楊樹腐爛病內生拮抗細菌鑒定及防治研究
2012-01-22 06:41:04金海強賈斌李熙英
中國森林病蟲 2012年6期
金海強,賈斌,李熙英
(延邊大學教育部長白山生物資源與功能分子重點實驗室,吉林延吉 133002)
楊樹Populus spp.是我國北方地區速生豐產造林樹種之一,具有防風固沙、水土保持,農田保護和維護生態平衡等作用。楊樹腐爛病Cytospora chrysosperma等多種枝干病害經常發生,直接危害楊樹生長。目前對楊樹腐爛病的防治主要依靠物理、化學防治并結合抗病品種的選用等。化學農藥見效快,防治效果好,但成本較高,易造成環境污染,病原菌易產生抗藥性等[1]。以往楊樹腐爛病的生物防治主要是篩選利用拮抗菌,但存在拮抗菌定植率差和防效不高、不穩定等問題[2-6]。近年來的研究結果表明,在健康的植物體中存在大量的內生拮抗細菌,它們是植物病害防治的潛在資源菌[7-8]。植物內生細菌長期生活于植物體內特殊環境并與植物協同進化,建立了和諧聯合的關系。一方面植物為內生細菌提供了良好的棲息地,另一方面內生細菌又可通過自身代謝產物或借助于信號傳導作用對植物產生影響,對寄主植物有促生、防病等多方面的生物學作用[9]。本研究從健康楊樹枝條中分離篩選出對楊樹腐爛病具有拮抗作用的內生細菌,為楊樹腐爛病生物防治提供依據。
1 材料與方法
1.1 供試菌種 楊樹腐爛病病原菌Cytospora chrysosperma來自延邊大學農學院植物病理實驗室。
1.2 內生細菌分離材料 分別于春季的3月中旬、夏季的7月中旬和秋季的10月中旬從吉林省延邊地區的延吉、龍井、安圖、和龍等地的楊樹林(品種為大青楊)中采集楊樹枝條,隨機取樣。樹齡為5~7 a,枝條直徑1 cm左右,長20 cm。
1.3 楊樹內生細菌的分離及純化 取不同地點采集到的楊樹枝條,用70%的酒精表面清洗后,剪成0.5~1 cm的枝段,浸泡在70%乙醇中1 min,再用3.25%的NaOCl浸泡15 min,用無菌蒸餾水沖洗4次,放置在PDA平板上培養2 d。
從上述培養基平板里挑出沒長出微生物菌落的(說明表面消毒徹底)楊樹枝條段5g放入已滅菌的研缽中,加入45 mL無菌水,碾碎,攪拌5 min,靜止15 min,按稀釋法稀釋后用10-3,10-4的稀釋液涂抹于PDA和NA培養基平板上,放入25℃的恒溫箱中培養。每處理重復5個培養皿。培養第5天,根據菌落形態、顏色、質地等調查微生物菌株數和每種微生物的菌落數,并對每種微生物進行純培養以作為參試菌株。
1.4 室內拮抗菌的篩選 在PDA平板上培養7 d的楊樹腐爛病菌菌落上,用打孔器打出直徑5 mm的菌碟,放置于新的PDA平板中間,在距培養皿邊緣10 mm處點接內生細菌菌株(2個點對稱),設只接病原菌的為對照,置于25℃恒溫箱中培養。每個處理重復5次。培養第5天測楊樹腐爛病菌菌落寬度,并計算抑菌率。

1.5 拮抗菌發酵液的抑菌活性 選擇在對峙培養中對楊樹腐爛病菌具有較強抑菌活性的14種內生拮抗菌,用KB培養基活化培養3 d后,分別接種于200 mL KB液體培養基中,在28℃、120 r/min恒溫雙層搖床中振蕩培養5 d。在經4 000 r/min離心25 min后,取上清液用直徑0.22 μm微孔濾膜過濾后,用旋轉蒸發儀濃縮至10 mL,即為拮抗菌發酵液。
抑菌活性測定采用紙片法。具體方法如下:先在PDA平板中間放置直徑5 mm的楊樹腐爛病菌的菌碟1片,在距離菌碟10 mm處放置滴150 μL拮抗菌發酵液的濾紙片2片(2片對稱),以只放置楊樹腐爛病菌菌碟的為對照,放入25℃恒溫箱培養。每個處理重復5次。培養5 d后測楊樹腐爛病菌菌落寬度,并計算抑菌率。
1.6 離體枝條防病試驗 在PDA平板上培養7 d的楊樹腐爛病菌菌落上,用打孔器打出直徑5 mm的菌碟,備用。
將14種內生拮抗菌在KB培養基上活化培養3 d后,分別接入100 mL KB液體培養基中,在28℃、120 r/min恒溫雙層搖床中振蕩培養5 d,備用。
將直徑1.5~2.5 cm楊樹枝條剪成段,每段長30 cm。用75%酒精表面消毒,在距兩頭5 cm處用消毒好的解剖刀劃開“T”字形口,先滴內生拮抗菌培養液1 mL,然后接楊樹腐爛病菌菌碟1片。用蘸無菌水的脫脂棉保濕,用膠布將其固定,枝條兩頭同樣需用蘸水脫脂棉保濕。每一組有30個枝條。其中只接腐爛病病菌的枝條為對照。最后每種處理放在一個塑料袋中,在室溫下(21℃左右)培養。40 d后調查發病率、病斑大小、病級及愈傷組織形成情況。楊樹腐爛病病級調查標準參見文獻[10]。

1.7 拮抗菌菌株的鑒定
1.7.1 拮抗菌形態及生理生化特性 將菌株Y-SY12接種在NA平板上,培養1 d后進行革蘭氏染色,觀察菌體形態和菌落特征。
1.7.2 16S rDNA基因序列及系統發育分析 以菌株Y-S-Y12 DNA為模板,利用16S rDNA通用引物對 27F-FOR(5′-GAGTTTGATCCTGGCTCAG-3)和1492R-REV(5′-TACGGTTACCTTGTTACGACTT-3′)為探針進行16S rDNA分析。PCR擴增反應條件為:94℃ 變性 5 min;94℃ 變性 40 s,55℃ 復性40 s,72℃延伸1 min,循環數為35。PCR 產物經瓊脂糖凝膠電泳回收純化后,將擴增得到的片段連接到pMD18-T載體上,并轉化大腸桿菌Escherichia coli DH5α后測序。所得16S rDNA片段序列在NCBI GenBank上利用BLASTN模塊進行BLAST,對16S rDNA序列進行同源性分析;根據BLAST結果,將與Y-S-Y12在16S rDNA序列上的同源性超過96.6%的前20個菌株的16S rDNA序列利用DNASATR軟件構建系統發育樹。
2 結果與分析
2.1 楊樹內生細菌的分離 供試2種培養基上均能分離到內生細菌,其中PDA培養基上分離到的內生細菌菌株數多于NA培養基上;NA培養基上出現的內生細菌菌落數多于PDA培養基上。從不同時間段上分離到的內生細菌菌株數和菌落數量上看,春季分離到的最少,夏季分離到的最多,秋季分離到的比夏季少(見表1)。
采用平板稀釋法,從大青楊楊樹枝條中共分離純化得到56株內生細菌菌株。

表1 不同季節楊樹枝條中分離到的內生細菌菌株數與菌落數
2.2 對楊樹腐爛病菌具有高效抑菌作用的內生拮抗菌篩選 分離到的56株內生細菌菌株與楊樹腐爛病菌進行室內對峙培養,結果23株具有拮抗作用,其中14株內生拮抗菌菌株的抑菌率達到了60%以上(見表2)。Y-S-Y2,Y-S-Y12的抑菌率最高,達到了 100%;其次為 Y-S-Y3,抑菌率為92.68%;再次為Y-S-Y1,抑菌率為73.17%;其余的抑菌率均在60%~70%之間。

表2 楊樹內生拮抗菌對楊樹腐爛病菌的室內抑菌效果
2.3 拮抗菌發酵液對楊樹腐爛病菌的抑菌活性供試14種內生拮抗菌菌株發酵液對楊樹腐爛病菌均具有一定的抑菌活性,其中Y-S-Y12菌株發酵液的抑菌活性顯著高于其它內生拮抗菌代謝產物的粗提取液的抑菌活性,其抑菌率為67.78%;其次Y-SY2,Y-S-Y8,Y-S-Y13,這三者之間沒有顯著性差異。另外,Y-S-Y7,Y-S-Y9,Y-S-Y10,Y-S-Y18,YS-Y20等5種也具有較高的抑菌活性,其抑菌率均為50%以上。詳見表3。

表3 楊樹內生拮抗菌發酵液對楊樹腐爛病菌的抑菌活性
2.4 離體枝條防病試驗 結果見表4。
從發病率上看,除Y-S-Y7處理的發病率與對照之間沒有顯著性差異外,其它各處理與對照之間均有顯著性差異。其中以Y-S-Y12處理的發病率最低,為31.67%;其次為Y-S-Y2;再次為Y-S-Y13和Y-S-Y1。從防效上看,各處理均有一定的防治效果,其中Y-S-Y12處理的防效最高,為84.95%;其次為Y-S-Y2,防效為80.64%;再次為Y-S-Y1和YS-Y13,防效分別為73.11%和71.05%,之間沒有顯著性差異。這幾種防效高的處理枝條愈傷組織形成情況也較好,其中Y-S-Y12和Y-S-Y2處理的愈傷組織形成率分別為66.2%和62.2%;Y-S-Y1和YS-Y13處理的愈傷組織形成率分別為50.3%和52.4%。這說明,防病效果較高的內生拮抗細菌對楊樹枝條形成愈傷組織也有促進作用。
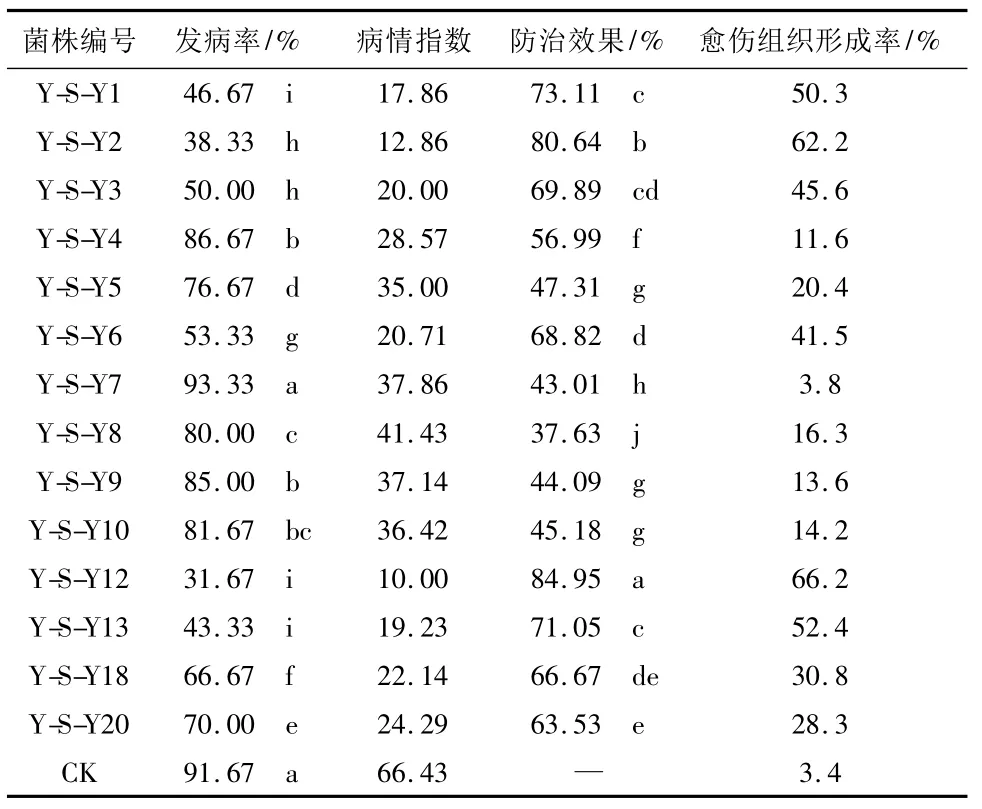
表4 離體枝條上內生拮抗菌對楊樹腐爛病的防病效果
2.5 拮抗菌的鑒定
2.5.1 拮抗菌形態及培養特征 在室內抑菌試驗和楊樹枝條防病試驗中表現突出的Y-S-Y12菌株的形態特征和培養特征:在NA平板上菌落表面粗糙不透明,有褶皺,乳白色至淡黃色。光學顯微鏡下觀察細胞呈短桿狀,大小為(0.6~0.7)μm×(2.0~2.5)μm,產芽孢,革蘭氏染色陽性。
2.5.2 16S rDNA序列及系統發育分析 在NCBI Genbank上進行的16S rDNA序列分析的結果表明,本研究中所分離的Y-S-Y12菌株屬于芽孢桿菌屬Bacillus;與Y-S-Y12菌株16S rDNA序列(Gen-Bank登錄號為JX094152)同源性超過99.5%的菌株有 136個。其中,解淀粉芽孢桿菌 Bacillus amyloli-quefaciens菌株 SWM1的16S rDNA序列(GenBank登錄號為JN851189)與Y-S-Y12菌株的同源性最高,達99.8%(見圖1、表4),其BLAST得分(Max Score)為2778,E-value為0;這兩個菌株的16S rDNA序列的目標序列1 513 bp中僅有3個堿基存在差異。雖然Y-S-Y12與芽孢桿菌屬的其他菌株B.subtilis(菌株Aj080718IA-2)及 GJ24在16S rDNA序列上同源性也達到了99.7%,但在菌落特性上存在不同。結合形態學及培養特征,菌株Y-SY12初步鑒定為解淀粉芽孢桿菌Bacillus amyloliquefaciens。

圖1 拮抗菌株Y-S-Y12與相關菌株16S rDNA序列的系統發育樹

表4 Y-S-Y12與芽孢桿菌屬20個菌(株)的16S rDNA序列一致性Percent ldentity
3 結論及討論
本試驗中,在各地采集到的楊樹枝條中分離純化得到56株內生細菌。與楊樹腐爛病菌進行室內對峙培養中,結果23株具有拮抗作用,其中14株內生拮抗菌的抑菌率60%以上。
通過室內對峙培養、檢測內生拮抗細菌發酵液對楊樹腐爛病菌的抑菌作用以及室內離體枝條防病試驗,楊樹內生拮抗細菌Y-S-Y12菌株對楊樹腐爛病的抑菌防病作用最顯著,其次為Y-S-Y2菌株。對Y-S-Y12菌株采用16S rDNA的序列分析方法進行鑒定,結果為解淀粉芽孢桿菌。
解淀粉芽孢桿菌能產生脂肽類抗生素、聚酮類抗生素以及抗菌蛋白等多種抗菌物質,這些代謝產物具有廣泛抑制植物病原真菌[11-12]和植物病原細菌[13-14]的活性,但還沒發現有關抑制楊樹腐爛病菌的報道。本試驗中分離到的解淀粉芽孢桿菌在室內條件下對楊樹腐爛病有明顯的防病作用,但在田間的防效如何、對楊樹生長有沒有促進作用,對其他病害有沒有防病作用、如何開發生物農藥以及使用方法等有待于進一步研究。
[1] 王光亮,于金友,石玉萍,等.植物病害生物防治研究進展[J].山東農業科學,2004(4):75-77.
[2] 項存悌,高克祥,何秉章,等.楊樹爛皮病生物防治的研究[J].東北林業大學學報,1999,19(6):15-25.
[3] 孫冬梅,楊謙,宋金柱.黃綠木霉菌代謝產物對楊樹爛皮病菌抑菌能力的研究[J].北京林業大學學報,2006,28(1):76-79.
[4] 原彝輯,曹支敏.楊樹腐爛病生防放線菌篩選及其活性產物研究[J].林業科學研究,2006,19(6):139-144.
[5] 任嘉紅,王艷芳,葉建仁.楊樹腐爛病拮抗細菌的篩選及其定殖研究[J].西部林業科學,2009,38(2):6-11.
[6] 袁秀英,白紅霞,白玉明,等.楊樹內生真菌的分離與拮抗生防菌的篩選[J]西北農林科技大學學報:自然科學版,2007,35(4):713-717.
[7] 鮑敏,康明浩.植物內生細菌研究發展現狀[J].青海草業,2011,20(1):21-25.
[8] 林玲,喬勇升,顧本康,等.植物內生細菌及其生物防治植物病害的研究進展[J].江蘇農業學報,2008,24(6):969-974
[9] 史應武,婁愷,李春.植物內生細菌在生物防治中的應用[J].微生物學雜志,2011,29(6):61-66.
[10] 宋瑞清,冀瑞卿.四種毒蘑菇菌株及其毒素對楊樹爛皮病菌生長的抑制作用[J].北京林業大學學報,2005,27(2):88-91.
[11] 陳士云,楊寶玉,高梅影.一株抑制油菜核盤菌菌核形成的解淀粉芽孢桿菌[J].應用與環境生物學報,2005,11(3):373-376.
[12] 權春善,王軍華,徐洪濤,等.一株抗真菌解淀粉芽孢桿菌的分離鑒定及其發酵條件的初步研究[J].微生物學報,2006,46(6):7-12.
[13] 張榮勝,劉永鋒,陳志誼.水稻細菌性條斑病菌拮抗細菌的篩選、評價與應用研究[J].中國生物防治學報,2011,27(4):510-514.
[14] 王小兵,駱永明,劉五星.花生青枯病內生拮抗細菌的鑒定、抗菌活性及其田間防效[J].中國生物防治學報,2011,27(1):88-92.
