miR-34s與大腸癌的發生
2012-01-26 07:59:50王一飛綜述梁徳森審校
重慶醫學 2012年18期
王一飛 綜述,梁徳森 審校
(哈爾濱醫科大學普外八科 150001)
miR-34s與大腸癌的發生
王一飛 綜述,梁徳森 審校
(哈爾濱醫科大學普外八科 150001)
微RNAs;細胞凋亡;細胞周期;大腸;腫瘤
隨著現代人類的生活環境和生活方式的改變,腫瘤的發生率也呈現出迅速增長的趨勢,其中消化道腫瘤是惡性腫瘤中發生率相對較高的疾病,大腸癌(colorectal cancer,CRC)是消化道腫瘤常見疾病之一,僅次于胃癌?食管癌?大腸癌是一個多基因?多階段?長期形成的復雜的病變過程,是大腸黏膜上皮起源的惡性腫瘤,雖然近年來新的診斷治療方法相繼問世,但大腸癌的病死率仍較高,目前大腸癌發生?發展的基因調控機制越來越受到人們的關注,其中MicroRNAs(miRNAs)作為一種新的內源性非編碼RNA與大腸癌的發生?發展密切相關,本文將就miR-34家族(miR-34s)與大腸癌的發生及其研究進展進行綜述?
1 miRNAs簡介
miRNAs是一種新的內源性非編碼 RNA(non-coding RNAs,ncRNAs),它廣泛存在于真核生物中,是生物體內重要的基因調節器[1-4],研究證明,miRNAs在細胞的分化?增殖?發育?新陳代謝及凋亡過程中都起著非常重要的作用,它與人類腫瘤細胞的生成?增殖?發育過程關系非常密切,在miRNAs編碼的基因中既有癌基因又有抑癌基因[1,3-4],所以它的不恰當的表達也就成了腫瘤發生的一種特征性標志[3,5-6],實驗證明,不同家族成員的miRNAs的表達與對應類型癌的發生有著較為明確的關系[7]?它們在腫瘤的發生?發展過程中扮演著重要的角色?
2 miRNAs與腫瘤發生
目前發現,在人類中有近1 000多種miRNAs,30%的人類基因都不同程度的受其控制[8],隨著研究的深入,miRNAs調控的靶基因越來越多地被揭示,人們發現一種miRNAs在不同腫瘤的表達作用情況不同甚至相反,因此,單純以抑癌與促癌把 miRNAs分成兩類顯然是不準確的[9-10]?3種機制:(1)miRNAs定位的染色體異常[11];(2)表觀遺傳學改變[12];(3)miRNAs加工相關的基因及其蛋白的異常變化[13]?通過以上3種機制不難發現miRNAs與腫瘤的發生在基因水平上有著極為密切的關系,所以說miRNAs與腫瘤細胞的發生?發展?抑制等活動有著不容置疑的聯系?
3 miR-34家族介紹及激活
3.1 miR-34家族介紹 miR-34是最早從線蟲中被發現的一段保守miRNAs,而后在脊椎動物中發現3個成員:miR-34a?miR-34b以及miR-34c?不同物種中miR-34基因的同源性很高,其中成熟序列的同源性為68%,前體序列的同源性為38.89%[14]?人源 miR-34a的編碼序列位于轉錄前體的第2外顯子內[15-16],miR-34b和 miR-34c的編碼序列分別位于第1內含子和第2外顯子中[16]?染色質免疫沉淀反應和位點突變實驗表明上述序列即為P53蛋白結合位點,并且該結合序列前存在一個CpG島(CpG island)[17]?在某些腫瘤細胞中,正是由于該CpG島被甲基化阻礙了P53對miR-34的激活,從而使p53通過miR-34對細胞增殖的調控出現障礙,進而加速了腫瘤細胞的生長[18]?這一現象已在多種癌細胞中被發現,如肺癌?乳腺癌?前列腺癌,直腸癌和黑色素瘤等[19-20]?
3.2 miR-34的激活 miR-34激活機制在近年來也得到了廣泛的研究,通過RT-PCR方法 He等[16]對鼠胚胎成纖維細胞的145種miRNAs進行了檢測,發現miR-34的表達與p53基因有關?Tarasov等[15]發現miR-34a在p53激活后增加了300余倍,又進一步研究發現在 MCF-7細胞系中,加入能夠激活p53基因的依托泊苷可誘發miR-34a原初轉錄本(pri-mir-34a)的表達?隨后,人們通過核蛋白免疫共沉淀法發現miR-34存在著與p21類似的p53結合區域[16,18,21],并且發現在 Has-mir-34a的外顯子1內及Has-mir-34b/c的啟動區域內存在與p53經典結合區域相吻合的回文序列[15-16]?這些研究為p53直接作用并激活miR-34提供了有力的證據?截止到目前經確認的miR-34s作用靶蛋白及細胞因子有 CDK4?CDK6?CyclinE2?E2F3?E2F5?Met?Bcl-2及原癌基因c-myc等[22-23],miR-34通過被p53激活,抑制這些靶蛋白表達從而抑制腫瘤細胞生長?
4 miR-34a與大腸癌的發生
目前已發現多種miRNAs在大腸癌組織及大腸癌細胞系中異常表達,其中一部分在癌細胞中較正常細胞表達明顯下降,如 miR-143?miR-145?let-7?miR-34a等,一部分表達則升高,如 miR-31?miR-21等?近年來 miR-34作為一種比較新穎和熱門的大腸癌抑癌基因得到了人們的廣泛研究?
4.1 miR-34a促進大腸癌細胞凋亡
4.1.1 通過降低Bcl-2含量促進大腸癌細胞凋亡 Bcl-2蛋白通過抗細胞凋亡作用,在腫瘤的發生?進展以及耐藥性等方面發揮重要作用,作為一種癌基因的產物Bcl-2在大腸癌細胞中有著過度的表 達[24-25],Cole等[26]發 現 miR-34a的 過 度 表 達 能夠導致 Bcl-2mRNA的減少?同時 Bommer等[18]通過向Bcl-2的3′非編碼區熒光素酶報告質粒轉染細胞系中加入miR-34模擬子后發現miR-34a能夠不同程度地抑制熒光素酶報告基因的表達,表明miR-34a能直接作用于Bcl-2的3′非編碼區抑制Bcl-2蛋白的合成?因此,不難推導出miR-34可通過對Bcl-2進行調控從而影響大腸癌細胞凋亡?
4.1.2 通過抑制SIRT1促進大腸癌細胞凋亡 SIRT1是一類NAD依賴的脫乙酰化酶,在腫瘤的形成過程中發揮重要作用[27],SIRT1能夠通過脫乙酰化作用使p53基因活性下降從而抑制細胞凋亡,SIRT1的3′UTR存在miR-34a的結合位點,通過熒光素酶實驗確認其為miR-34a的靶序列,通過該序列miR-34a可以降解SIRT1mRNA從而下調SIRT1,人們通過一定方法敲除HCT116的miR-34a后能夠明顯增加SIRT1的表達,同時能明顯下調p53基因的表達并降低PUMA基因的穩定性;另外,miR-34a能夠直接作用于SIRT1mRNA的3′非編碼區,降低SIRT1蛋白的水平,而敲除后則使得乙酰化的p53增加,并上調其靶基因p21[28]?由此可見,miR-34a通過抑制SIRT1的表達,增強p53的穩定性并提高其活性?
4.2 miR-34a下調E2F3蛋白的表達影響大腸癌腫瘤細胞周期 作為細胞周期調控和凋亡過程中重要的調控分子,E2F1~3使靜止的細胞進入S期?在G1期,E2F1~3被視網膜母細胞瘤蛋白家族(Rb?p107和p130)結合抑制.在G1的中期和晚期,Rb被細胞周期素和細胞周期素依賴性激酶的復合體(cyclinD/Cdk4-Cdk6)磷酸化,Rb釋放 E2F,E2F與啟動子結合隨后激發基因的轉錄,開始G1/S期的進程轉變[29],Tazawa等[17]將miR-34a注射入裸鼠的結腸癌細胞 HCT116及RKO移植瘤,兩周后腫瘤明顯消退,發現轉染 miR-34a后E2F1以及E2F3蛋白的表達都減少?Welch等[30]研究發現在將表達E2F3預測靶目標的寡核苷酸序列克隆進熒光素酶基因的3′非編碼區后,通過向神經母細胞瘤SK-N-AS細胞系聯合轉染該熒光素酶基因和pre-miR-34a,miR-34a轉染后熒光素酶活性明顯下降?表明miR-34a直接作用于E2F3,通過下調其蛋白的表達影響細胞周期過程,從而抑制結腸癌細胞的生長?
4.3 通過反饋循環通路提高miR-34a效能 通過上面所述可以得知,SIRT1的3′UTR存在 miR-34a的結合位點,是 miR-34a的靶序列,通過該序列miR-34a可以降解SIRT1mRNA從而下調 SIRT1,進而正反饋作用于 p53構成 p53-miR-34s-SIRT1-p53正反饋調節循環?而當miR-34a含量增加后,大腸癌細胞中的E2F3會出現下調,進而又正反饋作用于p53構成p53-miR-34a-E2F-p53環 路,而 前 邊 已 經 敘 述 miR-34a可 由p53激活?從而認識到miR-34a是通過對E2F3通路及SIRT1通路的負性調節作用以及p53信號通路的正反饋調節循環來抑制腫瘤細胞增殖的[31],見圖1?
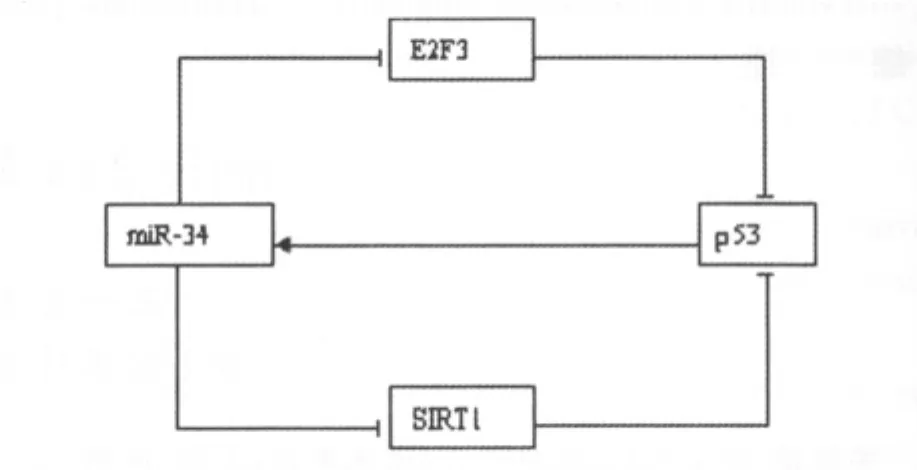
圖1 miR-34對E2F3?SIRT1的負性調節作用及p53信號通路的正反饋調節循環
5 miR-34b和miR-34c與大腸癌的發生
以上所述均為前人對miR-34a的研究,而對miR-34b和miR-34c的研究雖少但也有一個初步認識,之所以將二者同時敘述是因為miR-34b和miR-34c作為一個基因簇共同轉錄表達[32]?最近有學者對 miR-34c進行了單方面研究,并發現Myc基因是DNA損傷反應中miR-34c的主要靶點,因在人結腸癌細胞系中存在著c-myc基因的擴增,說明miR-34c可通過對DNA損傷過程的作用抑制人結腸癌細胞DNA的合成從而抑制腫瘤生長[33]?大多數人體組織中miR-34b和miR-34c的表達量小于miR-34a,只有在肺中miR-34b和miR-34c的表達量才遠遠大于miR-34a[34]?但是在大腸癌的發生及發展過程中miR-34b和miR-34c的作用同樣沒有被人們忽視?Toyota等[20]研究顯示在 CRC細胞系(9/9?100%)和原位 CRC腫瘤(101/111?90%)中 miR-34b和 miR-34c啟動子附近的 CpG島在結腸癌中常被甲基化,從而促使miR-34b和miR-34c基因的表觀沉默,此時結腸癌細胞增殖加快?然而一旦去甲基化劑作用后抑制其下游靶蛋白肝細胞生長因子受體(c-Met)?細胞周期蛋白依賴激酶4(CDK4)和SFRS2的表達,啟動子被重新激活,結腸癌細胞內即可檢測出大量 miR-34b和 miR-34c,同時結腸癌細胞增殖被明顯抑制?因CDK4是p53的作用靶點,而MET和SFRS2都與p53調節網絡有關,提示DNA甲基化導致的miR-34b/c失活也是通過p53網絡的作用來促進大腸癌的發生?發展的?因此,miR-34b和miR-34c通過p53網絡顯示出了潛在的抑癌作用,是大腸癌中新的抑癌基因?同時證明miR-34b/c CpG島是CRC中表觀沉默的重要靶點?
6 展 望
目前,對miR-34家族的研究剛剛處于起步階段,研究者對此的研究尚不夠全面,尤其是在大腸癌發展過程中miR-34a減少的原因是什么還沒有得到證實,是腫瘤發生的原因還是存在其他的調控過程而導致miR-34a的減少,還不得而知?另外,有研究發現在前列腺癌細胞中,只有miR-34a和miR-34c聯合作用才能夠產生p53介導的細胞凋亡效應[35]?所以miR-34家族成員之間的調控關系在大腸癌之中的作用還是一個未知數,也為進一步研究提供了一個方向?
綜上所述,從基因層面治療惡性腫瘤為人們在治療大腸癌方面指出了一個新的方向,如何做到不手術或者手術后如何減少復發?轉移來治療癌癥是人們一直追求的目標,而近些年對miRNAs在惡性腫瘤中的發生?發展特點的深入研究為此提供了理論依據,有理由相信,通過對miR-34s的深入研究,可以對大腸癌的發生?診斷以及治療有更深層次的認識?
[1] Bartel DP.MicroRNAs:Genomics,Biogenesis,Mechanism,and Function[J].Cell,2004,116(36):281-297.
[2] Bartel DP.MicroRNAs:Target recognition and regulatory functions[J].Cell,2009,136(2):215-233.
[3] Esquela-Kerscher A,Slack FJ.Oncomirs -microRNAs with a role in cancer[J].Nat Rev Cancer,2006,6(4):259-269.
[4] Flynt AS,Lai EC.Biological principles of microRNA-mediated regulation:shared themes amid diversity[J].Nat Rev Genet,2008,9(11):831-842.
[5] Hammond SM.MicroRNAs as oncogenes[J].Curr Opin Genet Dev,2006,16(1):4-9.
[6] Medina PP,Slack FJ.MicroRNAs and cancer:an overview[J].Cell Cycle,2008,7(16):2485-2492.
[7] Volinia S,Calin GA,Liu CG,et al.A microRNA expression signature of human solid tumors defines cancer gene targets[J].Proc Natl Acad Sci U S A,2006,103(7):2257-2261.
[8] Lewis BP,Burge CB,Bartel DP.Conserved seed pairing,often flanked by adenosines,indicates that thousands of human genes are microRNA targets[J].Cell,2005,120(1):15-20.
[9] Felli N,Fontana L,Pelosi E,et al.MicroRNAs 221and 222inhibit normal erythropoiesis and erythroleukemic cell growth via kit receptor down-modulation[J].Proc Natl Acad Sci U S A,2005,102(50):18081-18086.
[10]Pineau P,Volinia S,McJunkin K,et al.MiR-221overexpression contributes to liver tumorigenesis[J].Proc Natl Acad Sci U S A,2010,107(1):264-269.
[11]Calin GA,Sevignani C,Dumitru CD,et al.Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancer[J].ProsNatlAcad Sci U S A,2004,101(9):2999-3004.
[12]Iorio MV,Visone R,Di Leva G,et al.MicroRNA signatures in human ovarian cancer[J].Cancer Res,2007,67(18):8699-8707.
[13]Thomson JM,Newman M,Parker JS,et al.Extensive post-transcriptional regulation of microRNAs and its implications for cancer[J].Genes Dev,2006,20(16):2202-2207.
[14]Chang TC,Wentzel EA,Kent OA,et al.Transactivation of miR-34aby p53broadly influences gene expression and promotes apoptosis[J].Mol Cell,2007,26(5):745-752.
[15]Tarasov V,Jung P,Verdoodt B,et al.Differential regulation of microRNAs by p53revealed by massively parallel sequencing:miR-34ais a p53target that induces apoptosis and G1-arrest[J].Cell Cycle,2007,6(13):1586-1593.
[16]He L,He XY,Lim LP,et al.A microRNA component of the p53tumour suppressor network[J].Nature,2007,447(7148):1130-1134.
[17]Tazawa H,Tsuchiya N,Izumiya M,et al.Tumor-suppressive miR-34ainduces senescence-like growth arrest through modulation of the E2Fpathway in human colon cancer cells[J].Proc Natl Acad Sci U S A,2007,104(39):15472-15477.
[18]Bommer GT,Gerin I,Feng Y,et al.p53-mediated activation of miRNA34candidate tumor-suppressor genes[J].Curr Biol,2007,17(15):1298-1307.
[19]Lodygin D,Tarasov V,Epanchintsev A,et al.Inactivation of miR-34aby aberrant CpG methylationin multiple types of cancer[J].Cell Cycle,2008,7(16):2591-2600.
[20]Toyota M,Suzuki H,Sasaki Y,et al.Epigenetic silencing ofmicroRNA-34b/c and B-cell translocation gene 4isassociated with CpG island methylation in colorectalcancer[J].Cancer Res,2008,68(11):4123-4132.
[21]Raver-Shapira N,Marciano E,Meiri E,et al.Transcriptional activation of miR-34acontributes to p53-mediated apoptosis[J].Mol Cell,2007,26(5):731-743.
[22]Hermeking H.p53enters the microRNA world[J].Cancer Cell,2007,12(5):414-418.
[23]Corney DC,Flesken-Nikitin A,Godwin AK,et al.MicroRNA-34band MicroRNA-34care targets of p53and cooperate in control of cell proliferation and adhesion-independent growth[J].Cancer Res,2007,67(18):8433-8438.
[24]Oltersdorf T,Elmore SW,Shoemaker AR,et al.An inhibitor of Bcl-2family proteins induces regression of solid tumours[J].Nature,2005,435(7042):677-681.
[25]Han Z,Hong L,Han Y,et al.Phospho Akt mediatesmultidrug resistance of gastric cancer cells through regulation of P-gp,Bcl-2and Bax[J].J Exp Clin Cancer Res,2007,26(2):261-268.
[26]Cole KA,Attiyeh EF,Mosse YP,et al.A functional screen identifies miR-34aas a candidate neuroblastoma tumor sup-pressor gene[J].Mol Cancer Res,2008,6(5):735-742.
[27]Longo VD,Kennedy BK.Sirtuins in aging and age-related disease[J].Cell,2006,126(2):257-268.
[28]Yamakuchi M,Ferlito M,Lowenstein CJ.miR-34arepression of SIRT1regulates apoptosis[J].Proc Natl Acad Sci U S A,2008,105(36):13421-3426.
[29]Ogawa H,Ishiguro K,Gaubatz S,et al.A complex withchromatin modifiers that occupies E2F-and Myc-responsive genes in G0cells[J].Science,2002,296(5570):1132-1136.
[30]Welch C,Chen Y,Stallings RL.MicroRNA-34afunctions as a potential tumor suppressor by inducing apoptosis in neuroblastoma cells[J].Oncogene,2007,26(34):5017-5022.
[31]Wu L,Timmers C,Maiti B,et al.The E2F1-3transcription factors are essential for cellular proliferation[J].Nature,2001,414(6862):457-462.
[32]He X,He L,Hannon GJ.The guardian′s little helper:microRNAs in the p53tumor suppressor network[J].Cancer Res,2007,67(23):11099-11101.
[33]Ian G,Cannell B,Bushell M.Regulation of Myc by miR-34c[J].Cell Cycle,2010,9(14):2726-2730.
[34]Corney DC,Flesken-Nikitin A,Godwin AK,et al.MicroRNA-34band MicroRNA-34care targets of p53and cooperate in control of cell proliferation and adhesion-independent growth[J].Cancer Res,2007,67(18):8433-8438.
[35]Rokhlin OW,Scheinker VS,Taghiyev AF,et al.MicroRNA-34mediates AR-dependent p53-induced apoptosis in prostate cancer[J].Cancer Biol Ther,2008,7(8):1288-1296.
10.3969/j.issn.1671-8348.2012.18.035
A
1671-8348(2012)18-1871-04
2011-10-09
2011-12-08)
?臨床護理?
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
西南國防醫藥(2016年7期)2016-12-01 06:01:15
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
河南醫學研究(2014年3期)2014-02-27 14:51:48
沈陽醫學院學報(2014年1期)2014-02-16 06:19:24
