外源多胺對薄皮甜瓜花芽分化及花發育的影響
2012-02-23 06:17:34
中國蔬菜 2012年10期
(沈陽農業大學園藝學院,設施園藝省部共建教育部重點實驗室,遼寧省設施園藝重點實驗室,遼寧沈陽110866)
多胺是生物體代謝過程中產生的具有調控作用的低分子量脂肪族含氮堿,包括腐胺(Put)、亞精胺(Spd)、精胺(Spm)和尸胺(Cad)等。它們廣泛存在于各種植物中,與植物的生長發育密切相關。植物的性別分化與激素和多胺有關,已經在很多植物上得到證明。外源多胺能顯著促進紅富士蘋果花芽分化(Altamura & Tomassi,1998),其中Put 被認為是形成花的主要多胺。也有人認為,Put 對板栗結實花分化有顯著抑制作用,而Spd 對結實花分化有促進作用(張新生等,2005)。在油菜中發現較低含量的多胺(主要是Put 和Spd)有利于油菜花芽分化的起始,含量的增加有利于花芽的發育,抽薹越早的品種其體內多胺含量越早達到較高水平(艾育芳 等,2011)。開花時期,在花的萼片、花瓣和性器官中,多胺水平上升,子房中積累大量的Spm(de Dios et al.,2006)。高含量的Cad 有利于甜瓜雄蕊的成熟,高含量的Spd 可能有利于早期雌蕊的發育(張建農和李計紅,2007)。在盛花期之前噴施Spm 可以有效地延緩花的衰老(Seo et al.,2009)。目前,外源多胺對甜瓜花芽分化進程和時期的影響還不明確,本試驗通過外源噴施多胺,研究甜瓜葉片和花芽中內源多胺和激素水平的變化,以期從生理水平上揭示外源多胺對內源多胺和植物激素及甜瓜花芽分化的影響,也為甜瓜性別分化外源調控提供依據。
1 材料與方法
1.1 試驗材料
供試材料為薄皮甜瓜(Cucumis meloL.var.makuwaMakino)永甜3 號,來自黑龍江省齊齊哈爾市永和甜瓜經濟作物研究所。
1.2 試驗方法
2008年8月播種,穴盤基質育苗,每盤50 株。8月14日子葉展開時開始噴施外源多胺(Sigma公司),設4 個處理:T1,Put 1×10-3mol·L-1;T2,Put 1×10-4mol·L-1;T3,Spd 1×10-3mol·L-1,T4,Spd 1×10-4mol·L-1。以噴清水為對照(CK)。每處理3 盤,3 次重復。以后每7 d 噴施1次,共噴3 次。子葉展開當天開始取樣,每隔5 d 取1 次樣。當甜瓜幼苗4 片真葉時,于9月4日定植于高效節能日光溫室,隨機區組排列,每5 畦為1 個小區,3 次重復,高畦雙行栽培,株距40 cm,黑色地膜覆蓋,滴灌,三蔓整枝,爬地栽培,田間管理與生產相同。
1.3 調查項目
1.3.1 花芽分化狀態和時期的顯微結構觀察、結實花花期及第1 節位結實花率調查 子葉展開時開始取樣(張多嬌 等,2009),每隔5 d 取1 次樣,常規石蠟切片法觀察花芽分化狀態與時期。結實花開花日期和節位:結實花開花當天采用防落素噴花,并掛標簽記錄開花日期及節位,統計第1 結實花開花日期、初花期(30%結實花開放)和盛花期(70%結實花開放)。
1.3.2 內源激素和內源多胺的測定 子葉展開時開始取樣,每隔5 d 取1 次樣,取花芽和幼葉1.0 g,液氮速凍后貯藏在-80 ℃冰柜中。GA3、IAA、ZT 和ABA 等內源激素含量測定采用高效液相色譜法(張建農和李計紅,2007)。色譜條件:色譜柱Spherisorb C18,柱溫為25 ℃,洗脫液為甲醇∶水=64V∶36V,流速0.5 mL·min-1,紫外檢測器,檢測波長230 nm,進樣量20 μL,外標法定量。提取方法:稱取植物材料0.5 g,加入2 mL 預冷的5% HClO4研磨,研磨混合液在冰浴中放置1 h 后離心(0~4 ℃,48 000×g,20 min)。取上清液1 mL,加入2 mol·L-1的NaOH 2 mL 和苯甲酰氯20 μL,渦漩20 s 后30 ℃下保溫60 min;加入飽和NaCl 2 mL 和乙醚2 mL 混合萃取苯甲酸多胺,振蕩后1 500×g 離心5 min。收集1 mL 醚相,熱氣流(50 ℃)蒸干,-20 ℃保存。高效液相色譜測定前用100 μL 甲醇溶解樣品,取20 μL 進樣。
用高效液相色譜法(HPLC)測定Spm、Spd 和Put 含量(郝建軍和劉廷吉,2001)。
試驗數據采用Excel、DPS 軟件進行相關分析,Duncan’s 多重差異顯著性分析。
2 結果與分析
2.1 外源多胺對薄皮甜瓜花芽分化進程的影響
外源多胺對薄皮甜瓜花芽分化進程的影響見表1。8月14日未進行多胺處理之前,各處理均為子葉展開,石蠟切片觀察,花芽未分化。經過5 d,8月19日取樣時,即噴施1 次多胺后,各處理仍相同,均處于花芽分化初期。8月24日后,各處理花芽分化進程產生差距,表現為T1和T4 處理均進入花冠裂片分化期;T3 處理次之,進入萼片分化期;而T2 處理和CK 仍停留在花芽分化初期。到9月3日時,仍是T1 處理花芽分化最快,已經發育形成結實花;T4 處理其次,進入雌蕊分化期;然后是T3 處理,CK 和T2 處理花芽分化最慢。
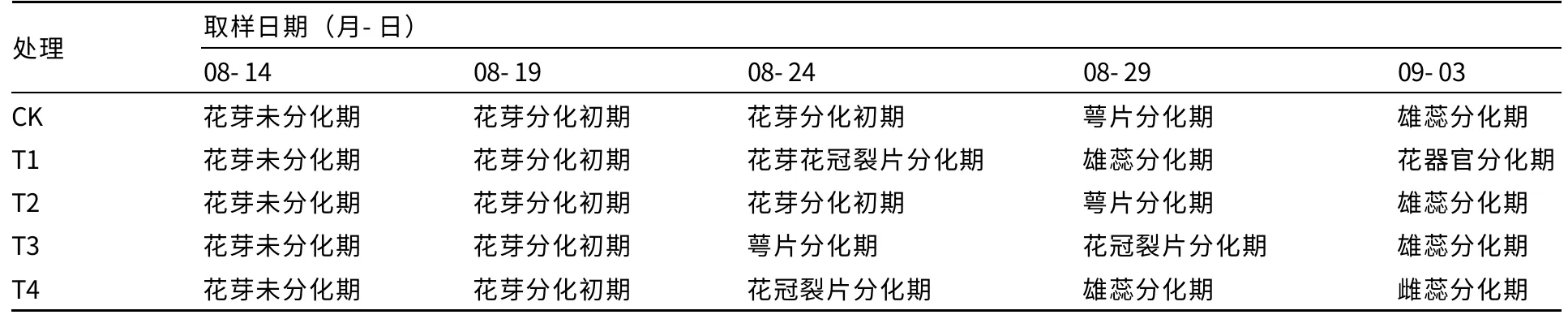
表1 外源多胺對薄皮甜瓜花芽分化進程的影響
2.2 外源多胺對薄皮甜瓜結實花花期和子蔓第1 節位結實花率的影響
由表2 可以看出,經過外源多胺處理后,T1 和T4 處理的花期基本相同或相近,第1 結實花開花期比CK 和其他兩個處理均早1~2 d;T2 處理花期最晚,盛花期比T1 處理晚6 d。CK與T3 處理第1 結實花開花期、初花期、盛花期均相同。T4 處理的子蔓第1 節位結實花率最高,T1 處理次之,T3 處理最低,僅為66.67%。
由此可以看出,經過不同濃度多胺處理后,T1 和T4 處理的花期最早,子蔓第1 節位結實花率較高;CK 與T3 處理花期相同,子蔓第1 節位結實花率較低;T2 處理花期雖然最晚,但是子蔓第1 節位結實花率極顯著高于CK 和T3 處理。
2.3 外源多胺對薄皮甜瓜葉片中內源激素含量的影響
2.3.1 對GA3含量的影響 從圖1-a 可以看出,子葉展開未進行多胺處理時,葉片中GA3含量較低,進入花芽分化期后迅速升高,各處理在苗齡15~20 d 時均達到峰值,其中T4 處理葉片GA3含量在苗齡15 d 時(即雄蕊分化期)達到峰值,為195.1 μg·g-1(FW),顯著高于其他處理,但此后下降;其他處理在苗齡20 d 時達到峰值,CK 和T1 處理GA3含量低于其他處理。說明外源噴施多胺后,GA3含量產生變化,其中噴施1×10-4mol·L-1的Spd(即T4 處理)后,GA3含量明顯高于其他處理,較早達到峰值,同時T4 處理花芽分化速度快于其他處理。因此,外源多胺通過影響GA3含量影響了花芽分化,外源噴施多胺后GA3含量增高,花芽分化加快。
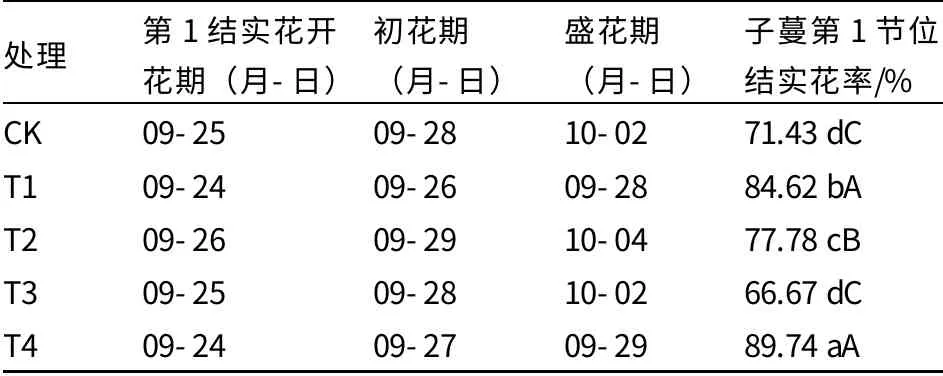
表2 外源多胺對薄皮甜瓜結實花花期和子蔓第1 節位結實花率的影響
2.3.2 對IAA 含量的影響 由圖1-b 可知,子葉展開時葉片中IAA 含量處于較高水平,為8.817 μg·g-1(FW);在苗齡5 d 時,各處理均進入花芽分化初期,葉片中IAA 含量急劇降低;苗齡10 d 時,各處理IAA 含量均有所上升,升高幅度不同,T1 和T4 處理顯著高于其他處理,此時正值第1 片真葉展開,植株生長迅速,石蠟切片顯示T1 和T4 處理正進入花冠裂片分化期,快于其他處理;到苗齡20 d 時,各處理IAA 含量均有所下降。因此,外源噴施多胺后對IAA 含量變化影響不大,IAA 含量變化可能與植株生長速度有關。
2.3.3 對ZT 含量的影響 由圖1-c 可知,葉片中ZT 含量在整個花芽分化期呈下降的趨勢,子葉展開時ZT 含量處于較高水平,為26.178 μg·g-1(FW),苗齡5 d 時進入花芽分化初期,各處理ZT 含量變化幅度較小,苗齡10~15 d 時,各處理ZT 含量均有所下降,其中T1 和T4 處理下降幅度較大,在苗齡15 d 時T1 和T4 處理ZT 含量到達最低值,苗齡20 d 時ZT 含量有所上升,而其他3 個處理均在苗齡20 d 時到達最低值,這可能與多胺處理后花芽分化速度和進程有關。T1 和T4 處理較其他3 個處理提前進入雄蕊分化期。因此,外源多胺處理影響葉片中ZT 含量,外源噴施1×10-3mol·L-1的Put(T1)和1×10-4mol·L-1的Spd(T4)使葉片中ZT 含量降低,花芽分化速度快于其他處理。
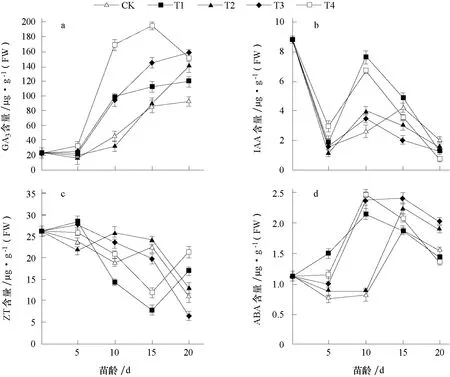
圖1 外源多胺對薄皮甜瓜葉片中內源激素含量的影響
2.3.4 對ABA 含量的影響 由圖1-d 可知,各處理葉片中ABA 含量變化幅度較小,子葉展開時ABA 含量處于相對較低水平,苗齡5~15 d 時各處理ABA 含量均有所上升,其中T1 和T4處理在苗齡10 d 時到達峰值,此后下降;而其他3 個處理均在苗齡15 d 時到達峰值,此后下降。因此葉片中ABA 含量變化與外源噴施多胺后花芽分化進程有關,T1 和T4 處理花芽分化速度較快。
2.4 外源多胺對薄皮甜瓜花芽中內源多胺含量的影響
由圖2-a 可知,子葉展開當天,即花芽分化的生理分化期時,花芽中內源Put 含量處于相對較高水平,為736.324 nmol·g-1(FW),說明花芽分化開始前需要積累大量的內源Put。此后花芽分化開始,各處理內源Put 含量迅速下降,在花芽分化初期(苗齡5 d 和10 d)各處理的內源Put 含量下降到最低值,其中CK 在苗齡10 d 時內源Put 含量僅為14.087 nmol·g-1(FW),苗齡10~15 d 時各處理內源Put 含量均呈上升趨勢,苗齡15~20 d 期間CK、T1 和T3 處理內源Put含量均保持上升趨勢,而T2 和T4 處理有下降趨勢。這可能與多胺處理后花芽分化進程不同有關,結合石蠟切片的花芽分化進程發現,雄蕊分化期時內源多胺的含量普遍較低,花芽分化后進入花器官分化期時內源多胺的含量相對較高。
圖2-b 中,在花芽分化的生理分化期時,內源Spd 含量處于較低水平,花芽分化初期各處理內源Spd 含量變化幅度較小,進入花芽形態分化期后內源Spd 含量有不同幅度上升,苗齡10 d時T1 和T4 處理內源Spd 含量達到峰值,而其他3 個處理在苗齡15 d 時才達到峰值,這可能與外源噴施多胺后花芽分化速度有關,T1 和T4 處理花芽分化速度快于其他處理,但在兩性分化期時T1 和T4 處理內源Spd 含量均有所下降,苗齡20 d 時T1 和T4 處理的內源Spd 含量高于其他處理,這可能與花芽分化進程有關,此時T1 和T4 處理完成花芽分化,已進入花器官分化期。因此,外源噴施多胺后影響內源Spd 含量變化,T1 和T4 處理的內源Spd 含量較早達到高峰。
由圖2-c 可知,花芽中內源Spm 含量變化呈下降趨勢,在花芽分化的生理分化期時內源Spm 含量處于較高水平,進入花芽形態分化期后內源Spm 含量有下降趨勢,苗齡5~15 d 時內源Spm 含量變化不規律,但T2 處理的內源Spm 含量高于其他處理。外源多胺對內源Spm 含量影響不大。
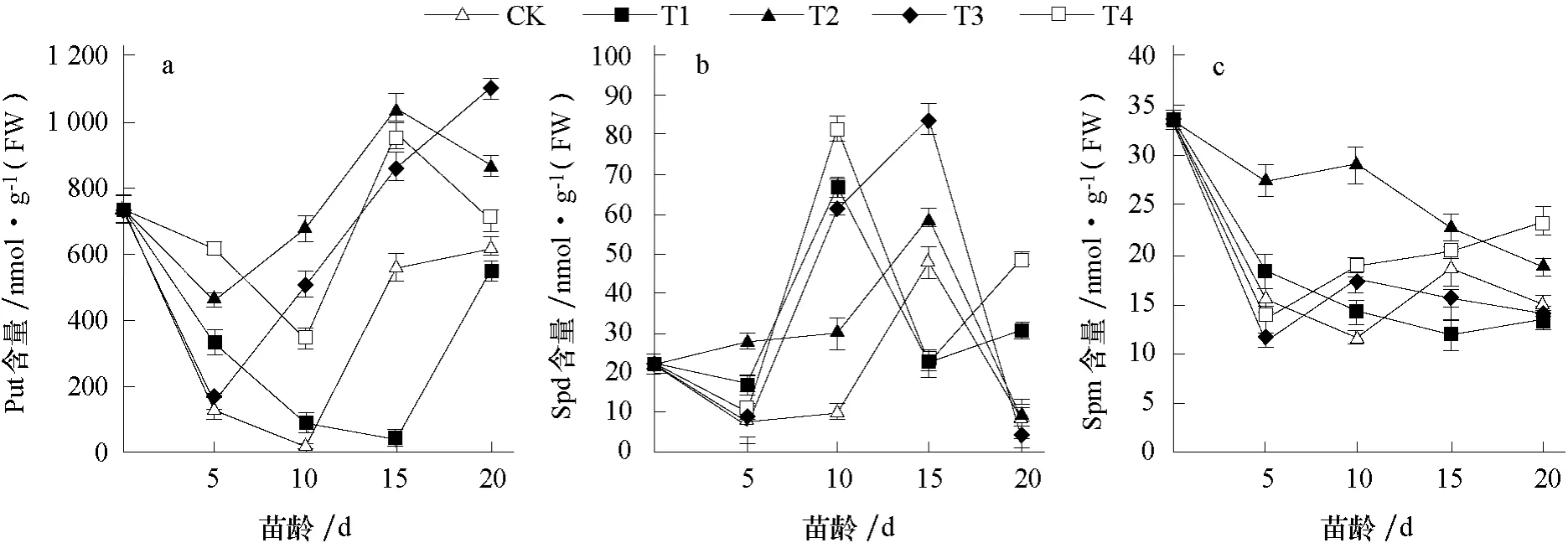
圖2 外源多胺對薄皮甜瓜花芽中內源多胺含量的影響
葉片中3 種內源多胺的變化趨勢與花芽中的大致相同,但花芽中3 種內源多胺的含量遠高于葉片中,其中花芽中內源Put 含量約為幼葉中的100 倍,葉片中的多胺可能大量運往花芽中以供給花芽分化。并且,外源噴施不同多胺后,內源多胺含量在大部分時期高于對照,可能是噴施外源多胺增加了內源多胺的含量。
3 結論與討論
3.1 外源多胺對薄皮甜瓜結實花花期、子蔓第1 節位結實花率和花芽分化進程的影響
不同濃度、不同種類的外源多胺對薄皮甜瓜花芽分化影響不同。外源噴施Put 和Spd 處理均能顯著增加核桃的結實花數量,提高雌雄花比例(徐繼忠 等,2004)。本試驗結果表明,外源噴施高濃度Put(1×10-3mol·L-1)和低濃度Spd(1×10-4mol·L-1)有利于提高甜瓜子蔓第1 節位結實花率,促進花芽分化,使結實花花期提前。
3.2 外源多胺對薄皮甜瓜葉片中內源激素含量的影響
植物激素與花芽分化關系密切。GA3與第1 結實花的發育有關(汪俏梅和曾廣文,1997);IAA 可能是黃瓜性別發育的關鍵性激素(陳學好 等,2002),外源噴施Spd 和Spm 可提高內源IAA 含量(桂仁意 等,2003);ZT 有助于甜瓜的性器官分化及發育(李計紅,2006);不同內源ABA 含量在花芽分化不同時期對蘋果梨的影響不同(李秉真 等,2000);在花芽分化期莖尖IAA、GA 含量保持較低的水平和適宜的PAs 含量有利于花芽分化(王廷芹和楊暹,2008)。通過本試驗可得出,外源噴施多胺使葉片中GA3含量增高,花芽分化加快。噴施高濃度Put(1×10-3mol·L-1)和低濃度Spd(1×10-4mol·L-1)后,葉片中IAA 含量高于其他處理,ZT 含量降低,ABA 含量提前達到峰值,這與兩處理的花芽分化速度快于其他處理有關。
3.3 外源多胺對薄皮甜瓜花芽中內源多胺含量的影響
外源噴施一定濃度的Put 和Spd 后,內源多胺的含量也隨之受到影響,從而影響花的性別分化和花發育(汪俏梅和曾廣文,1997;徐繼忠 等,2004)。前人對荔枝花芽分化研究時發現花芽和葉片的Spd 和Spm 比值在1~3 范圍,說明成熟組織也合成多胺,而且葉片的數量遠遠超過頂芽或花芽數量,它們合成的多胺總量比花芽多(肖華山 等,2006),因此,葉片多胺的合成對花芽的分化起著至關重要的作用。本試驗中,外源噴施多胺后,花芽中內源多胺含量在大部分時期高于對照,說明外源多胺施用促進了內源多胺的合成。
外源多胺可能通過影響內源激素含量進而調控基因表達影響花芽分化,或通過內源多胺含量進而影響激素平衡而調控基因表達影響花芽分化,或內源多胺含量升高直接調控基因表達影響花芽分化,這些還需要進一步試驗予以確定。
艾育芳,陳觀水,周以飛,潘大仁,寧文君.2011.油菜早熟突變體成花過程中葉片多胺代謝研究.中國農學通報,27(12):101-105.
陳學好,曾廣文,曹碚生.2002.黃瓜花性別分化和內源激素的關系.植物生理學通訊,38(4):317-320.
桂仁意,曹福亮,沈惠娟,謝寅峰.2003.多胺代謝對石竹試管苗成花中內源激素含量的影響.南京林業大學學報:自然科學版,27(4):27-30.
郝建軍,劉延吉.2001.植物生理學實驗技術.沈陽:遼寧科學技術出版社.
李秉真,孫慶林,張建華,馬青枝.2000.蘋果梨花芽分化期內源激素含量的變化(簡報).植物生理學通訊,36(1):27-29.
李計紅.2006.甜瓜性別分化的生理生化特性研究〔碩士論文〕.蘭州:甘肅農業大學.
汪俏梅,曾廣文.1997.苦瓜性別分化的激素調控.浙江農業大學學報,23(5):551-556.
王廷芹,楊暹.2008.外源赤霉素對青花菜莖尖內源激素含量的影響.中國蔬菜,(7):22-25.
肖華山,呂柳新,陳志彤.2006.荔枝花芽分化過程中多胺、核酸和蛋白質的動態.應用與環境生物學報,12(5):640-642.
徐繼忠,陳江海,李曉東,張志華,王艷輝.2004.外源多胺對核桃雌雄花芽分化及葉片內源多胺含量的影響.園藝學報,31(4):437-440.
張多嬌,齊紅巖,陳璐璐.2009.嫁接對薄皮甜瓜花芽分化和花發育的影響.中國蔬菜,(6):25-30.
張新生,陳湖,于麗辰,傅友.2005.多胺與板栗花性別分化的關系.河北農業科學,4(9):5-9.
張建農,李計紅.2007.甜瓜不同性別花蕾發育中激素和多胺的變化.園藝學報,34(5):1195-1200.
Altamura M M,Tomassi M.1998.Auxin,photoperiod and putrescine affect flower neoformation in normal androlB-transformed tobacco thin cell laye.Plant Physiology and Biochemistry,36(6):441-448.
de Dios P,Matilla A J,Gallardo M.2006.Flower fertilization and fruit development prompt changes in free polyamines and ethylene in damson plum(Prunus insiticaL.).J Plant Physiol,16:386-397.
Seo S G,Kang S W,Shim I S,Kim W,Fujihara S.2009.Effects of various chemical agents and early ethylene production on floral senescence ofHibiscus syriacusL.Plant Growth Regul,57:251-258.
