一株高產低溫纖維素酶南極細菌的篩選、鑒定及酶學性質初步研究
2012-05-07 01:33:50繆錦來
化學與生物工程 2012年2期
高 叢,繆錦來,鄭 洲,金 青
(1.青島科技大學化工學院,山東 青島 266042;2.國家海洋局第一海洋研究所,山東 青島 266061)
纖維素酶廣泛存在于自然界的生物體中,是一種能夠將纖維素降解為葡萄糖的多組分酶系,通常將其分為內切葡聚糖酶、外切葡聚糖酶及β-葡萄糖苷酶,可廣泛用于中藥成分提取、食品、紡織、飼料、釀酒、石油勘探等諸多領域,尤其在生物能源工業中具有重要地位。利用纖維素酶將廢棄纖維類物質糖化,用于制備生物乙醇,緩解能源緊張,是解決未來資源問題的一條重要途徑。
目前,人們對常溫和高溫纖維素酶的研究較多,關于低溫纖維素酶的研究相對較少。低溫酶既可以實現在低溫條件下進行高效酶解反應,同時也可以通過較低溫度的熱處理使酶失活,從而節約能量,降低生產成本。因此,對低溫纖維素酶產生菌的篩選及酶學性質研究已成為熱點。
我國關于南極低溫纖維素酶的研究才剛剛開始。在南極地區,科研人員已在冰雪、土壤及巖石樣品中發現了數量眾多的微生物,并證明微生物在南極自然環境下可以產生大量活性物質(如低溫酶、抗生素、多糖及脂類等),為研究低溫纖維素酶提供了豐富資源。
作者在此從82株南極細菌中篩選出1株高產低溫纖維素酶菌株NJ64,通過16S rDNA序列進行鑒定,并對其產酶條件及酶學性質進行初步研究,以期為南極低溫纖維素酶的開發利用奠定基礎。
1 實驗
1.1 菌株與培養基
南極嗜冷細菌分離于2001年10月~2002年3月在中國第十八次南極科學考察時采集的南極海水水樣和海冰樣品。
發酵培養基為2216E液體培養基,配方為:蛋白胨5 g、酵母粉1 g;用過濾陳海水定容至1000 mL,pH值7.0~7.5,121 ℃下濕熱滅菌20 min。
篩選培養基為添加1%羧甲基纖維素鈉的2216E培養基。
1.2 菌株的篩選
采用剛果紅染色法[1]在篩選培養基上對82株南極細菌進行初步篩選。
1.3 纖維素酶的活性測定
將菌株于10 ℃搖床培養96 h后,取發酵液在7000 r·min-1的條件下離心15 min,取上清液,過濾,得粗酶液。
以羧甲基纖維素鈉為底物,采用DNS法測定纖維素酶活性[2]。在520 nm下用分光光度計測定吸收值。以每分鐘每毫升纖維素酶粗酶液水解羧甲基纖維素鈉產生1 μg還原糖所需要的酶量定義為1個酶活單位(U)。
1.4 菌株的鑒定
DNA模板的制備[3]:將50 μL菌體培養液加入到100 μL預冷的蒸餾水中,在冰上放置1 h以上或者3次凍融循環(-10 ℃/60 ℃)。
采用細菌16S rDNA的通用引物8-27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和1492R(5′-GGTTACCTTGTTACGACTT-3′) 進行PCR擴增[4]。PCR反應條件為:95 ℃,1 min;55 ℃,1 min;72 ℃,1.5 min;循環30次,72 ℃延伸10 min。PCR產物經純化后由華大基因上海鼎安生物科技有限公司進行測序。將所測定的細菌的16S rDNA序列同GenBank數據庫進行相似性比較分析,選取與實驗菌株親緣關系較近的細菌用BioEdit軟件的多序列比對排列(Clustalw multiple alignment)進行序列比對,系統發育分析采用Mega3.1軟件的鄰接法(Neighbor-joining method)進行。
1.5 菌株產酶條件的研究
分別研究培養時間、初始pH值和培養溫度對NJ64產酶的影響。將菌株NJ64接種于發酵培養基中,10 ℃搖床培養120 h,每24 h取發酵液制備粗酶液測定纖維素酶活性;將菌株NJ64分別接種于初始pH值為5.5、6.5、7.5、8.5、9.5及10.5的發酵培養基中,10 ℃搖床培養96 h,測定發酵液中纖維素酶活性;將菌株NJ64接種于發酵培養基中,分別于5 ℃、10 ℃、15 ℃及20 ℃搖床培養96 h,測定發酵液中纖維素酶活性。
1.6 酶學性質初步研究
取10 ℃搖床培養96 h的發酵液,離心后得到粗酶液,分別在0 ℃、10 ℃、20 ℃、30 ℃、40 ℃、50 ℃及60 ℃的條件下進行酶促反應,40 min后用DNS法測酶活,確定酶促反應最適溫度;將粗酶液分別在pH值為6.0、7.0、8.0、9.0、10.0及11.0的條件下進行酶促反應,40 min后用DNS法測酶活,確定酶促反應最適pH值。
2 結果與討論
2.1 菌株篩選結果
采用剛果紅法從82株南極嗜冷細菌中初篩到7株產纖維素酶的細菌。在平板培養基上進行剛果紅染色并脫色后,菌落周圍出現明顯透明圈。將此7株菌進行復篩,用DNS法測定發酵液的酶活性,比較7株菌產酶活性高低,最終確定菌株NJ64為產酶量最高的細菌。
2.2 菌株NJ64分子生物學鑒定結果
將南極嗜冷菌NJ64的16S rDNA的PCR克隆子進行測序,獲得長度為1390 bp的16S rDNA序列,將此16S rDNA序列提交GenBank數據庫,獲得的序列注冊號為:HQ453988。將此序列與GenBank數據庫進行比較。結果表明,與菌株NJ64同源性較高的菌株均為假交替單胞菌屬(Pseudoalteromonassp.),其同源性均大于98.5%。16S rDNA序列同源性小于98%,可以認為屬于不同的種;同源性小于93%~95%,可以認為屬于不同的屬[5,6]。因此,通過16S rDNA序列的比較分析,判斷菌株NJ64應屬于假交替單胞菌屬。用BioEdit軟件進行序列比對,采用Mega3.1軟件的鄰接法構建系統發育樹,結果見圖1。
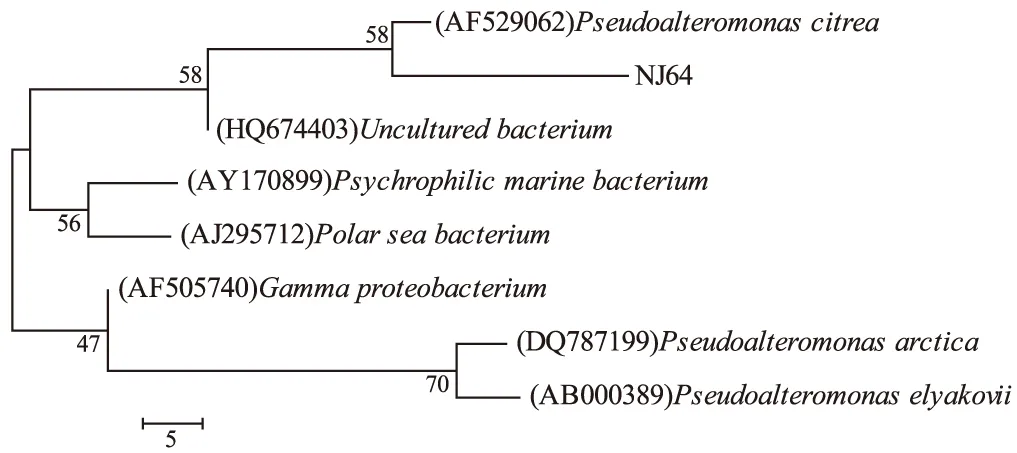
圖1 NJ64系統發育樹
由圖1可知,系統發育分析與16S rDNA的序列同源性比較結果相一致。菌株NJ64屬于交替單胞菌目(Alteromonadales),交替單胞菌科(Alteromonadaceae),假交替單胞菌屬(Pseudoalteromonassp.)。
2.3 菌株NJ64產酶條件的研究
2.3.1 培養時間對NJ64產酶的影響(圖2)
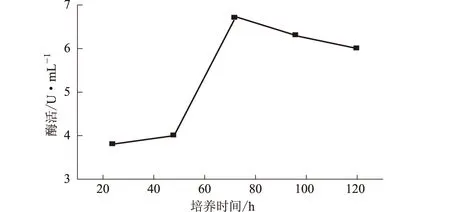
圖2 培養時間對NJ64產酶的影響
由圖2可知,隨著培養時間的延長,纖維素酶活性迅速上升,在72 h時達到最大值,而后酶活性逐漸下降。因此,確定NJ64產酶的最適培養時間為72 h。
2.3.2 初始pH值對NJ64產酶的影響(圖3)
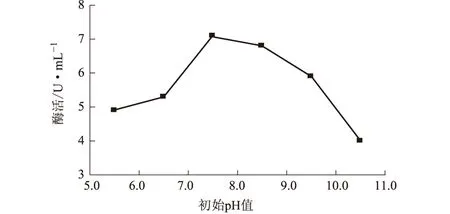
圖3 初始pH值對NJ64產酶的影響
由圖3可知,pH值為7.5時,纖維素酶的活性最高。因此,確定NJ64產酶的最適初始pH值為7.5。
2.3.3 培養溫度對NJ64產酶的影響(圖4)
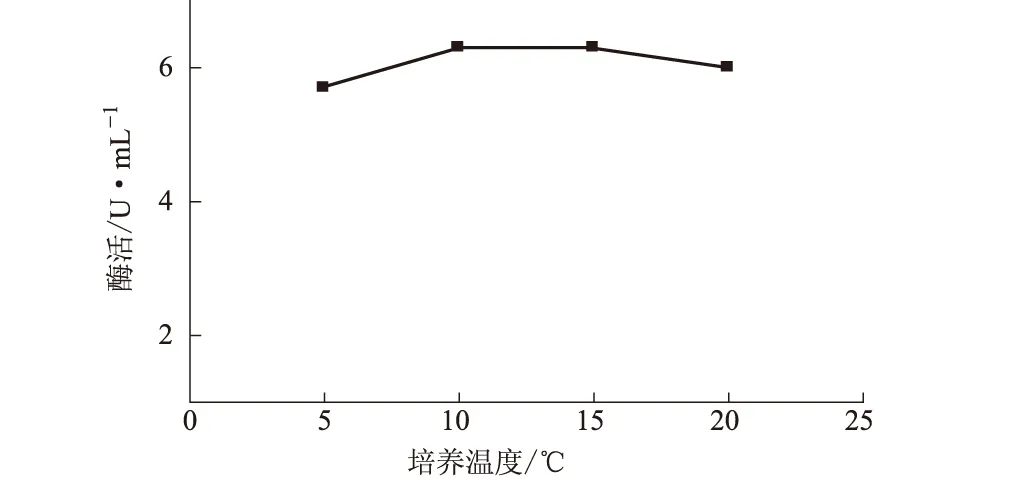
圖4 培養溫度對NJ64產酶的影響
由圖4可知,培養溫度為5~20 ℃時,纖維素酶活性變化不大;10~15 ℃范圍內的酶活性相對較高。因此,確定NJ64產酶的最適培養溫度為10 ℃。
2.4 酶學性質初步研究
2.4.1 酶促反應最適溫度
反應溫度對纖維素酶活性的影響如圖5所示。

圖5 反應溫度對纖維素酶活性的影響
由圖5可知,反應溫度為10~40 ℃時,纖維素酶具有較高的活性,且酶活性隨反應溫度的升高變化較為平緩,均保持在較理想水平;反應溫度高于40 ℃后,纖維素酶活性急劇下降。因此,可以確定該纖維素酶的最適反應溫度為40 ℃,屬于低溫酶[7]。
2.4.2 酶促反應最適pH值
反應pH值對纖維素酶活性的影響如圖6所示。
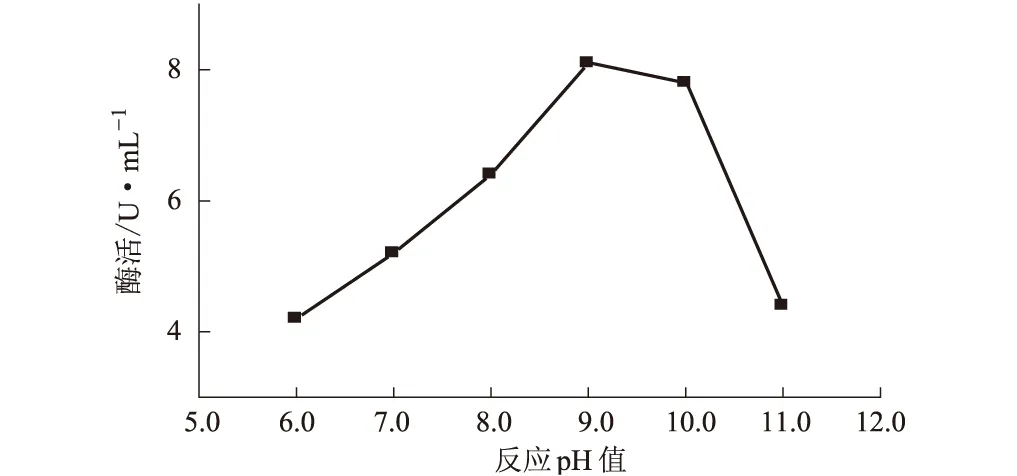
圖6 反應pH值對纖維素酶活性的影響
由圖6可知,pH值為6.0~9.0時,隨pH值的增大,纖維素酶活性逐漸升高;pH值大于9.0后,酶活性急劇下降。因此,可以確定該纖維素酶的最適反應pH值為9.0左右。
3 結論
從82株南極嗜冷菌中篩選分離出1株高產低溫纖維素酶的南極細菌NJ64,經16S rDNA鑒定屬于假交替單胞菌屬(Pseudoalteromonassp.);其在初始pH值7.5、10 ℃的環境中發酵培養72 h時產酶量最高;所產纖維素酶的最適酶促反應pH值為9.0左右、最適反應溫度為40 ℃,為典型的低溫纖維素酶。
目前在利用纖維素酶對廢棄纖維素物質進行糖化時,常用的纖維素酶最適酶促反應溫度(50 ℃左右)高于酵母的發酵溫度(37~40 ℃)[8],而本實驗篩選得到的低溫纖維素酶的酶促反應溫度與酵母的發酵控制溫度范圍較為一致,可以將纖維素糖化與發酵同步進行生產乙醇,具有較好的應用前景。
參考文獻:
[1] 錢文佳,闞光鋒,徐仲,等.產低溫纖維素酶南極細菌的篩選、生長特性及酶學性質的初步研究[J].食品科技,2010,35(1):15-18.
[2] 康紀婷,吳翔,甘炳成,等.纖維素酶活力測定方法[J].河北農業科學,2010,14(4):151-153.
[3] von Stetten F,Francis K P,Lechner S,et al.Rapid discrimination of psychrotolerant and mesophilic strains of theBacilluscereusgroup by PCR targeting of 16S rDNA[J].J Microbiol Methods,1998,34(2):99-106.
[4] Weisburg W G,Barns S M,Pelletier D A,et al.16S Ribosomal DNA amplification for phylogenetic study[J].J Bacteriol,1991,173(2):697-703.
[5] Devereux R,He S H,Doyle C L,et al.Diversity and origin ofDesulfovibriospecies:Phylogenetic definition of a family[J].J Bacteriol,1990,172(7):3609-3619.
[6] Fry N K,Warwick S,Saunders N A,et al.The use of 16S ribosomal RNA analyses to investigate the phylogeny of the familyLegionellaceae[J].J Gen Microbiol,1991,137(5):1215-1222.
[7] 王玢,汪天虹.冷活性纖維素酶性質研究[J].海洋科學,2004,28(2):40-44.
[8] 靳勝英,張禮安,張福琴.纖維質原料制乙醇的關鍵技術[J].化學工業與工程技術,2009,30(2):32-37.
