稻草添補百脈根體外甲烷產量的動態響應
2012-03-13 00:49:42李龍瑞張吉鹍鄒慶華
草業科學 2012年1期
關鍵詞:產量
李龍瑞,張吉鹍,鄒慶華
(1.華南農業大學獸醫學院 廣東 廣州510642;2.江西新天地藥業有限公司獸藥研究院,江西 峽江 331400;3.江西省農業科學院畜牧獸醫研究所 江西 南昌 330200)
稻草是我國南方的主要農作物秸稈,由于其自身營養素的缺乏及硅、木質素等抗營養因子含量較高,不僅使飼喂單一稻草的反芻動物過瘤胃蛋白與生葡萄糖物質水平低,而且稻草在瘤胃內不能很好地被微生物發酵而導致消化率降低,從而不能有效地利用日糧的能量[1]。致使采食單一稻草的反芻動物飼料轉化效率與生產性能低下,而單位畜產品的甲烷產量增加,加劇了當前的溫室效應[2]。近年來,人們將研究的重點轉向具有調控瘤胃發酵功能的植物次生代謝產物[3-5]。在日糧中補飼含皂苷類天然物質如無患子(Sapindussp.)果,可以降低反芻動物瘤胃原蟲數量,增加真菌與細菌的產量,提高飼料利用率與生產性能[6-10]。牧草中的單寧可以增進反芻動物健康,減少甲烷的排放[11]。飼喂低質粗飼料的反芻動物,其生產性能低,主要是因為日糧中,蛋白質缺乏。所以,確保瘤胃中微生物生長所需的適宜氨氮水平是優化瘤胃低質粗飼料發酵的首選[12-13]。對飼喂以稻草為基礎飼料的反芻動物以豆科牧草進行補飼,為瘤胃微生物提供足夠的營養源(可發酵氮源、碳源和一些必須的礦物質元素),利用飼料間的組合效應來改善進入反芻動物體內的營養平衡,促進瘤胃發酵,是提高稻草等秸稈飼料利用率的重要舉措[14-15]。可是,用豆科牧草進行補飼,雖可滿足瘤胃微生物對氮的需求,增加瘤胃中纖維的降解率,但同時又會導致瘤胃甲烷產量的增加[2]。有報道指出,豆科牧草百脈根(Lotuscorniculatus)中高含量的濃縮單寧,能降低粗蛋白在瘤胃的降解率,放牧家畜即使過量采食,也不會發生臌脹病[16]。同時還可以保護過瘤胃蛋白,通過使較多的蛋白質進入小腸消化而提高氮的利用效率,增加肉、乳與毛的產量,使得畜體對環境的氮排泄與甲烷的排放量減少[17-19]。糞臭素與吲哚是由瘤胃微生物對色氨酸的脫氨基與脫羧基作用形成、存在于肉與脂肪中的風味物質,對肉的風味與品質有著負面影響。體外試驗表明,百脈根中的濃縮單寧通過減少瘤胃蛋白的降解來減少糞臭素的產量,從而改善肉質[20]。百脈根葉量豐富,產量高,再生力強,粗蛋白含量高于其他豆科牧草,而粗纖維含量相對較低,特別是其莖稈細軟更令家畜喜食而利用率高。百脈根適口性、營養價值均優于紫花苜蓿(Medicagosativa)等豆科牧草。此外,百脈根的生態適應性強(如南方高溫多雨尤其是在苜蓿易爛根的平原地區),在南方無論是放牧利用還是調制干草或制作青貯都是較優良的豆科牧草[21]。近年來,有關植物次生代謝物調控瘤胃發酵的報道多見于其提取物[22-24],富含植物次生代謝物(如單寧)豆科牧草的過瘤胃蛋白保護作用[25-26]及其中的次生代謝物對瘤胃代謝與飼糧降解率的影響[16],作為含單寧豆科牧草在反芻動物日糧中的適宜比例[27]以及作為添補料對飼喂秸稈基礎飼料綿羊體質量增加的組合效應[28],而有關反芻動物低質秸稈基礎飼料補飼富含植物次生代謝物豆科牧草的瘤胃體外發酵及甲烷產量的報道較少。本研究根據豆科牧草百脈根富含單寧這一特性,就山羊(Caprasp.)稻草基礎飼料補飼百脈根對瘤胃體外發酵參數及甲烷產量的動態變化進行研究。
1 材料與方法
1.1試驗材料 體外批次培養發酵用稻草與百脈根均取自江西新天地乳業現代有機牧場。稻草為收獲稻谷后的晚稻草,百脈根為在開花期刈割(留茬高度8 cm)的整株植株。稻草與百脈根粉碎過粒徑為0.35 mm篩,備作常規化學成分分析與體外發酵。
1.2化學成分分析 試驗所用稻草、百脈根中干物質(DM)、粗蛋白(CP)、粗脂肪(EE)與粗灰分(Ash)的測定依據AOAC[29]的方法進行,中性洗滌纖維(NDF)、酸性洗滌纖維(ADF)與酸性洗滌木質素(ADL)采用Van Soest等[30]的方法進行測定。采用Folin-Ciocalteu比色法測定百脈根干草中的總單寧(Tannin)[31]。體外發酵所用稻草與百脈根的常規營養成分見表1。
稻草-百脈根混合發酵基質營養成分含量=稻草的營養成分含量×稻草比例+百脈根的營養成分含量×百脈根比例。
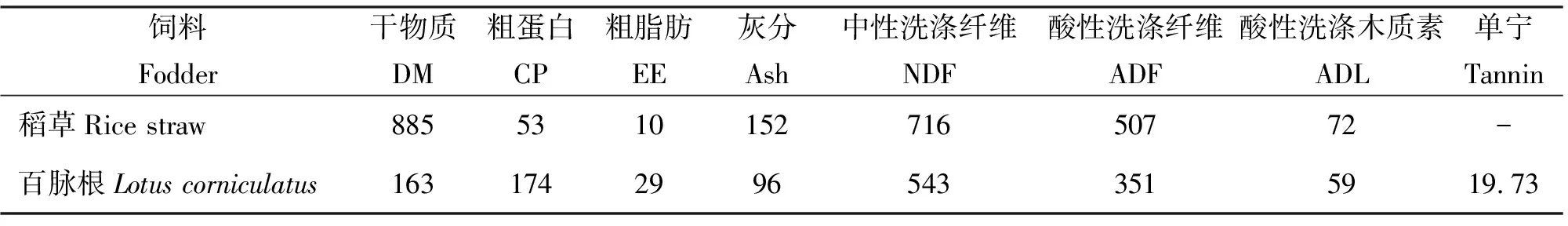
表1 試驗用粗飼料的常規營養成分
1.3試驗設計 本試驗采用單因子11處理重復試驗設計,RS與百脈根分別以100∶0(LC0)、90∶10(LC10)、80∶20(LC20)、70∶30(LC30)、60∶40(LC40)、50∶50(LC50)、40∶60(LC60)、30∶70(LC70)、20∶80(LC80)、10∶90(LC90)與0∶100(LC100)的比例各組成11個組合進行體外發酵,共進行2個批次。在進行批次發酵時,每個樣品設3個重復,同時設1個空白對照組(無發酵樣品的瘤胃液與培養液混合物,3個重復)與1個標準干草組(3個重復)用于校正各時間點因取樣、培養瓶的體積差異以及瘤胃液菌源變異所引起的產氣量變化。
1.4瘤胃液供體動物 選3只體況良好、體質量相近[(33±1) kg]、安裝有永久性瘤胃瘺管的山羊供采集瘤胃液用。日糧的精粗比為3∶7,以混合粗飼料(稻草與百脈根各半)700 g·d-1為基礎飼料,另按干物質計日補充300 g精料。日喂2次(08:00和18:00),自由飲水,常規光照。
1.5體外培養裝置 采用由Theodorou等[32]建立并經Mauricio等[33]改進的壓力傳感器體外產氣技術進行體外批次培養,壓力傳感技術主體裝置由若干產氣瓶、產氣瓶支架、壓力傳感器、帶特定軟件的計算機及恒溫培養箱等組成。每批可同時培養60個樣品。
1.6緩沖液配制 本試驗所用發酵液的配制按照Mauricio等[33]的介紹配制。
1.7體外批次發酵
1.7.1試驗步驟 準確稱取(1 000±10)mg已按要求粉碎的待發酵底物于發酵瓶中,然后加入100 mL緩沖液-瘤胃液混合液(9∶1)。小心排出發酵瓶中的多余氣體,記錄活塞的初始位置,然后將發酵瓶置于39 ℃恒溫培養箱中培養,每個批次培養48 h。分別在發酵的3、6、12、24、36與48 h取發酵液樣測定pH值與氨氮(NH3-N)濃度。
1.7.2測定方法 pH值用pH計(Sartorius PB-20型)直接測定。NH3-N濃度參照馮宗慈和高民[34]的比色方法進行測定。分別記錄培養后3、6、12、24、36與48 h的壓力,每次讀數,取甲烷測定用氣樣后即將瓶內的氣體放掉,將各時間點的壓力轉化為氣體體積即產氣量,然后將各時段的產氣量累加,即為特定時間點全程累計產氣量,48 h產氣量為全程各時間點產氣量之和。甲烷濃度參照Hu等[35]的氣相色譜法測定。
1.8統計分析 用SAS(6.12)軟件的一般線性模型(GLM)程序進行方差(AVOVA)分析和Dancan多重比較。
2 結果
2.1百脈根不同補添水平對發酵液pH值的影響 48 h內,發酵液各時間點pH值隨發酵基質中百脈根添補水平的增加呈降低的趨勢,且降幅有隨添補量增加而加大的趨勢(表2)。各添補組間多數差異顯著(P<0.05)。從時間點分析,各添補組pH值總體上隨培養時間的延長呈下降趨勢,24 h降到最低點,而后有所回升。究其原因,可能是體外批次培養,隨著發酵的進行,揮發性脂肪酸等有機酸(如乳酸)的積累引起pH值下降,同時由于飼料蛋白質分解加速使得氨的濃度增加,導致pH值上升。微生物對氨的利用又會降低氨的濃度,繼而降低發酵液的pH值,而微生物的溶菌與原蟲的自溶又會釋放出氨而增加發酵液的pH值。此外,體外批次培養發酵液的pH值還與所用緩沖液的緩沖能力有關,而發酵基質中添補的豆科牧草中所含的豐富礦物質又會加強這種緩沖能力[2]。這些因素綜合作用,決定著體外發酵液的最終pH值。
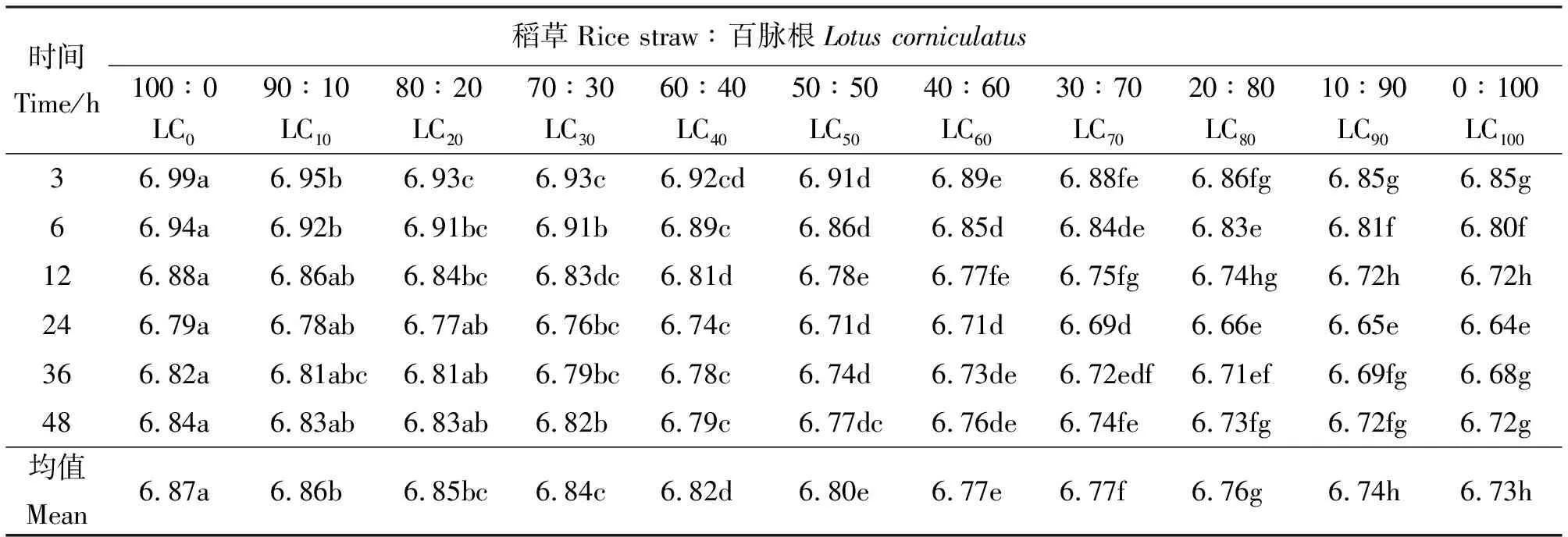
表2 稻草與不同比例百脈根組合體外發酵3~48 h在所測定時間點的pH值
2.2百脈根不同添補水平對發酵液氨氮質量濃度的影響 0~12 h,除12 h的NH3-N質量濃度LC100略低于LC90外,氨氮濃度在各時間點均隨百脈根在混合發酵基質中比例的增加而升高。除12 h的LC100、LC90與LC80組間差異不顯著外(P<0.05),其余各組間差異顯著(P>0.05)(表3),這可能與豆科牧草百脈根較秸稈稻草易在瘤胃降解,微生物發酵百脈根中的含氮物質產生大量氨有關。可見,0~12 h的NH3-N質量濃度基本反映了發酵基質中蛋白質的降解情況。24~36 h,發酵液中NH3-N質量濃度除24 h的 LC10顯著高于LC0外(P<0.05),其余添補組NH3-N質量濃度均隨百脈根在混合發酵基質中比例的增加而降低,可能是因為微生物利用氨合成自身蛋白的速度大于對發酵底物產生氨的速度所致。24 h各組的NH3-N質量濃度差異顯著(P<0.05),但在36 h,LC0與LC10,LC20與LC30,LC30、LC40與LC50以及LC80、LC90與LC100等多個添補組間差異不顯著(P>0.05)。說明在24~36 h,隨著發酵時間的延長,各添補組NH3-N質量濃度差異趨向不顯著(P>0.05)。發酵至48 h,NH3-N質量濃度隨混合發酵基質中百脈根的增加,由LC0的11.19 mg·dL-1降至LC30的8.42 mg·dL-1,隨后又隨百脈根的增加而增加,至LC50的9.13 mg·dL-1,接著又隨百脈根的增加而降低,至LC80的7.24 mg·dL-1,最后又隨百脈根的增加而增加,至LC100的9.57 mg·dL-1。NH3-N質量濃度在整個變化過程中出現了3處拐點,即LC30、LC50與LC80。發酵液中NH3-N質量濃度的回升,可能是細菌溶菌、纖毛蟲自溶或纖毛蟲的吞噬作用釋放出氨所致。

表3 稻草與不同比例百脈根組合體外培養3~48 h NH3-N質量濃度的變化
2.3百脈根不同添補水平對甲烷產量的影響 體外批次發酵的3~12 h,甲烷產量隨百脈根在混合發酵基質中比例的增加而增加,3 h LC0的最低,為0.038 mmol,12 h時LC90和LC100的最高,為0.072 mmol(表4)。隨著發酵的進行,各組合的甲烷產量均緩慢增加,組間差異趨向不顯著。在3 h時,除LC10與LC20、LC40與LC50、LC90與LC100組間差異不顯著外(P>0.05),其余組間差異顯著(P<0.05)。然而,在6 h與12 h,則多數組間差異不顯著(P>0.05)。這可能與本試驗所用發酵基質為稻草與百脈根有關,由于稻草為難以消化的秸稈,瘤胃微生物需要較長時間進行降解,故在發酵初期,甲烷產量較少。百脈根的纖維較稻草的容易降解,使得混合發酵基質甲烷產量隨著百脈根在其中比例的增加而增加,尤其在發酵最初的3 h增加明顯,6~12 h 增加較慢,可能與百脈根中的單寧部分地保護其中的蛋白免受瘤胃微生物的降解有關。發酵至24 h,各組合的甲烷產量均迅速增加,以后緩慢增加直至發酵終點(48 h),甲烷產量變化范圍在1.006~1.313 mmol,最小的為24 h的LC100,最大的為48 h的LC0。24~48 h各組合的甲烷產量,隨百脈根在混合發酵基質中比例的增加而減少,組間差異顯著(P<0.05)。各組甲烷產量均隨發酵時間的延長而增加,尤其在24 h后顯著增加,這與所用發酵基質均為粗飼料有關,通常秸稈類粗飼料在瘤胃中發酵降解需要36~48 h。總單寧含量隨百脈根在混合發酵基質中比例的增加而增加,即LC0、LC10、LC20、LC30、LC40、LC50、LC60、LC70、LC80、LC90與LC100中相當于添加了0、1.973、3.946、5.919、7.892、9.865、11.838、13.811、15.784、17.757與19.730 mg的單寧。這些成比例增長的單寧在一定程度上保護百脈根中蛋白免受瘤胃微生物降解的功能隨混合發酵基質中粗蛋白含量的增加而得到加強,因而甲烷產量隨百脈根在混合發酵基質中比例的增加而減少。
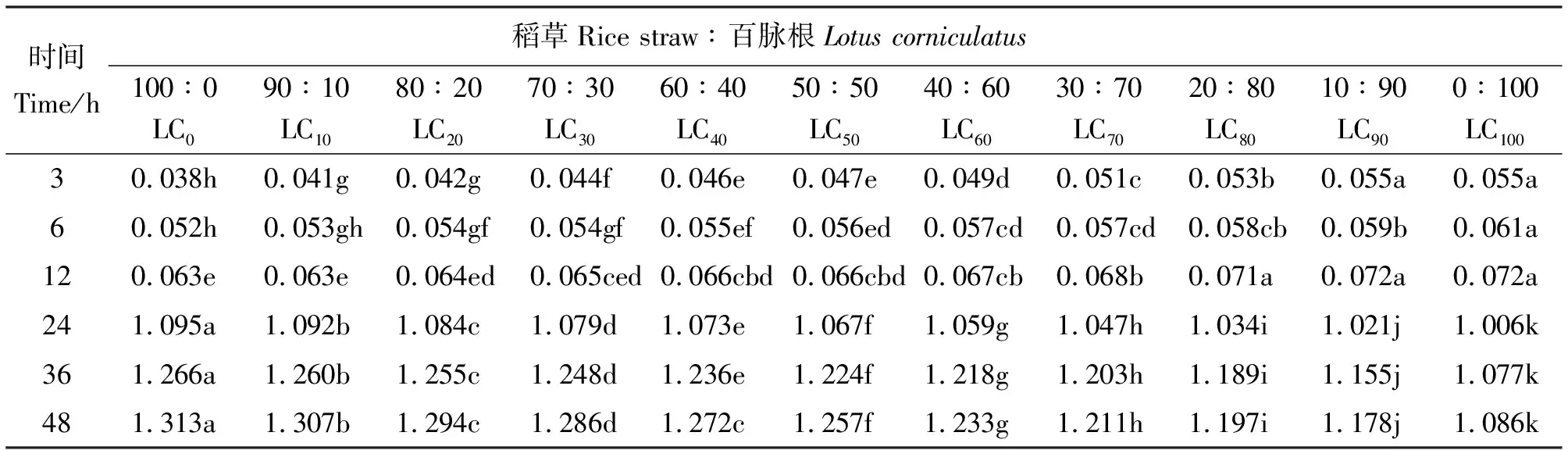
表4 稻草與不同比例百脈根組合體外發酵3~48 h甲烷產量的變化
2.4百脈根不同添補水平對48 h產氣量的影響 各組48 h的產氣量同甲烷一樣,隨百脈根在混合發酵基質中比例的增加而減少,多數組間差異不顯著(P>0.05),而降低的幅度則隨之增加(表5),同樣可能與隨混合發酵基質中百脈根比例增加而增加的單寧有關。
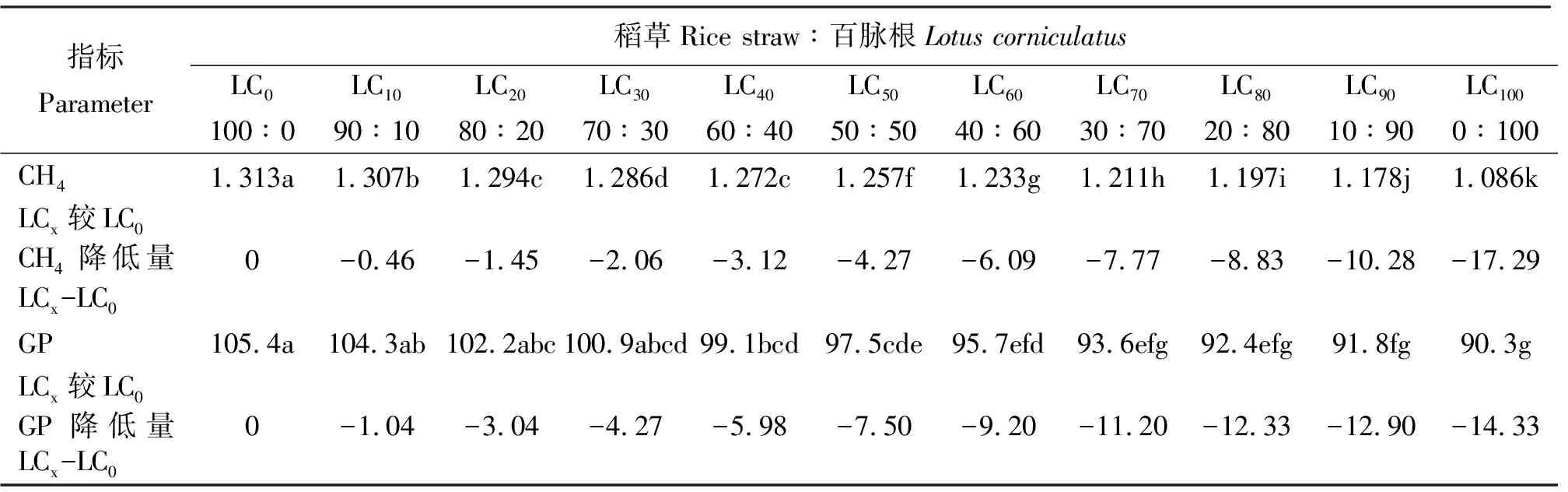
表5 稻草與不同比例百脈根組合體外發酵48 h的總產氣量及甲烷產量
3 討論
3.1百脈根不同補添水平對發酵液pH值的影響 瘤胃是瘤胃微生物賴以生存的直接環境,因此相對穩定的瘤胃內環境是瘤胃微生物正常生長及發揮正常功能的必要條件。瘤胃液pH值是反映瘤胃內環境的重要指標之一,它是飼料在瘤胃內發酵程度與模式的一個表觀特征,是衡量瘤胃發酵狀況的敏感指標,反映了瘤胃的綜合發酵水平。它受唾液分泌量、揮發性脂肪酸及其他有機酸生成量、吸收與排出量等因素的影響。瘤胃液pH值的大幅度波動會影響到瘤胃微生物的合成效率,其波動的原因與動物個體、日糧組成與性質以及環境等因素有關。瘤胃液pH值正常的變化范圍一般在6~7,這是因為過高或過低的pH值對于正常瘤胃微生物生長、發育及底物發酵均有不利的影響。pH值越低,說明發酵所產的酸累積得越多。當pH值低于5.5時,瘤胃MCP合成的效率會受到影響[36]。本試驗所采用的體外批次培養法,因無底物外流與揮發性脂肪酸的吸收以及唾液的緩沖作用,發酵產物大量累積,故各百脈根添補組pH值在所測定的時間點,總體上均隨培養時間的延長而呈現不斷下降趨勢。在培養后的3~24 h,各組發酵液pH值均有所降低,除與此間發酵基質內一些營養物質發酵產酸(主要是揮發性脂肪酸)有關外,亦可能與發酵液中氨被瘤胃微生物吸收利用有關。各百脈根添補組24 h后的pH值又有所回升,這種回升可能與原蟲自溶以及細菌溶菌有一定關系。稻草基礎日糧添補不同水平百脈根的48 h內,所測定發酵液各時間點pH值的變動范圍在6.64~6.99,在瘤胃微生物特別是纖維分解菌生長所需的適宜pH值范圍(6.2~7.0)內[37]。這說明本體外發酵試驗沒有改變瘤胃內環境,對瘤胃發酵沒有產生不利影響。同時亦表明,本研究所用緩沖液緩沖能力較強,足以維持培養液的pH值在正常范圍之內。段智勇[38]采用相同緩沖液研究稻草NDF添加不同水平的玉米(Zeamays)淀粉組合效應時,測得的稻草NDF和玉米淀粉混合物培養液24 h的pH值在6.68~6.84,并沒有隨玉米淀粉含量的增加與培養時間的延長而顯著降低。Sommart等[39]研究稻草或5%尿素處理稻草添補不同水平的木薯(Manihotesculenta)對體外微生物發酵特性與微生物蛋白合成的影響時,發現盡管pH值受到木薯的添加水平及粗飼料的影響,且隨培養時間的延長而顯著降低,但pH值在6.70~6.88。本研究與這些研究結果相似。
3.2百脈根不同添補水平對發酵液氨氮濃度的影響 瘤胃液氨氮質量濃度是瘤胃內環境參數中反映氮代謝的一個重要指標,主要取決于日糧含氮物質降解速度與微生物對氨利用速度的動態平衡,其濃度基本可以反映出日糧蛋白質的降解情況與微生物的合成效率[40]。瘤胃內NH3-N既是瘤胃氮代謝過程中外源(飼料)蛋白質,內源含氮物質降解的重要產物,同時也是在有能量和碳鏈的情況下,瘤胃微生物合成菌體蛋白的原料。Clark等[41]通過體外批次發酵研究證明,在瘤胃微生物區系中,約80%的瘤胃細菌以NH3作為生長的唯一氮源,約50%的細菌可以以NH3-N或氨基酸作為生長的氮源,另26%的細菌的生長離不開NH3-N。因此,瘤胃內NH3-N濃度是瘤胃內環境參數的重要指標,是瘤胃內飼料蛋白降解及微生物對NH3-N利用的綜合反映,故保證最佳的瘤胃NH3-N濃度是獲得最大瘤胃微生物蛋白合成量的關鍵因素。體外批次發酵體系中的NH3-N濃度主要取決于3個因素。首先是發酵基質中的蛋白質含量及其降解率與理化特性;其次是含氮物質與能量載體物質的比例,能氮釋放是否同步;再次為MCP的合成效率與微生物自溶。這3個因素關系緊密、相互影響。NH3-N濃度過低會影響微生物蛋白產量,過高表明氨釋放速度高于其利用與吸收速度,造成氨的損失。Owens和Bergen[42]指出,瘤胃內微生物蛋白合成所需的NH3-N濃度為0.35~29 mg·dL-1,本研究的NH3-N濃度為3.92~26.23 mg·dL-1。一些學者就適于微生物生長的最佳NH3-N濃度作了較為深入的探討,Schaefer等[43]報道,體外培養液中滿足微生物生長需要的理想NH3-N濃度為2~5 mg·dL-1。Petersen[44]報道,放牧反芻動物瘤胃液中NH3濃度在1~2 mg·dL-1時,就可滿足細菌對纖維降解的需要。Clark等[41]證實奶牛瘤胃液中的NH3-N濃度在2 mg·dL-1時,就能滿足瘤胃微生物合成蛋白的需要。可見瘤胃微生物以氨作氮源合成瘤胃微生物蛋白的效率很高,即使瘤胃液中的氨態氮很低,亦能為微生物利用[43]。因此,有理由認為本研究中的氨氮濃度足以滿足瘤胃微生物生長的需要,有效能(稻草纖維的有效降解)成為合成微生物蛋白的主要制約因素。
3.3百脈根不同添補水平對產氣量及甲烷產量的影響 瘤胃發酵所產氣中最主要的氣體為CO2與CH4,分別占總量的70%與30%左右。產氣量是一個綜合反映飼料中有機物可發酵程度的指標,表現了瘤胃微生物活動的總體趨勢[13]。飼料中糖(可發酵纖維)的產氣量要高于蛋白質的產氣量,因此,產氣量同時也是反映飼料蛋白價值的綜合指標[14]。飼料的可發酵有機物含量越多,瘤胃微生物的活性越高,產氣量就越大;飼料中的可發酵有機物含量越少,瘤胃微生物的活性越弱,產氣量也越少[41]。產氣量 (或有機物降解率)主要反映的是飼料中可利用能量的多少,而瘤胃微生物蛋白反映的是飼料提供微生物蛋白能力的大小,只有把這2個指標結合起來,進行綜合的整體評定[45],才能較為準確地評定稻草與百脈根的組合效應。本研究中,48 h的產氣量隨百脈根在混合發酵基質中比例的增加而減少,而瘤胃微生物蛋白則隨百脈根在混合發酵基質中比例的增加而增加[46],表明提高混合發酵基質中百脈根的比例,可以使得混合發酵基質的能氮平衡趨于合理。本研究所報道的產氣量及甲烷產量,隨混合發酵基質中百脈根(單寧)比例的增加而下降,且下降的幅度亦隨之增加。王麗鳳[47]用不同的甲烷抑制劑進行體外抑制甲烷的試驗,亦發現甲烷產量隨著不同甲烷抑制劑在混合發酵基質中比例的增加而減少。胡偉蓮[23]用皂苷進行體外抑制甲烷的試驗,亦得出了同樣的結論。此外,本研究體外發酵的甲烷產量在段志勇[38]與胡偉蓮[23]報道的范圍之內。
4 結論
山羊稻草基礎日糧添補不同水平百脈根均可降低山羊瘤胃體外發酵甲烷產量,可以根據實際需要進行補飼。
[1]張吉鹍,李龍瑞,鄒慶華.稻草與不同飼料混合在體外消化率上的組合效應研究[J].草業科學,2010,27(11):137-144.
[2]Diaz A,Avendano M,Escobar A.Evaluation ofSapindussaponariaas a defaunating agent and its effects on different ruminal digestion parameters[J].Livestock Research for Rural Development,1993(5):1-6.
[4]Wallace R J,McEwan N R,McIntosh F M,etal.Natural products as manipulators of rumen fermentation[J].Asian-Australasian Journal of Animal-Science,2002,15,1458-1468.
[5]Wina E,Muetzel S,Becker K.The impact of saponins or saponincontaining plantmaterials on ruminant production ——a review[J].Journal of Agricultural Food Chemistry,2005,53:8093-8105.
[6]Hess H D,Lascano C E,Carulla J E,etal.Potential of forage legumes and of saponin-containing fruits as tropical feed resources to manipulate rumen fermentation and to improve ruminant nutrition[J].Tropical and Subtropical Agroecosystems,2003:555-559.
[7]Ningrat R W S,Garnsworthy P C,Newbold C J.Saponin fractions inSapindusrarak:effects on rumen microbes[J].Reproduction Nutrition Development,2002,42(suppl.1):82.
[8]Thalib A,Widiawati Y,Hamid H,etal.The effects of saponin fromSapindusrarakfruit on rumen microbes and performance of sheep[J].Journal Ilmu Ternak dan Veterinary,1996,2:17-20.
[9]Wina E.The utilization ofSapindusrarakDC.Saponins to improve ruminant production through rumen manipulation[D].Stuttgart,Germany:University of Hohenheim,2005.
[10]Wina E,Muetzel S,Hoffmann E,etal.Saponins containing methanol extract ofSapindusrarakaffect microbial fermentation,microbial activity and microbial community structureinvitro[J].Animal Feed Science and Technology,2005,121:159-174.
[11]Waghorn G C,McNabb W C.Consequences of plant phenolic compounds for productivity and health of ruminants[J].Proceedings of the Nutrition Society,2003,62:383-392.
[12]Leng R A.Factors affecting the utilization of‘ poor-quality’ forages by ruminants particularly under tropical conditions[J].Nutrition Research Reviews,1990(3):277-303.
[13]張吉鹍,鄒慶華,鐘小軍.稻草添補矮象草體外發酵組合效應的綜合評定研究[J].中國畜牧雜志,2008,44(21):38-41.
[14]張吉鹍,劉建新.用壓力傳感器產氣技術評定玉米秸稈添補苜蓿的組合效應[J].中國畜牧雜志,2007,43(21):40-43.
[15]張吉鹍.飼料間的組合效應及其在配方設計中的應用[J].草業科學,2009,26(12):113-117.
[16]張曉慶,郝正里,李發弟,等.紅豆草縮合單寧對綿羊瘤胃代謝及飼糧尼龍袋降解率的影響[J].草業學報,2010,19(1):166-172.
[17]Barry T N,McNabb W C.The implications of condensed tannins on the nutritive value of temperate forages fed to ruminants[J].British Journal of Nutrition,1999,81:263-272.
[18]Min B R,Barry T N,Attwood G T,etal.The effect of condensed tannins on the nutrition and health of ruminants fed fresh temperate forages: a review[J].Animal Feed Science and Technology,2003,106:3-19.
[19]Woodward S L,Waghorn G C,Ulyatt M J,etal.Early indication that feeding lotus will reduce methane emission from ruminants[J].Proceedings of the New Zealand Society of Animal Production,2001,61:23-26.
[20]Schreurs N M,Tavendale M H,Lane G A,etal.Effect of white clover (Trifoliumrepens),perennial grass (Loliumperenne) andLotuscorniculatusoninvitroskatole and indole formation[A].In:Stockdale R,Heard J,Jenkin M.Animal Production in Australia,Proceedings of the 25th Biennial Conference of the Australian Society of Animal Production[C].Melbounce,Australia:CSIRO Publishing,2004:164-167.
[21]張鴨關,匡崇義,陳功.云南引進帝國百脈根的研究[J].四川草原,2004(12):9-11.
[22]王雅倩,趙國琦,閆韓韓,等.利用體外法研究不同水平苜蓿皂苷對山羊瘤胃內pH值和揮發性脂肪酸的影響[J].飼料工業,2009,30(3):35-40.
[23]胡偉蓮.皂苷對瘤胃發酵與甲烷產量及動物生產性能影響的研究[D].杭州:浙江大學,2005.
[24]Yuan Z P,Zhang C M,Zhou L,etal.Inhibition of methanogenesis by tea saponin and tea saponin plus disodium fumarate in sheep[J].Journal of Animal and Feed Sciences,2007,16:560-565.
[25]牛菊蘭,馬文生.紅豆草中單寧對過瘤胃蛋白的保護研究[J].草業科學,1995,12(3):60-65.
[26]張曉慶.紅豆草中單寧對飼糧蛋白質保護的效果[D].蘭州:甘肅農業大學,2005.
[27]張曉慶,李勇,李發弟,等.紅豆草單寧含量對綿羊養分消化率及氮利用的影響[J].畜牧獸醫學報,2009,40(3):356-362.
[28]Ben Salem H,Makkar H P S,Nefzaoui A,etal.Benefit from the association of small amounts tannin-rich shrub foliage (AcaciacyanophyllaLindl) with soya bean meal given as supplements to Barbarine sheep fed on oaten hay[J].Animal Feed Science and Technology,2005,122:173-186.
[29]AOAC.Official Methods of Analysis. Association of Official Analytical Chemists[M].15th Ed.Washington, DC,USA:Association of Official Analytical Chemists,1990.
[30]Van Soest P J,Robertson J B,Lowis B A.Methods for dietary fiber,neutral detergent fiber,and nonstarch polysaccharides in relation to animal nutrition[J].Journal of Dairy Science,1991,74:3583-3597.
[31]Horvath P J.The nutritional and ecological significance of acer-tannins and related polyphenols[D].Ithaca,N.Y.P:Cornell University,1981:138.
[32]Theodorou M K,Williams B A,Dhanoa M S,etal.A simple gas production method using a pressure transducer to determine the fermentation kinetics of ruminant feeds[J].Animal Feed Science and Technolgy,1994,48:185-197.
[33]Mauricio R M,Mould M L,Dhanoa M S,etal.A semi-automatedinvitrogas production technique for ruminant feedstuff evaluation[J].Animal Feed Science and Technolgy,1999,79:321-330.
[34]馮宗慈,高民.通過比色測定瘤胃液氨氮含量方法的改進[J].內蒙古畜牧科學,1993(4):40-41.
[35]Hu W L,Wang J K,Lv J M,etal.Rapid gas chromatogram determination of methane,organic acid ininvitroruminal fermentation products[J].Journal of Zhejiang University(Agriculture & Life Science),2006,32(2):217-221.
[36]Calsamiglia S,Ferret A,Devant M.Effects of pH and pH fluctuations on microbial fermentation and nutrient flow from a dual-flow continuous culture system[J].Journal of Dairy Science,2002,85:574-579.
[37]Mould F L,?rskov E R,Mann S O.Associative effects of mixed feeds.I.Effects of type and level of supplementation and the influence of the rumen fluid pH on cellulolysisinvivoand dry matter digestion of various roughages[J].Animal Feed Science and Technolgy,1983,10(1):15-30.
[38]段智勇.反芻動物日糧中淀粉與纖維的組合效應及其機理的研究[D].杭州:浙江大學動物科學學院,2006.
[39]Sommart K,Parker D S,Rowlinson P,etal.Fermentation characteristics and microbial protein synthesis in aninvitrosystem using cassava, rice straw and dried ruzi grass as substrates[J].Asian-Australasian Journal of Animal-Science,2000,8:1084-1093.
[40]張吉鹍.粗飼料分級指數參數的模型化及粗飼料科學搭配的組合效應[D].呼和浩特:內蒙古農業大學,2004.
[41]Clark J H,Klusmeyer T H,Cameron M R.Nitrogen metabolism and amino acid nutrition in dairy cattle:Microbial protein synthesis and flows of nitrogen fractions to the duodenum of dairy cows[J].Journal of Dairy Science,1992,75:2304-2323.
[42]Owens F N,Bergen W G.Nitrogen metabolism of ruminant animals:Historical perspective,current understanding and future implications[J].Journal of Animal Science,1983,57 (Suppl.2):498.
[43]Schaefer D M,Davis C L,Bryant M P.Ammonia saturation constants for predominant species of rumen bacteria[J].Journal of Dairy Science,1980,63:1248.
[44]Petersen M K.Nitrogen supplementation of grazing liverstock[A].Proceeding Grazing Liverstock Nutrition Conference[C].Jackson:University of Wuoming,1987:115-122.
[45]張吉鹍,鄒慶華,王金芬,等.稻草與多水平苜蓿混合瘤胃體外發酵組合效應的整體研究[J].飼料工業,2011,32(17):40-48.
[46]張吉鹍,鄒慶華,王金芬,等.稻草補飼百脈根瘤胃體外發酵及微生物蛋白合成的組合效應研究[J].江西農業大學學報,2011, 33(5):0942-0948.
[47]王麗鳳.日糧中添加甲烷抑制劑對綿羊瘤胃中甲烷產量影響的研究[D].呼和浩特:內蒙古農業大學,2004.
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
