BDSF對白假絲酵母體外作用的研究
2012-01-23 01:12:38周阿峰汪聯輝翁麗星
微生物與感染 2012年1期
周阿峰,汪聯輝,翁麗星
1. 復旦大學生命科學學院微生物學和微生物工程系,上海 200433;2. 南京郵電大學信息材料與納米技術研究院,南京 210046
假絲酵母是一類重要的人類致病菌。在美國,假絲酵母病在醫院感染的大病中占第3或第4位,排名高于很多細菌感染[1,2]。更嚴重的是,假絲酵母病患者的病死率逐年增加,有報道稱已達40%~60%[3,4]。從無害的共生菌轉變為致病菌,這與宿主的免疫狀態密切相關。在過去的10年內,隨著免疫衰減人群(包括艾滋病患者、乙型肝炎患者、器官移植受者、腫瘤化療患者)大幅增加,假絲酵母病激增。實際上,在兒童和成人乙型肝炎、艾滋病患者中,假絲酵母病是最常見的真菌感染[5]。假絲酵母也是感染器官移植受者的主要真菌,其中假絲酵母病導致的病死率高達50%[6]。
在假絲酵母眾多致病因素中,酵母形態轉變為菌絲形態是相當重要的因素之一。白假絲酵母是雙相型單細胞致病菌,卵圓形的酵母相是非致病型,而菌絲相是致病型[7]。
目前,國內外學者的研究集中在不同物質對白假絲酵母形態轉變的影響,且主要以酵母相為起點,鮮見以菌絲相為起點。其中,對法尼醇的研究最普遍。法尼醇由白假絲酵母分泌,是白假絲酵母群體感應分子,在白假絲酵母形態轉變過程中起重要作用,達一定濃度時可阻礙其從酵母相向菌絲相轉變。法尼醇的化學結構見圖1。汪聯輝等于2008年就可擴散信號因子(diffusible signal factor,DSF)對以酵母相為起點的白假絲酵母菌絲生長的影響進行了研究[8]。
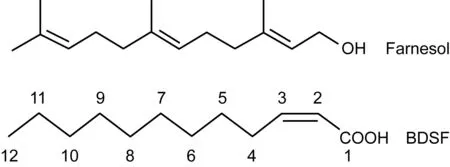
圖1 法尼醇和BDSF的結構
Fig.1 Structures of farnesol and BDSF
BDSF由Burkholderiacenocepacia分泌產生,是小分子短鏈脂肪酸。BDSF屬DSF家族,在結構上與DSF十分相似。DSF是黃單胞菌(Xanthomonascampestris)的群體感應分子[9,10]。BDSF的化學結構見圖1。近幾年,Ryan等研究發現,BDSF可影響Burkholderiacenocepacia的毒力[11,12]。
白假絲酵母形態轉變的調控網絡極其復雜,其中已闡明位于白假絲酵母形態學調控中心位置的是Ras1p相關調控網絡[11]。Ras1p是一種小GTP酶,啟動2條通路:促絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)通路和cAMP依賴的蛋白激酶A(protein kinase A,PKA)通路[13]。MAPK通路是一系列的磷酸化級聯反應:從MAPK激酶的激酶(mitogen-activated protein kinase kinase kinase,MAPKKK)至MAPK導致一系列轉錄因子激活。MAPK有5種:Cek1p、Hog1p、Mkc1p、Cek2p和Csk1p。在cAMP依賴的PKA通路中,首先腺苷酸環化酶Cyr1p催化ATP產生第二信使cAMP,cAMP水平升高激活PKA,經過一系列反應后激活轉錄因子Efg1p。該轉錄因子負責調控大多數參與形態轉變的基因[13]。
本研究分別以酵母相和菌絲相為起點,初步探討BDSF對白假絲酵母兩相形態的影響,為進一步探索BDSF的功能奠定基礎,尤以菌絲相為起點的研究具有開拓性意義。
1 材料和方法
1.1 菌株、培養基和BDSF的配制
菌株SC5314是白假絲酵母標準株,由中國科學院上海生命科學院陳江野老師惠贈。菌株保存于含40%甘油溶液的SDA液態培養基中,混勻后于-80 ℃冷凍保藏。保藏的菌株在固體SDA平板中復蘇后4 ℃保存,每月傳代1次。
復蘇后的菌株于GS培養基中23 ℃搖瓶培養48 h,95%白假絲酵母保持酵母形態[12]。將培養液4 ℃保存以備實驗使用。將備用酵母相移至新鮮GS培養基中,37 ℃靜置培養3 h后,95%轉變為菌絲相[12]。GS培養基的組成(/L):葡萄糖5 g、Na2HPO4·12H2O 0.26 g、KH2PO40.66 g、MgSO4·7H2O 0.88 g、NH4Cl 0.33 g、生物素16 μg、1%(V/V)新生牛血清(3 kD無菌濾管過濾處理)。調整pH值為7.0[13],過濾除菌后于4 ℃保存。
BDSF由復旦大學先進材料實驗室化學合成。準確稱取適量BDSF,將其溶于適量甲醇作為母液,4 ℃保存。用甲醇倍比稀釋達實驗所需濃度。法尼醇購自Sigma公司,配制方法同BDSF。
1.2 方法
1.2.1 BDSF對白假絲酵母酵母相的最低抑制濃度 實驗分為空白對照組、法尼醇組和BDSF組。空白對照組含GS培養基、白假絲酵母和溶劑甲醇;法尼醇組含GS培養基、白假絲酵母和法尼醇;BDSF組含GS培養基、白假絲酵母和BDSF。調整白假絲酵母終濃度為1×105~5×105cfu/ml,BDSF終濃度為240、120、60、45、30、20、15、7.5、3.75和1.9 μmol/L。BDSF組和法尼醇組每個濃度設3個對照,以200個細胞為計數單位,37 ℃靜置培養4 h,在倒置顯微鏡下計數酵母相或菌絲相的數量。采用SPSS軟件計算50%最低抑制濃度(50% minimum inhibitory concentration,MIC50)和MIC90。
1.2.2 BDSF對白假絲酵母酵母相的作用 實驗分為空白對照組和BDSF組。空白對照組含GS培養基、白假絲酵母;BDSF組含GS培養基、白假絲酵母和BDSF。調整白假絲酵母終濃度為1×105~5×105cfu/ml,BDSF終濃度為120、60、30 μmol/L。37 ℃靜置培養。從第4小時開始,每隔2 h拍照1次。
1.2.3 BDSF對白假絲酵母菌絲相的作用 實驗分為空白對照組和BDSF組。空白對照組含GS培養基、白假絲酵母;BDSF組含GS培養基、白假絲酵母和BDSF。調整白假絲酵母終濃度為1×104~5×105cfu/ml,BDSF終濃度為120、60、30 μmol/L。37 ℃靜置培養。BDSF組于4 h后再加入BDSF,并以此為0點,每隔3 h拍照1次。
2 結果
2.1 BDSF對白假絲酵母的MIC
以法尼醇為陽性對照,比較BDSF和法尼醇對白假絲酵母的MIC。以濃度為橫坐標、酵母相向菌絲相轉化的抑制率為縱坐標作圖 (圖2)。采用SPSS和Excel軟件處理數據,求出BDSF對白假絲酵母的MIC50為(20.7±1.6) μmol/L,MIC90為(55.1±2.8) μmol/L;而法尼醇對白假絲酵母的MIC50為(38.8±2.4) μmol/L,MIC90為(105.4±6.2) μmol/L。結果表明,對白假絲酵母菌絲生長的抑制作用BDSF比法尼醇更有效,兩者差異較顯著。
2.2 BDSF對白假絲酵母從酵母相轉為菌絲相的抑制作用
2.2.1 30 μmol/L BDSF對白假絲酵母菌絲生長的抑制 為了解4 h后白假絲酵母的生長情況及是否有新菌絲生長,在培養4 h后,每隔2 h對白假絲酵母觀察并拍照,記錄其生長情況;并以200個細胞為計數單位,對其中的菌絲計數,至10 h止。以時間為橫坐標、以酵母相向菌絲相轉化率為縱坐標作圖(圖3)。結果表明,BDSF在30 μmol/L時可有效抑制90%以上酵母相向菌絲相的轉化。

The dosage effects of BDSF and farnesol on morphology of C. albicans were determined microscopically on about 200 yeast cells in each treatment. The means and standard errors from three repeats are presented.

圖3 以酵母相為起點BDSF對白假絲酵母菌絲生長的影響Fig.3 Effect of BDSF on hyphae growth of C. albicans from yeast form
2.2.2 30、60和120 μmol/L BDSF對白假絲酵母菌絲生長的抑制 BDSF在30 μmol/L時可有效抑制白假絲酵母菌絲生長,但隨時間延長并不能抑制白假絲酵母出芽生殖。為了解增加BDSF濃度和延長培養時間對白假絲酵母菌絲生長及出芽生殖的影響,設置空白對照組和30、60和120 μmol/L BDSF組,并于4、6、8和10 h置倒置顯微鏡下拍照(圖4)。結果表明,BDSF在30、60 μmol/L時只能抑制白假絲酵母菌絲生長;當濃度增至120 μmol/L時,白假絲酵母的菌絲生長和出芽生殖均被有效抑制。原因可能是菌絲生長和出芽生殖采取不同的代謝通路,BDSF在30、60 μmol/L時只能影響菌絲生長代謝通路,而120 μmol/L時菌絲生長和出芽生殖代謝通路均受影響。

圖4 BDSF對酵母相白假絲酵母形態轉變的劑量和時間效應
Fig.4 Dosage and time effects of BDSF on morphology of yeastC.albicans
2.3 BDSF對白假絲酵母從菌絲相轉為酵母相的促進作用
白假絲酵母在酵母相和菌絲相之間可逆轉變。一般認為,酵母相為非致病形態,而菌絲相為致病形態。白假絲酵母從酵母相轉為菌絲相,毒性增加[7]。BDSF對已形成菌絲相的白假絲酵母影響見圖5。空白組中,24 h內隨時間增加,白假絲酵母菌絲生長旺盛,新生菌絲不斷延長。BDSF在 30、60 μmol/L時可促使新生菌絲不斷轉變為酵母相;在120 μmol/L時,白假絲酵母的菌絲生長受強烈抑制,菌絲保持最初形態,無新生菌絲。
3 討論
以酵母相為起點,當BDSF≥30 μmol/L時菌絲生長受強烈抑制。以菌絲相為起點,在30和60 μmol/L時,BDSF并不抑制菌絲進一步生長和產生分支,但隨菌絲分支生長,新生的分支菌絲不斷轉變為酵母相;當BDSF增至120 μmol/L時,菌絲生長和分支狀況幾乎完全受抑制。
以酵母相為起點,用不同物質影響白假絲酵母菌絲生長的研究較為普遍。2001年Oh等發現法尼酸可抑制酵母相白假絲酵母的菌絲生長[12],并將其視作調節白假絲酵母形態轉變的自誘導物質;但后來證實為法尼醇。2008年汪聯輝等發現DSF可抑制酵母相白假絲酵母的菌絲生長[8]。以菌絲相為起點,用不同物質影響白假絲酵母菌絲生長的研究較罕見。2001年Oh等曾發表觀點:法尼酸可逆轉白假絲酵母從菌絲相轉為酵母相[14],但至今并無充分證據。本文研究結果表明,BDSF對白假絲酵母從酵母相轉為菌絲相起抑制作用,對從菌絲相轉為酵母相起促進作用。
位于白假絲酵母形態學調控中心的是Ras1p相關調控網絡,其中研究較細致的是MAPK通路和cAMP依賴的PKA通路。2004年Sato等證實,法尼醇經MAPK通路抑制白假絲酵母菌絲生長[15]。至于BDSF是否經其中1個或2個通路,抑或其他通路對白假絲酵母菌絲生長產生抑制作用,有待進一步實驗證實。
白假絲酵母致病時多在病灶部位形成生物膜。該生物膜是由酵母相、假菌絲和真菌絲3種形態相互交織形成的致密結構。BDSF在實驗室條件下可有效抑制菌絲生長并促使菌絲相轉化為酵母相,為防治白假絲酵母病提供了嶄新思路。但臨床上能否攻克生物膜這個堡壘,尚需進一步研究。

圖5 BDSF對菌絲相白假絲酵母形態轉變的劑量和時間效應
Fig.5 Dosage and time effects of BDSF on morphology of hyphaeC.albicans
[1] Banerjee SN, Emori TG, Culver DH, Gaynes RP, Jarvis WR, Horan T, Edwards JR, Tolson J, Henderson T, Martone WJ. Secular trends in nosocomial primary bloodstream infections in the United States, 1980-1989 [J]. Am J Med, 1991, 91(3B): 86S-89S.
[2] Edmond MB, Wallace SE, McClish DK, Pfaller MA, Jones RN, Wenzel RP. Nosocomial bloodstream infections in United States hospitals: a three-year analysis [J]. Clin Infect Dis, 1999, 29(2): 239-244.
[3] Wenzel RP, Gennings C. Bloodstream infections due to Candida species in the intensive care unit: identifying especially high-risk patients to determine prevention strategies [J]. Clin Infect Dis, 2005, 41(Suppl 6): S389-S393.
[4] Colombo AL, Nucci M, Park BJ, Nouér SA, Arthington-Skaggs B, da Matta DA, Warnock D, Morgan J, Brazilian Network Candidemia Study. Epidemiology of candidemia in Brazil: a nationwide sentinel surveillance of candidemia in eleven medical centers [J]. J Clin Microbiol, 2006, 44(8): 2816-2823.
[5] Samaranayake LP, Fidel PL, Naglik JR, Sweet SP, Teanpaisan R, Coogan MM, Blignaut E, Wanzala P. Fungal infections associated with HIV infection [J]. Oral Dis, 2002, 8(Suppl 2): 151-160.
[6] Patterson TF. Approaches to fungal diagnosis in transplantation [J]. Transpl Infect Dis, 1999, 1(4): 262-272.
[7] Lo HJ, K?hler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, Fink GR. Nonfilamentous C. albicans mutants are avirulent [J]. Cell, 1997, 90(5): 939-949.
[8] Boon C, Deng Y, Wang LH, He Y, Xu JL, Fan Y, Pan SQ, Zhang LH. A novel DSF-like signal from Burkholderia cenocepacia interferes with Candida albicans morphological transition [J]. ISME J, 2008, 2(1): 27-36.
[9] He YW, Wu J, Cha JS, Zhang LH. Rice bacterial blight pathogen Xanthomonas oryzae pv. oryzae produces multiple DSF-family signals in regulation of virulence factor production [J]. BMC Micobiol, 2010, 10: 187.
[10] He YW, Boon C, Zhou L, Zhang LH. Co-regulation of Xanthomonas campestris virulence by quorum sensing and a novel two-component regulatory system RavS/RavR [J]. Mol Microbiol, 2009, 71(6): 1464-1476.
[11] Ryan RP, McCarthy Y, Watt SA, Niehaus K, Dow JM. Intraspecies signaling involving the diffusible signal factor BDSF (cis-2-dodecenoic acid) influences virulence in Burkholderia cenocepacia [J]. J Bacteriol, 2009, 191(15): 5013-5019.
[12] McCarthy Y, Yang L, Twomey KB, Sass A, Tolker-Nielsen T, Mathenthiralingam E, Dow JM, Ryan RP. A sensor kinase recognizing the cell-cell signal BDSF (cis-2-dodecenoic acid) regulates virulence in Burkholderia cenocepacia [J]. Mol Microbiol, 2010, 77(5): 1220-1236.
[13] Hall RA, Cottier F, Mühlschlegel FA. Molecular networks in the fungal pathogen Candida albicans [J]. Adv Appl Microbiol, 2009, 67: 191-212.
[14] Oh KB, Miyazawa H, Naito T, Matsuoka H. Purification and characterization of an autoregulatory substance capable of regulating the morphological transition in Candida albicans [J]. Proc Natl Acad Sci USA, 2001, 98(8): 4664-4668.
[15] Sato T, Watanabe T, Mikami T, Matsumoto T. Farnesol, a morphogenetic autoregulatory substance in the dimorphic fungus Candida albicans, inhibits hyphae growth through suppression of a mitogen-activated protein kinase cascade [J]. Biol Pharm Bull, 2004, 27(5): 751-752.
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
小讀者(2021年2期)2021-03-29 05:03:48
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
