CCH1或MID1基因缺失對白念珠菌藥物耐受性及致病性的影響
2012-02-10 01:20:52王慧蘆廣慶楊寶鵬王帆喻其林徐寧程欣欣邢來君李明春
生物工程學報 2012年6期
關鍵詞:小鼠
王慧,蘆廣慶,楊寶鵬,王帆,喻其林,徐寧,程欣欣,邢來君,李明春
1 南開大學微生物學系 分子微生物學與技術教育部重點實驗室,天津 300071
2 農業部環境保護科研監測所,天津 300191
白念珠菌Candida albicans是醫院獲得性真菌病的主要感染源之一,也是深部真菌感染的首要病原菌,引起的致死率可達38%~49%[1-2]。臨床上,在白念珠菌感染的治療過程中,凸顯的嚴重問題就是菌株對各類藥物產生的抗藥性,尤其是對唑類藥物[3]。在光滑念珠菌Candida glabrata和釀酒酵母中,對唑類藥物的耐受性都是受到Ca2+信號調控的[4-5],當暴露于抗真菌藥物下時,細胞膜的穩定性受到干擾,導致胞內Ca2+濃度上升[6]。
鈣細胞存活 (Calcium cell survival,CCS)[7]途徑是 Bonilla等首次在釀酒酵母中發現的一個鈣信號轉導途徑,通過 Cch1-Mid1鈣通道引發Ca2+內流,并借助鈣信號的傳遞,激活鈣調神經磷酸酶 (Calcineurin,CaN) 及其下游轉錄因子Crz1p來介導細胞參與多種外界壓力下的存活。通常情況下,Cch1和Mid1組成蛋白復合體[8],Cch1是核心,作為復合體的催化亞基,而Mid1作為調節亞基輔助Cch1完成鈣通道復合體的功能,二者缺一不可。在釀酒酵母中,內質網壓力能夠通過 Mpk1途徑激活 Cch1-Mid1鈣離子通道,引起胞內 Ca2+濃度的上升,變化的 Ca2+與CaM結合,進而激活CaN[9]。因此,Cch1和Mid1對CaN的激活具有調控作用,之后借助胞內Ca2+的轉運體,降低胞內的Ca2+水平,同時也調控著這些胞內鈣泵的活性。
CCS途徑中的CaN參與抗真菌藥物的耐受性[10],其催化亞基Cna1或調節亞基Cnb1任何一個的缺失都會對抗真菌藥物表現出強烈的敏感性,同時對白念珠菌菌株毒力也具有減弱的作用,有研究表明,對唑類藥物例如氟康唑FLUC呈現耐受性的菌株往往具有更強的致病性[11],可見白念珠菌的藥物耐受性與致病性之間有密切的關系。研究發現,作用于 CaN的免疫抑制劑環孢菌素 (Cyclosporin A,CsA) 或他克莫司(FK506) 與氟康唑 (FLUC) 的聯合使用具有殺滅真菌的作用,而FLUC的單獨使用僅僅具有抑制真菌的作用[12]。CCS途徑對于真菌在唑類藥物壓力下的存活發揮著重要作用,缺少 CCS途徑的酵母在僅僅能抑制細胞生長的唑類藥物濃度下就死亡了[13],CCS途徑中其他成員或是尚未鑒定出來的新成員也有可能參與到藥物耐受性中。是否 Cch1-Mid1也參與白念珠菌的藥物耐受性?我們將利用微量液基稀釋法和藥物平板敏感性試驗進行研究,并進一步通過建立小鼠系統感染模型的方法研究CCH1或MID1基因的缺失對菌株毒力的影響。
1 材料與方法
1.1 菌株、質粒和引物
實驗使用的菌株和質粒見表1,引物見表2。
1.2 主要試劑
CIAP堿性磷酸酶、各種限制性內切酶、Taq DNA聚合酶等購自寶生物大連有限責任公司;RPMI1640培養基粉末購自GBCO公司;5-氟乳清酸 (5-FOA) 購自北京鼎國生物技術有限責任公司;氟康唑 (FLUC)、伊曲康唑 (ITRA) 等唑類藥物購自Sigma公司;鹽酸維拉帕米 (Vpi)、鹽酸特比萘芬 (TER) 等藥物購自中國藥品生物制品檢定所;其余有機試劑和藥品均為國產分析純,所有引物均由北京奧科公司合成。
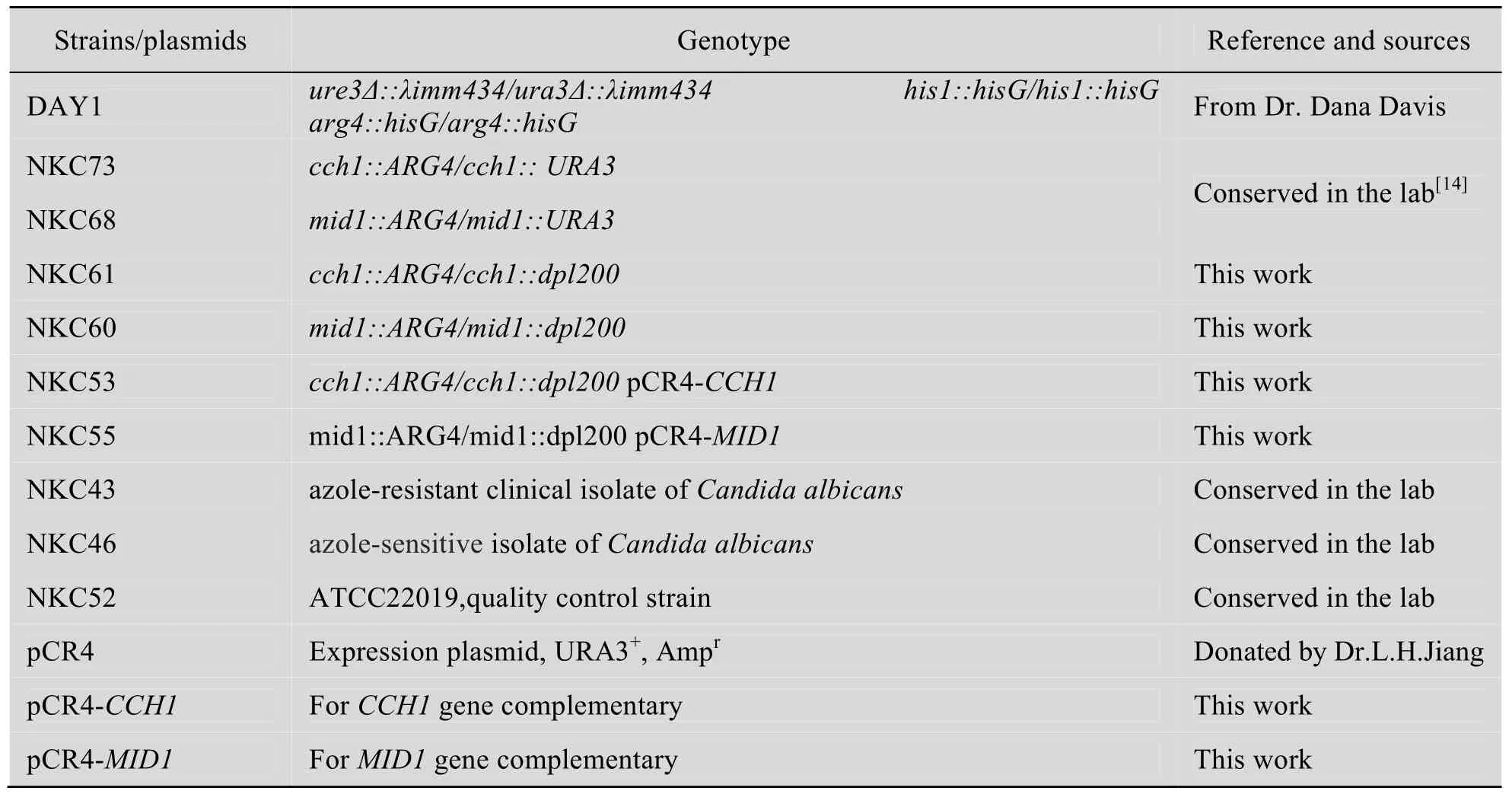
表1 本實驗使用的菌株和質粒Table 1 Strains and plasmids used in this study

表2 本實驗所用的引物Table 2 Primers used in this study
1.3 培養基
白念珠菌培養使用的所有培養基 (SC-ura除外) 均添加終濃度為80 mg/L的尿苷。YPD培養基:1%酵母浸出粉,2%蛋白胨,2%葡萄糖。SC培養基:0.67%無氨基酵母氮源,0.2%氨基酸混合物粉末,2%葡萄糖。5-氟乳清酸 (5-FOA)培養基 (1 L):6.7 g無氨基酵母碳源,20 g葡萄糖,0.8 g尿苷,2 g無氨基酸混合粉末,20 g瓊脂粉,1 g 5-氟乳清酸 (5-FOA)。RPMI 1640培養基:2.080 g RPMI 1640粉末,6.906 g苯磺酸 (MOPS),40 mL 10%葡萄糖溶液,加蒸餾水定容到200 mL,混勻后用1 mol/L NaOH溶液調pH至7.0,使用前進行抽濾除菌。
1.4 白念珠菌CCH1和MID1基因回補質粒的構建
設計一對帶有PstⅠ酶切位點的引物CCH1-5RC和CCH1-3RC,以NKC18基因組DNA為模板,擴增獲得一條8.7 kb的片段,其中包括1 kb的啟動子、CCH1基因的ORF區和終止子區。將該片段用PstⅠ進行酶切,純化后與同樣經過PstⅠ內切酶消化過,并經堿性磷酸酶CIAP去磷酸化處理的白念珠菌表達質粒pCR4進行連接,然后將連接產物轉化大腸桿菌,并在 LB+Amp的固體培養基中篩選轉化子,獲得正確的質粒pCR4-CCH1。pCR4-MID1質粒的獲得需要設計一對帶有 BamHⅠ和 KpnⅠ酶切位點的MID1-5RC和MID1-3RC引物,同樣以NKC18基因組DNA為模板,進行PCR擴增,獲得一條3.1 kb包含MID1基因1 kb啟動子、ORF區和終止子區的片段。將該片段用BamHⅠ和KpnⅠ進行雙酶切,純化后與同樣經過 BamHⅠ/KpnⅠ消化過的pCR4質粒進行連接,轉化大腸桿菌并獲得正確的質粒pCR4-MID1。
1.5 藥物平板敏感性試驗
挑取不同實驗菌株的單菌落,接種至 5 mL YPD液體培養基中,30 ℃過夜培養,再轉接至30 mL YPD液體培養基中使起始OD600約為0.1,30 ℃培養大約4~5 h到對數生長期,加入藥物氟康唑或伊曲康唑至各預計的終濃度,在37 ℃進行振蕩培養。分別在預先設定的時間點:0、5、10、20、30、60、90 min時取樣,每次取5 μL點在固體YPD平板上,點樣后,37 ℃培養24 h,觀察并拍照。
1.6 微量液基稀釋法
依照酵母菌敏感性測定 CLSI-M27-A2方案[15]的微量液基稀釋法對野生型菌株和其他各缺失菌株進行藥物敏感性測定。
1.7 小鼠感染模型的建立
進行毒力分析建模的小鼠為ICR雌性小鼠,鼠齡為6~8周。注射用的各菌株在SC培養基中過夜培養并轉接至新鮮培養基中再培養3~4 h,然后取適量菌液離心后懸浮于1 mL 0.9%生理鹽水中,用血球計數板計數。分別用生理鹽水將各株注射用菌株的菌液濃度調至 5×106cells/mL。用0.25 mL注射器和4號針頭注射小鼠尾靜脈,每只小鼠注射0.1 mL菌液,每種菌株注射10只小鼠。隨后每天對小鼠的死亡情況進行觀察和記錄,觀察周期為30 d。通過繪制小鼠的存活率曲線,利用SPSS軟件進行Kaplan-Meier Test統計學分析,來判定不同菌株的毒力變化情況。
2 結果與分析
2.1 白念珠菌CCH1和MID1基因回補缺失菌株的構建
質粒pCR4含有的選擇標記為URA3,缺失菌株cch1Δ/Δ (cch1::CCH1/cch1:: URA3-dpl200)和 mid1Δ/Δ (mid1::MID1/mid1:: URA3-dpl200)中的URA3標記必須被消除后,才能使用構建的pCR4-CCH1和pCR4-MID1質粒用于相應基因的回補。分別將cch1Δ/Δ和mid1Δ/Δ缺失菌株涂布于含有5-FOA和尿苷的SC培養基中,用于標記的彈出。3 d后挑取轉化子,提取基因組并以CCH1/MID1-5det和CCH1/MID1-3det為引物,檢測基因型。對于沒有去除URA3標記的菌株,PCR得到兩條帶,分別為ARG4和URA3標記帶[14];而對于去除URA3標記的菌株,ARG4標記帶保留,由于缺失菌株中的URA3-dpl200片段兩端含有同源序列,在 5-FOA選擇壓力下能夠將自身環化,從基因組中彈出一部分,使檢測帶變短。如圖 1所示,對于 URA3標記消除的cch1Δ/Δ菌株,PCR檢測正確的結果為 2.3 kb (ARG4) 和1.0 kb (泳道1,2);對于mid1Δ/Δ菌株,PCR檢測正確的結果為2.4 kb (ARG4) 和750 bp (泳道3,4),表明我們獲得了正確的URA3標記被消除的菌株 cch1::ARG4/cch1::dpl200 和mid1::ARG4/mid1:: dpl200,分別命名為 NKC61和NKC60。
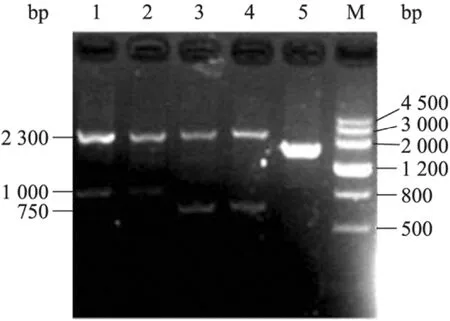
圖1 URA3標記消除的cch1Δ/Δ和mid1Δ/Δ菌株的鑒定Fig. 1 PCR analysis of URA3 dropping out in cch1Δ/Δ and mid1Δ/Δ strains. 1?4: genomic DNA from cch1Δ/Δ and mid1Δ/Δ strains were used as templates for PCR detection; 5: NKC18 was used as template with primers MID1-5detect and MID1-3detect (1.7 kb); M: DNA marker Ⅲ.
將pCR4和pCR4-CCH1分別轉化NKC61菌株,再將pCR4和pCR4-MID1分別轉化NKC60菌株,在SC-Ura選擇性培養基中篩選轉化子,分別以CCH1/MID-RC1 和CCH1/MID1-RC2為檢測引物進行轉化子的 PCR驗證,該引物從CCH1和MID1基因所敲除的那部分序列中設計選取,鑒定結果如圖2所示:對于cch1Δ/Δ+CCH1回補菌株,擴增得到 1.0 kb的片段;對于mid1Δ/Δ+MID1回補菌株,擴增得到824 bp的片段;而轉化空質粒的 cch1Δ/Δ+pCR4 和mid1Δ/Δ+pCR4菌株,PCR擴增沒有能夠得到目的條帶,依此鑒定并獲得了正確的回補缺失菌株cch1Δ/Δ+CCH1和 mid1Δ/Δ+MID1,并將該菌株分別命名為NKC53和NKC55。
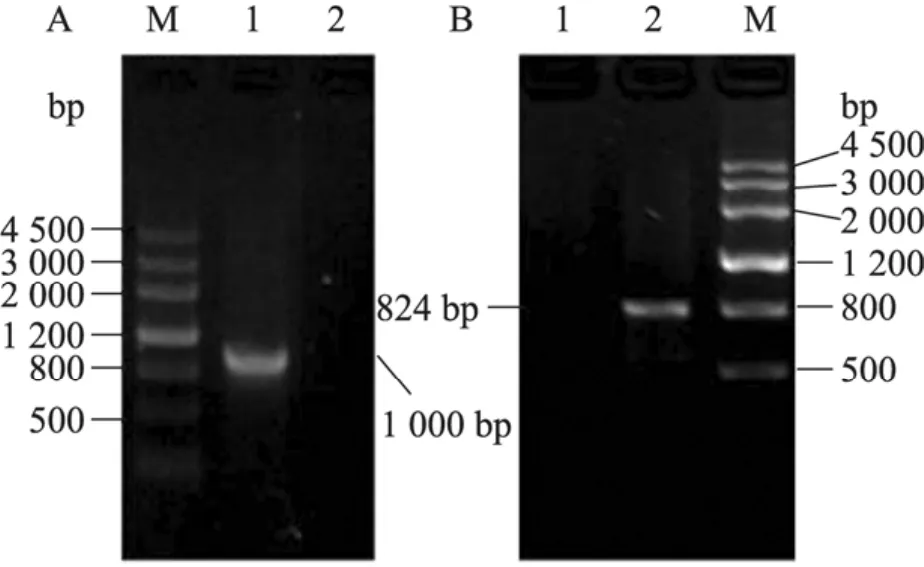
圖2 回補缺失菌株 cch1Δ/Δ+CCH1和 mid1Δ/Δ+ MID1的鑒定Fig. 2 PCR analysis of cch1Δ/Δ and mid1Δ/Δ recomplementation. M: markerⅢ. (A) 1: cch1Δ/Δ+ CCH1 (1.0 kb); 2: cch1Δ/Δ+pCR4. (B) 1: mid1Δ/Δ+ pCR4; 2: mid1Δ/Δ+MID1 (824 bp). Genomic DNA from all strains was used as templates with primers CCH1/MID-RC1 and CCH1/MID1-RC2.
2.2 CCH1或MID1基因的缺失對藥物敏感性的變化
為了更方便地對藥敏實驗結果進行比較分析,使用從臨床分離的耐藥型和敏感型的白念珠菌,作為實驗對照組。以ATCC22019作為質控菌株,如表3所示,質控菌株對FLUC的MIC值為2或4 mg/L,對ITRA的MIC值為0.064或0.128 mg/L,均在CLSI M27-A2規定的范圍內[16],證明操作無誤,測定結果準確可靠。用微量液基稀釋法測定菌株對藥物的敏感性時,通常用以下標準進行界限判定:以 FLUC對菌株的MIC≤8 mg/L為敏感,16~32 mg/L為劑量依賴性敏感,MIC≥64 mg/L為耐藥;以ITRA對菌株的MIC≤0.12 mg/L為敏感,0.25~0.5 mg/L為劑量依賴性敏感,≥1 mg/L為耐藥。表3的結果顯示:1) 對照組CA03997和CA90菌株的MIC測定結果均符合以上判定標準,分別為藥物耐受菌和敏感菌,保證了實驗體系沒有問題;2) 野生型菌株對FLUC和ITRA藥物的敏感性可以界定為劑量依賴性敏感;3) cch1Δ/Δ 和mid1Δ/Δ缺失菌株對FLUC和ITRA均表現出藥物敏感性,并接近于敏感菌CA90的MIC值;4) 回補缺失菌株cch1Δ/Δ+CCH1和mid1Δ/Δ+MID1對藥物的敏感性也在接近劑量依賴性敏感的范圍,接近于野生型菌株的水平,表明Cch1-Mid1參與了白念珠菌對唑類藥物耐受性的調控。
我們同時也利用固體平板藥物敏感性試驗,觀察并分析了上述不同菌株對藥物 FLUC敏感性的變化。將處于對數生長期的細胞轉移至添加有8 mg/L FLUC的YPD培養基中,在37 ℃振蕩培養,并且按圖3所示的不同時間點進行取樣,分別取樣5 μL,點種在YPD固體平板上。37 ℃靜置培養16 h后,觀察結果如圖3所示:cch1Δ/Δ和mid1Δ/Δ缺失菌株對FLUC表現出高度的敏感性,菌體生長狀況與敏感菌相似;CCH1和MID1基因的回補對這種敏感表型具有恢復作用,但不能恢復至與野生型菌株相似的敏感表型;以上結果再次表明CCH1和MID1基因的缺失與FLUC藥物敏感性有關。
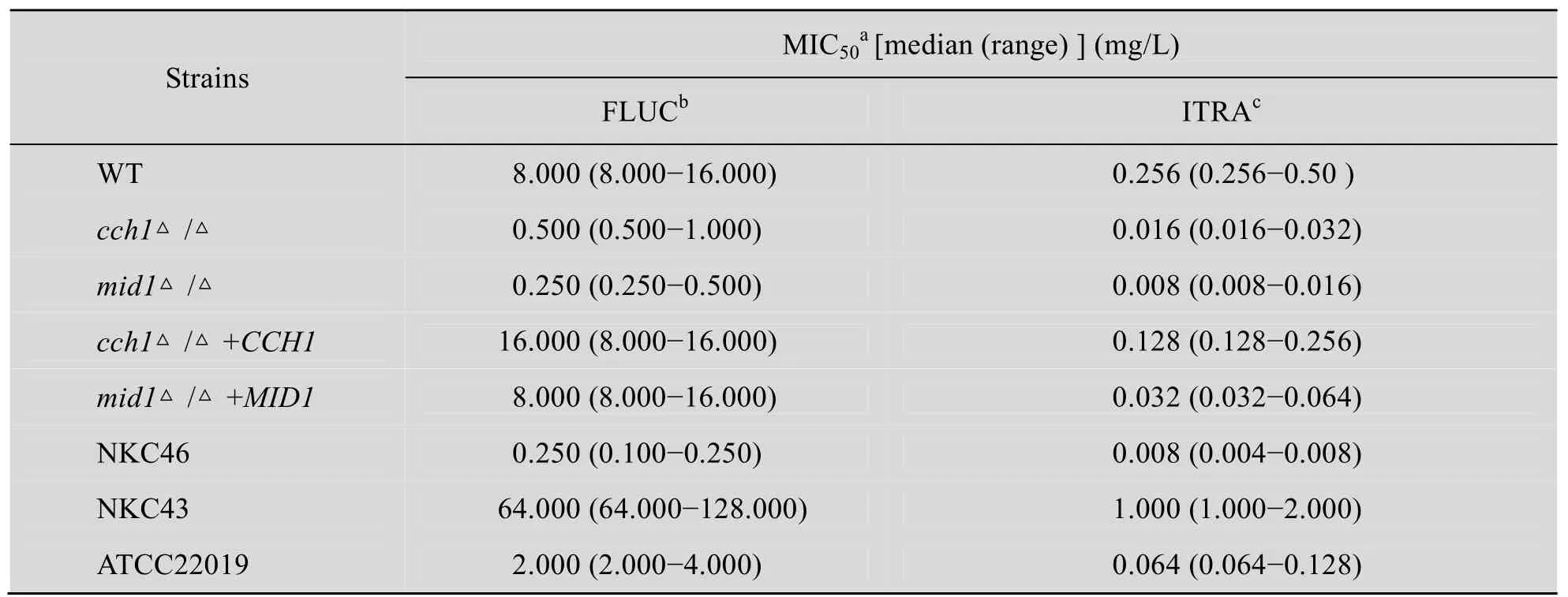
表3 各菌株對唑類藥物的敏感性Table 3 Susceptibility of C. albicans strains to azoles
2.3 CCH1和MID1基因參與藥物耐受的機制推測
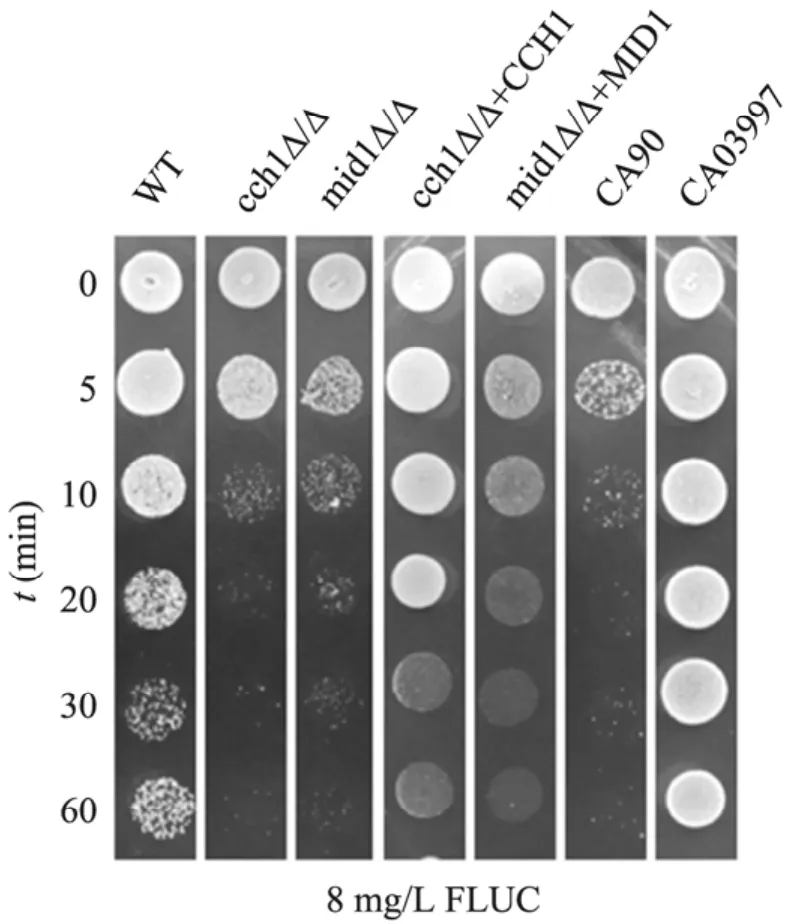
圖3 FLUC藥物平板敏感性試驗Fig. 3 Susceptibility assay to FLUC.
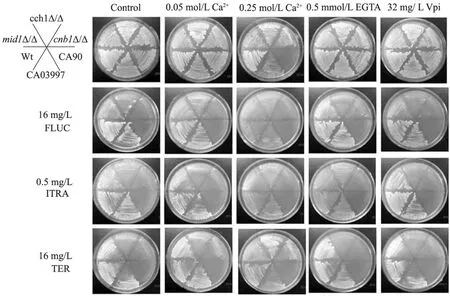
圖4 Ca2+對藥物抗真菌活性的調節作用Fig. 4 Calcium modulation of antifungal drug activity.
Cch1-Mid1鈣通道調控 Ca2+的內流,而CCH1和 MID1基因的缺失對藥物表現出敏感性,Ca2+的濃度調節很可能與藥物耐受性有關,所以我們在藥物FLUC、ITRA和TER作用的基礎上,分別添加0.05 mol/L或0.25 mol/L的Ca2+、0.5 mmol/L鈣螯合劑EGTA或32 mg/L鈣通道阻滯劑維拉帕米Vpi,利用劃線培養的方法,對以上不同菌株的生長情況進行了比較,來研究CCH1和MID1基因參與藥物耐受的機制。培養基放置于30 ℃培養,3 d后觀察結果,如圖4所示:1) 在對照組所在列,除野生型菌株和耐藥菌CA03997外,其他各缺失菌株和敏感菌CA90對唑類藥都表現出不同程度的敏感性;2) 低濃度0.05 mol/L Ca2+的添加抑制了唑類藥物對白念珠菌的抗真菌活性,與對照組相比,所有菌株的生長狀況有所好轉,表明少量 Ca2+的存在幫助細胞獲得了一定程度的對唑類藥物的耐受性;3) 高濃度0.25 mol/L Ca2+、EGTA或Vpi的添加,與對照組相比,都增強了唑類藥物對各菌株的抑制作用,使菌株的生長狀況變得更差了;4) 然而,低濃度和高濃度Ca2+的添加都抑制了丙烯酰胺類藥物TER的作用活性,但EGTA或Vpi的添加卻同樣也對 TER活性有增強作用,菌株表現出對藥物更加敏感。綜合以上結果表明:不同的 Ca2+濃度能夠通過改善或抑制藥物的抗真菌活性,來間接調節白念珠菌對藥物的耐受性,雖然對唑類藥物和丙烯酰胺類藥物耐受的調控方式不同,但可以確定的是CCH1和MID1基因參與藥物耐受性的調控,也是通過對Ca2+濃度變化的間接調節來實現的。
2.4 CCH1或MID1基因的缺失對白念珠菌毒力的影響
白念珠菌的藥物耐受性和致病性之間有密切關系,所以我們通過建立小鼠系統感染模型的方法來進一步分析CCH1和MID1基因在影響菌株毒力方面的作用。在實驗中,共飼養 50只小鼠,將這些小鼠隨機平均分成1~5組,各組小鼠通過尾靜脈分別注射野生型菌株、cch1Δ/Δ+ CCH1和mid1Δ/Δ+MID1回補菌株以及cch1Δ/Δ和mid1Δ/Δ缺失菌株,每種菌株注射10只小鼠,然后每天觀察并記錄小鼠的存活情況,觀察周期為30 d。利用Kaplan-Meier Test進行統計學分析,判定不同組小鼠之間的生存差異,結果表明:注射野生型菌株、cch1Δ/Δ+CCH1和 mid1Δ/Δ+ MID1小鼠的平均存活時間分別為 12.2、14.6、15 d,統計學分析結果顯示,在野生型菌株和回補菌株之間沒有統計學差異 (P=0.306或0.233),然而注射cch1Δ/Δ和mid1Δ/Δ缺失菌株小鼠的平均存活時間為22.9 d和21.3 d,與野生型菌株之間存在差異,并且差異在極顯著水平 (P<0.01) (圖5)。實驗結果表明,CCH1和MID1基因的缺失導致菌株的毒力大大降低。
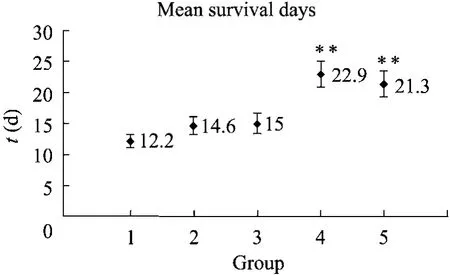
圖5 感染不同菌株的小鼠存活中位數分析Fig .5 Virulence assay of C. albicans strains.
3 討論
CCS途徑在介導酵母細胞存活的過程中發揮著非常廣泛的作用,在這個途徑中,鈣調神經磷酸酶的功能在許多真核細胞中已被證實,并且有證據表明Cch1-Mid1鈣通道能夠通過調控Ca2+的內流,對鈣調神經磷酸酶產生激活作用[7,13]。最初的 Ca2+波動引起一系列相關應答來幫助細胞在壓力環境下存活,說明Cch1p和Mid1p在保證細胞存活方面發揮著最為基礎的作用,因此,在我們的工作中,重點對白念珠菌 CCS途徑中CCH1和MID1基因缺失所引起的藥物耐受性和菌株毒力的變化進行研究。
CCH1和 MID1基因參與藥物耐受性的調控,可以理解為通過調節Ca2+的作用,參與介導細胞在藥物壓力下的存活。這在我們的實驗中也得到證實,CCH1和MID1基因的缺失在藥物作用下表現的更為敏感,使用Ca2+的螯合劑EGTA或鈣通道阻滯劑 Vpi聯合抗真菌藥物一起作用時,能夠改善藥物作用。所以,以此確定 CCS途徑通過對 Ca2+的調節,間接參與了藥物耐受性。小鼠感染模型實驗證明cch1Δ/Δ和mid1Δ/Δ菌株的毒力明顯比野生型菌株弱,這與許多基因缺失突變對菌株毒力的影響非常相似。白念珠菌毒力的形成也是多種因素累積的效果,比如蛋白酶的分泌[11],細胞表面黏附素的合成[17]等。當細胞在接受外界持續和難以預知的生物或物理因子干擾的時侯,包括溫度、環境pH、離子毒力、氧化脅迫壓力和藥物作用等,白念珠菌在借助適當的途徑來感知和應答這些環境信號的同時,就會引起細胞自身的生理學、形態以及粘附特性的改變,從而提高細胞對環境變化的適應性,因此表現為菌株毒力發生變化。在我們的實驗中同時也發現cch1Δ/Δ和mid1Δ/Δ菌株并不是完全毒力喪失,只是毒力減弱了,但仍然能夠對小鼠產生致死性的感染。這表明小鼠體內的這種環境可能誘導了其他一種或幾種途徑促進了 cch1Δ/Δ和mid1Δ/Δ菌株的菌絲形成,或者與鈣調神經磷酸酶的作用有關。總之,CCH1和MID1基因在參與白念珠菌藥物耐受性和致病性調控方面發揮著重要作用。白念珠菌的致病性與藥物耐受性之間有密切的關系。分泌型天冬氨酸蛋白酶 (Sap) 目前被認為是白念珠菌主要的毒力因子之一[18],與白念珠菌的黏附力和組織侵襲有關。研究發現對藥物呈現耐受性的菌株比藥物敏感株具有更大的毒性,而且耐藥株在FLUC濃度逐漸增加的環境下生長會導致劑量依賴性的體外分泌型天冬氨酸蛋白酶產量增加[19],以及耐藥基因 MDR1的過度表達。Fekete-Forgacs等將FLUC敏感型的野生型白念珠菌誘導為耐FLUC的實驗變異株,發現變異株的Sap分泌明顯強于自然分離菌株[20],在臨床白念珠菌耐藥和致病性的研究中,更多的是用蛋白酶活力作為反映耐藥菌株毒力和致病性強弱的直接指標[21]。所以,在我們的實驗中,是否在那些對藥物呈現敏感性的 cch1Δ/Δ和
mid1Δ/Δ菌株中,分泌型天冬氨酸蛋白酶的產量也會表現為降低,還有待進一步研究證實。
致謝:本實驗中的部分菌株和質粒由明尼蘇達大學Dr. Dana Davis教授和江南大學蔣伶活教授惠贈,在此表示衷心感謝!
[1] Koh AY, K?hler JR, Coggshall KT, et al. Mucosal damage and neutropenia are required for Candida albicans dissemination. PLoS Pathog, 2008, 4(2): e35.
[2] Ruhnke M, Maschmeyer G. Management of mycoses in patients with hematologic disease and cancer——review of the literature. Eur J Med Res, 2002, 7(5): 227?235.
[3] Andes D, Forrest A, Lepak A, et al. Impact of antimicrobial dosing regimen on evolution of drug resistance in vivo: fluconazole and Candida albicans. Antimicrob Agents Chemother, 2006, 50(7): 2374?2383.
[4] Edlind T, Smith L, Henry K, et al. Antifungal activity in Saccharomyces cerevisiae is modulated by calcium signalling. Mol Microbiol, 2002, 46(1): 257?268.
[5] Kaur R, Casta?o I, Cormack BP. Functional genomic analysis of fluconazole susceptibility in the pathogenic yeast Candida glabrata: roles of calcium signaling and mitochondria. Antimicrob Agents Chemother, 2004, 48(5): 1600?1613.
[6] Patterson RL, van Rossum DB, Gill DL. Storeoperated Ca2+entry: evidence for a secretion-like coupling model. Cell, 1999, 98(4): 487?499.
[7] Bonilla M, Cunningham KW. Mitogen-activated protein kinase stimulation of Ca2+signaling is required for survival of endoplasmic reticulum stress in yeast. Mol Biol Cell, 2003, 14(10): 4296?4305.
[8] Peiter E, Fischer M, Sidaway K, et al. The Saccharomyces cerevisiae Ca2+channel Cch1pMid1p is essential for tolerance to cold stress and iron toxicity. FEBS Lett, 2005, 579(25): 5697?5703.
[9] Aramburu J, Heitman J, Crabtree GR. Calcineurin: a central controller of signalling in eukaryotes. EMBO Rep, 2004, 5(4): 343?348.
[10] Sanglard D, Ischer F, Marchetti O, et al. Calcineurin A of Candida albicans: involvement in antifungal tolerance, cell morphogenesis and virulence. Mol Microbiol, 2003, 48(4): 959?976. [11] Wu T, Wright K, Hurst SF, et al. Enhanced extracellular production of aspartyl proteinase, a virulence factor, by Candida albicans isolates following growth in subinhibitory concentrations of fluconazole. Antimicrob Agents Chemother, 2000, 44(5): 1200?1208.
[12] Li Y, Sun SJ, Guo QJ, et al. In vitro interaction between azoles and cyclosporin A against clinical isolates of Candida albicans determined by the chequerboard method and time-kill curves. J Antimicrob Chemother, 2008, 61(3): 577?585.
[13] Bonilla M, Nastase KK, Cunningham KW. Essential role of calcineurin in response to endoplasmic reticulum stress. EMBO J, 2002, 21(10): 2343?2353.
[14] Wang H, Xu N, Xing LJ, et al. Effect of CCH1 and MID1 in calcium influx under alkaline pH and its regulation by Crz1p transcription factor in Candida albicans. Chin J Biotech, 2011, 27(6): 917?925.王慧, 徐寧, 邢來君, 等. 堿性pH條件下白念珠菌鈣離子通道CCH1和MID1基因對鈣內流的影響及Crz1p轉錄因子對其的調控作用. 生物工程學報, 2011, 27(6): 917?925.
[15] CLSI/NCCLS, Reference method for broth dilution antifungal susceptibility testing of yeasts. Approved standard M27-A2. National Committee for Clinical Laboratory Standards, 2002.
[16] Pfaller MA, Messer SA, Bolmstr?m A, et al. Multisite reproducibility of the Etest MIC method for antifungal susceptibility testing of yeast isolates. J Clin Microbiol, 1996, 34(7): 1691?1693.
[17] Krasowska A, Murzyn A, Dyjankiewicz A, et al. The antagonistic effect of Saccharomyces boulardii on Candida albicans filamentation, adhesion and biofilm formation. FEMS Yeast Res, 2009, 9(8): 1312?1321.
[18] Schaller M, Januschke E, Schackert C, et al. Different isoforms of secreted aspartyl proteinases (Sap) are expressed by Candida albicans during oral and cutaneous candidosis in vivo. J Med Microbiol, 2001, 50(8): 743?747.
[19] Costa CR, Jesuíno RSA, de Aquino Lemos J, et al. Effects of antifungal agents in sap activity of Candida albicans isolates. Mycopathologia, 2010, 169(2): 91?98.
[20] Fekete-Forgács K, Gyüre L, Lenkey B. Changes of virulence factors accompanying the phenomenon of induced fluconazole resistance in Candida albicans. Mycoses, 2000, 43(7/8): 273?279.
[21] Jayatilake JAMS, Samaranayake YH, Cheung LK, et al. Quantitative evaluation of tissue invasion by wild type, hyphal and SAP mutants of Candida albicans, and non-albicans Candida species in reconstituted human oral epithelium. J Oral Pathol Med, 2006, 35(8): 484?491.
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34