影響Fig.5 Effects of inoculation volume on the yield of enzyme
2.3.3 裝液量對產酶的影響
在微生物發酵過程中,能夠提供充足的氧分對于提高產酶量具有重要的作用,在一定范圍內,裝液量越低,產酶量越高.由圖 6結果顯示,裝液量在30,mL以上時,隨著裝液量的增加,發酵過程中溶氧量不足,菌體生長受到抑制,導致耐酸性高溫α-淀粉酶活力下降.當裝液量為30,mL時,耐酸性高溫α-淀粉酶活力達到最高,為 3,670,U/mL.裝液量為 20,mL時,雖然溶解氧量充足,但是由于養分不足,菌體生長緩慢,導致耐酸性高溫α-淀粉酶活力下降.所以本實驗選取30,mL/250,mL為最適裝液量.
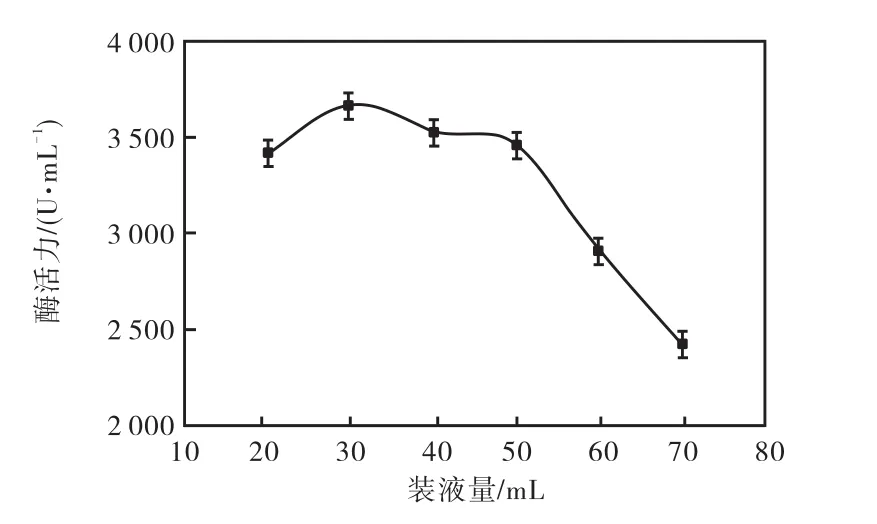
圖6 裝液量對產酶的影響Fig.6 Effects of medium volume on the yield of enzyme
2.3.4 搖床轉速對產酶的影響
搖床轉速同樣影響發酵液中氧氣的溶解量.由表 4結果顯示,當搖床轉速較低時,發酵液中氧氣的溶解量不足,不利于菌體生長,導致耐酸性高溫α-淀粉酶活力下降,因而耐酸性高溫α-淀粉酶活力隨著轉速的升高而變高,當達到200,r/min時,耐酸性高溫α-淀粉酶活力達到最高,為3,980,U/mL.隨著搖床轉速的繼續升高,耐酸性高溫α-淀粉酶活力卻隨之降低,所以本實驗選取200,r/min為最適搖床轉速.
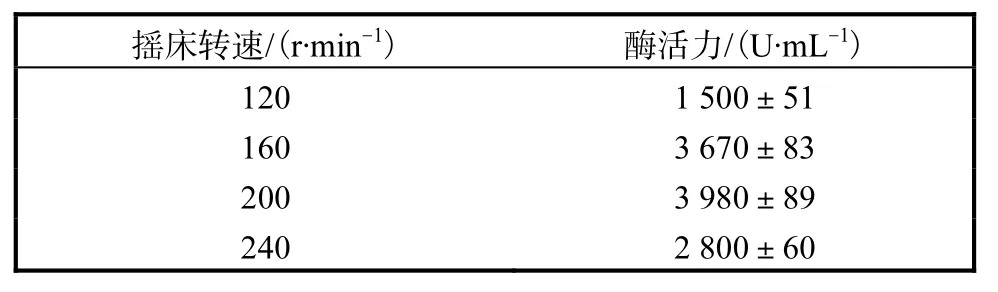
表4 轉速對產酶的影響Tab.4 Effects of rotation speed on the yield of enzyme
2.3.5 溫度對產酶的影響
培養溫度對菌株產酶的影響結果見表5.

表5 培養溫度對產酶的影響Tab.5 Effects of cultivated temperature on enzyme production
由表 5結果顯示,低溫不利于菌株生長產酶,隨著溫度升高,反應速度加大,生長代謝加快,生產期提前,菌株的產酶量大幅度提高,當溫度到達 37,℃時,菌體的生長最旺盛,菌株的產酶活力大幅度增加,耐酸性高溫α-淀粉酶活力達到最高,為 3,980,U/mL.但是隨著溫度的繼續升高使得菌體易于衰老,發酵周期縮短,導致產物的最終產量降低.所以本實驗選取37,℃作為最適發酵溫度.
3 結 論
枯草芽孢桿菌工程菌株 pWB-amyd/WB600,具有高拷貝的重組表達質粒,且具有良好地遺傳穩定性,可高效表達耐酸性高溫α-淀粉酶.本研究在此基礎上,對該工程菌株進行發酵培養基及發酵條件的優化.分別研究碳源、氮源、金屬離子以及磷酸鹽對耐酸性高溫α-淀粉酶活力的影響,通過正交實驗進一步分析,確定最適發酵培養基為(g/L):玉米粉 20,蛋白胨 30,CaCl20.5,Na2HPO48.同時,確定了搖瓶最佳發酵工藝:將重組菌的單菌落接種于種子培養基(50,mL/250,mL三角瓶)中,于 37,℃、200,r/min條件下培養18,h后,以2%接種量接種于優化后的發酵培養基(30,mL/250,mL 三角瓶)中,初始 pH 6.5,37,℃、200,r/min培養 36,h后,最終得到的耐酸性高溫α-淀粉酶活力為3,980,U/mL,是優化前活力的2.1倍.
[1] 胡建恩,曹茜,楊帆,等. 耐高溫α-淀粉酶高密度高表達發酵條件的優化[J]. 食品科學,2012,33(1):219-225.
[2] 張強,黃丹,王川. 耐高溫α-淀粉酶產生菌的選育研究[J]. 中國釀造,2008(7):21-23.
[3] Haki G D,Rakshit S K. Developments in industrially important thermostable enzymes:A review[J]. Bioresour Technol,2003,89(1):17-34.
[4] 劉逸寒,李玉,路福平,等. 耐酸性高溫α-淀粉酶突變基因在大腸桿菌中的表達及酶學性質研究[J]. 食品與發酵工業,2007,33(2):36-41.
[5] 王淑軍,陸兆新,秦松,等. 超嗜熱古菌耐熱酸性α-淀粉酶的發酵條件和酶學性質研究[J]. 海洋與湖沼,2009,40(1):19-26.
[6] 劉雅琴,烏日娜,段金華. 耐酸性α-淀粉酶產生菌的發酵條件優化[J]. 安徽農業科學,2010,38(35):19888-19890.
[7] 莫新迎,高梅瑩,韓丹,等. 耐酸性α-淀粉酶產生菌篩選及培養基優化[J]. 中國釀造,2010(4):85-88.
[8] 劉逸寒,李玉,田琳,等. 耐酸性高溫α?淀粉酶突變基因的異源表達及純化[J]. 化學與生物工程,2007,24(3):58-62.
[9] Liu Yihan,Lu Fuping,Li Yu,et al. Characterisation of mutagenised acid-resistant alpha-amylase expressed in Bacillus subtilis WB600[J]. Applied Microbiology Biotechnology,2008,78:85–94.
[10]Liu Yihan,Lu Fuping,Li Yu,et al. Acid stabilization of Bacillus licheniformis alpha amylase through introduction of mutations[J]. Applied Microbiology Biotechnology,2008,80:795–803.
[11]中國輕工總會. QB/T 2036–1997 耐高溫α-淀粉酶制劑[S]. 北京:中國輕工業出版社,1998.
[12]黃鶴,楊晟,李仁寶,等. 重組青霉素 G 酰化酶在枯草芽孢桿菌中的表達條件優化[J]. 中國生物化學與分子生物學報,2001,17(2):173–177.
[13]Machius M,Declerck N,Huber R,et al. Kinetic stabilization of Bacillus licheniformis α-amylase through introduction of hydrophobic residues at the surface[J]. J Biol Chem,2003,278(13):11546–11553.
[14]余曉紅,汪志君,方維明. 麥汁中啤酒酵母的最適接種量研究[J]. 釀酒科技,2003(4):73-74.

- 天津科技大學學報的其它文章
- 過量表達魯氏酵母耐鹽基因GPD1對釀酒酵母的影響