桃葉杜鵑菌根顯微結(jié)構(gòu)及侵染情況
2012-01-08 11:06:23劉仁陽(yáng)
中南林業(yè)科技大學(xué)學(xué)報(bào) 2012年11期
歐 靜 ,劉仁陽(yáng) ,陳 訓(xùn)
桃葉杜鵑菌根顯微結(jié)構(gòu)及侵染情況
歐 靜1,劉仁陽(yáng)1,陳 訓(xùn)2
(1.貴州大學(xué) 林學(xué)院,貴州 貴陽(yáng) 550025;2.貴州省科學(xué)院,貴州 貴陽(yáng) 550001)
采用酸性品紅染色法對(duì)貴州不同地域分布的桃葉杜鵑根系染色壓片,對(duì)其菌根進(jìn)行顯微結(jié)構(gòu)及菌根侵染情況觀察。結(jié)果表明,野生桃葉杜鵑菌根結(jié)構(gòu)復(fù)雜,根外被密集的菌絲包被形成菌絲鞘,無(wú)根毛,在菌根中具有杜鵑花類(lèi)菌根特有典型結(jié)構(gòu)菌絲結(jié),也有穿梭于細(xì)胞間的有隔菌絲和束狀菌絲等結(jié)構(gòu)。野生植株菌根侵染指標(biāo)較高;不同地域的菌根侵染指標(biāo)差異顯著,貴陽(yáng)鹿沖關(guān)成年開(kāi)花植株高于百里杜鵑方家坪成年開(kāi)花植株;不同季節(jié)侵染指標(biāo)兩地春季高于夏季。分布于百里杜鵑的開(kāi)花成年樹(shù)侵染指標(biāo)明顯低于幼樹(shù)。播種苗侵染指標(biāo)較低,2a播種苗侵染指標(biāo)較1a呈上升趨勢(shì)。
桃葉杜鵑菌根;杜鵑花類(lèi)菌根;顯微結(jié)構(gòu);侵染指標(biāo)
杜鵑花是我們傳統(tǒng)十大名花之一,在國(guó)際園藝觀賞植物中占有重要地位[1]。許多杜鵑花生長(zhǎng)于土壤貧瘠,晝夜溫差大的惡劣環(huán)境,其中菌根起到重要的作用。杜鵑花類(lèi)菌根(Ericoid Mycorrhiza,ERM),又稱(chēng)為擬歐石楠類(lèi)菌根,是杜鵑花科除漿果鵑屬Arbutus、熊果屬Arctostaphylos外植物根系與一些土壤真菌形成典型特征的共生體[2]。國(guó)外有較多報(bào)道證明ERM對(duì)杜鵑花類(lèi)植物克服惡劣環(huán)境、加強(qiáng)吸收養(yǎng)分起到很重要的作用[3-5],接種菌株能大大提高杜鵑花的生長(zhǎng)量[6-7]。我國(guó)雖然是杜鵑花的故鄉(xiāng),但關(guān)于ERM的研究卻極少[8-9]。ERM皮層細(xì)胞中有不同于其它菌根的特殊結(jié)構(gòu),即菌絲結(jié)(hyphal coil)[2]。據(jù)資料,杜鵑花科擬歐石楠Calluna vulgaris 、越橘Vaccinium spp.、北美杜鵑花Rhododendron spp.、彭土杜鵑Rh. ponticum、馬醉木Pieris spp.、石巖杜鵑變種 Rh.obtusum var.Kaempferi、云錦杜鵑Rh. fortunei、銀葉杜鵑Rh.argyrophyllum 、繁花杜鵑Rh. floribundum等植物根系內(nèi)具有ERM典型結(jié)構(gòu)菌絲結(jié)[8-12]。
桃葉杜鵑Rhododendron annae Franch.,分布于我國(guó)西南地區(qū),花清雅高潔或艷麗奪目,葉形美觀,樹(shù)冠圓滿(mǎn),花期5~6月,作為園藝觀賞花卉極有開(kāi)發(fā)潛力。近年來(lái)高山常綠杜鵑花得到了較多的重視[13-14],但對(duì)桃葉杜鵑的研究報(bào)道較少[15-16]。本文對(duì)桃葉杜鵑菌根顯微結(jié)構(gòu)及侵染情況進(jìn)行報(bào)道,為桃葉杜鵑菌根化育苗及資源保護(hù)與利用提供基礎(chǔ)依據(jù),并對(duì)豐富ERM資源有較大意義。
1 材料與方法
1.1 采樣地概況
根樣材料分別采自貴州省貴陽(yáng)市鹿沖關(guān)森林公園桃葉杜鵑模式標(biāo)本產(chǎn)地和貴州百里杜鵑風(fēng)景區(qū)普底方家坪桃葉杜鵑分布區(qū)。鹿沖關(guān)桃葉杜鵑產(chǎn)地位于東經(jīng) 106°43′17″, 北緯 26°38′12″,海拔1 360~1 390 m,山地黃壤土,土層厚度30~80 cm,pH4.8~5.1,種群優(yōu)勢(shì)種為馬尾松Pinus massoniana 和白櫟Quercus fabri ,桃葉杜鵑數(shù)量較少,且?guī)缀鯙槌赡陿?shù),幼樹(shù)少見(jiàn);百里杜鵑方家坪桃葉杜鵑分布區(qū)位于東經(jīng)105°53′30″,北緯27°14′51″,海拔 1 800 ~ 1 830 m,硅質(zhì)黃壤土,土層厚度20~50 cm,pH4.5~5.0,幾乎為桃葉杜鵑純林。
1.2 采樣與試驗(yàn)方法
實(shí)驗(yàn)樣地20 m×20 m,選擇5株樣本植株進(jìn)行采集根樣,記錄株高、地徑,鹿沖關(guān)菌根樣編號(hào)為L(zhǎng)CG-T,百里杜鵑方家坪菌根樣編號(hào)為BL-T。分別在春、夏兩季定點(diǎn)定樣兩批采樣。3次重復(fù),每重復(fù)樣株之間距離100 m以上。1 a、2 a苗取自百里杜鵑風(fēng)景區(qū)杜鵑苗圃,播種基質(zhì)混有杜鵑林下土壤。樣根均帶土保濕放置于冰桶內(nèi)帶回實(shí)驗(yàn)室,存放4℃冰箱中備用。
菌根侵染觀測(cè)采用酸性品紅染色鏡檢法。步驟采用Komanik P P. 等(1980)方法[17]:洗根-透明-漂洗-軟化-酸化-染色-脫色;隨機(jī)取50根段,每根段8 mm左右長(zhǎng),5個(gè)一組鏡檢。3次重復(fù)。
1.3 侵染記錄與計(jì)算
記錄菌根侵染情況,計(jì)算侵染指標(biāo)[18]。侵染分級(jí)標(biāo)準(zhǔn)分為0級(jí)(侵染比例0%)、1級(jí)(侵染比例<1%)、2 級(jí)(侵染比例<10%)、3級(jí)(侵染比例<50%)、4級(jí)(侵染比例<80%)、5級(jí)(侵染比例>90%);菌絲結(jié)豐富度(豐度)劃分為E0:沒(méi)有,E1:很少,E2:較多,E3:豐富。
侵染率F%=受侵染的根段數(shù)/觀察的根總段數(shù)*100;
整個(gè)根系的菌根侵染強(qiáng)度M%=(95n5+70n4+30n3+5n2+n1)/全部根段數(shù)(n5表示:5級(jí)侵染的根段數(shù);n4表示:4級(jí)侵染的根段數(shù),等等);
侵染根段的菌根侵染強(qiáng)度m%=M*全部根段數(shù)/侵染根段數(shù);
侵染根段的菌絲結(jié)豐度e%=(100mE3+50mE2+10mE1)/100;mE=((95n5E3+70n4E3+30n3E3+5n2E3+n1E3)/n)*100/m ;
整個(gè)根系的菌絲結(jié)豐度E%=e*M/100。
1.4 統(tǒng)計(jì)分析
使用EXCEL和SPSS軟件進(jìn)行數(shù)據(jù)統(tǒng)計(jì)分析及作圖。
2 結(jié)果與分析
2.1 桃葉杜鵑菌根的形態(tài)結(jié)構(gòu)及菌絲體形態(tài)
桃葉杜鵑根系由大量細(xì)弱如發(fā)的側(cè)根組成,因此被稱(chēng)作發(fā)根。新生根為乳白色至淡黃色,老根大部分為棕色至深棕色。經(jīng)過(guò)透明染色后,觀察到側(cè)根結(jié)構(gòu)簡(jiǎn)單,由1層表皮細(xì)胞、1-2層皮層細(xì)胞和中柱組成,其根部被密集的根外菌絲所包圍,形成菌絲鞘hyphal sheath(又稱(chēng)為菌套,見(jiàn)圖1.A),無(wú)根毛結(jié)構(gòu)。
被真菌侵染的菌根可見(jiàn)細(xì)胞增大現(xiàn)象,發(fā)根本身結(jié)構(gòu)并未改變。菌根表皮細(xì)胞及皮層細(xì)胞內(nèi)可見(jiàn)不同顏色深淺的菌絲,可見(jiàn)ERM特殊結(jié)構(gòu)菌絲結(jié)hyphal coil (又稱(chēng)為菌絲團(tuán)或菌絲卷,見(jiàn)圖1.BCL)。菌絲結(jié)大多在皮層細(xì)胞中央分布,有些幾乎充滿(mǎn)整個(gè)細(xì)胞(見(jiàn)圖1.C)。有些菌根的菌絲結(jié)只單獨(dú)在一個(gè)細(xì)胞內(nèi)分布,而有的真菌菌絲體會(huì)穿過(guò)細(xì)胞進(jìn)入另一個(gè)細(xì)胞內(nèi)。菌絲復(fù)合體形態(tài)為菌絲群隨意聚集、打結(jié)、延伸、反曲(見(jiàn)圖1.DE)。入侵點(diǎn)在生長(zhǎng)區(qū)與伸長(zhǎng)區(qū)均有見(jiàn),入侵菌絲在侵入點(diǎn)附近較細(xì),進(jìn)入細(xì)胞內(nèi)部后明顯變粗,侵入點(diǎn)的菌絲有單菌絲(見(jiàn)圖1.C,箭頭所示為入侵點(diǎn))和多菌絲等形式(見(jiàn)圖1.F,箭頭所示為入侵點(diǎn))。個(gè)別根的皮層內(nèi)觀察到泡囊結(jié)構(gòu)(見(jiàn)圖1.G)。
在桃葉杜鵑根內(nèi)還發(fā)現(xiàn)在側(cè)根的分支處可見(jiàn)較多的染色較深的物質(zhì)積累 (見(jiàn)圖1.H)。穿梭于細(xì)胞間的有隔菌絲(見(jiàn)圖1.I,箭頭示隔)和束狀菌絲(見(jiàn)圖1.J)也在樣根中發(fā)現(xiàn)。另外,在同一個(gè)菌根上觀察到分布于細(xì)胞內(nèi)、細(xì)胞間和細(xì)胞外的菌絲結(jié)構(gòu)(見(jiàn)圖1.K)。
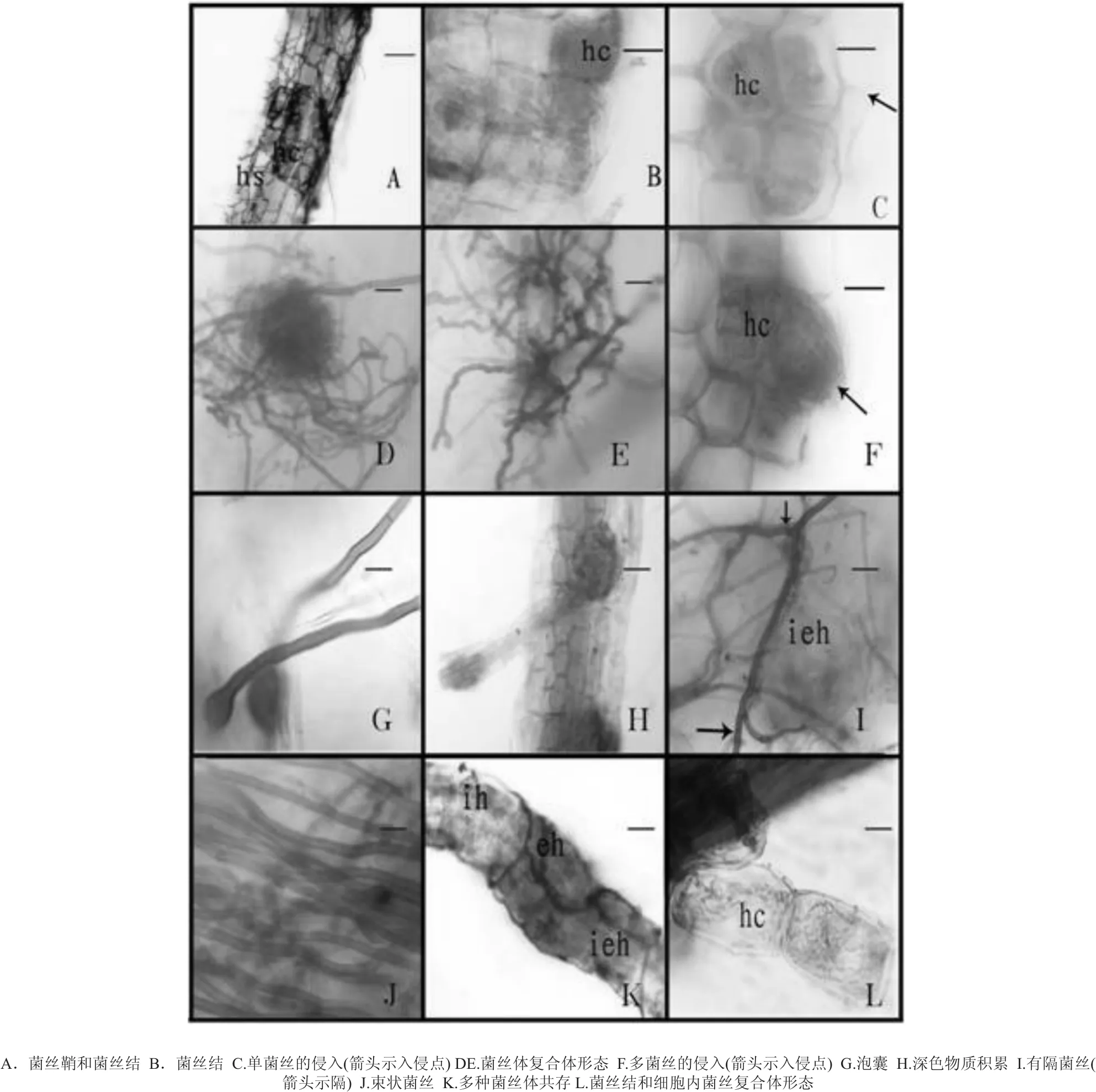
圖1 桃葉杜鵑側(cè)根顯微結(jié)構(gòu)(A,H,K標(biāo)尺=25 μm;其它標(biāo)尺=15 μm)Fig.1 Microstructure of hair root of R. annae (A,H,K Bars correspond to 25μm;other Bars correspond to 15μm)
2.2 不同分布地桃葉杜鵑菌根形態(tài)結(jié)構(gòu)
從表1可能看出,在不同的分布地,桃葉杜鵑菌根的形態(tài)結(jié)構(gòu)有所差異。百里杜鵑分布區(qū)的樣根內(nèi)大多數(shù)具有ERM典型結(jié)構(gòu)菌絲結(jié),而在鹿沖關(guān)分布區(qū)則觀察到更多復(fù)雜的菌根結(jié)構(gòu)。
2.3 桃葉杜鵑根系真菌侵染情況
2.3.1 不同分布地野生桃葉杜鵑根系真菌侵染比較
對(duì)不同分布地,株高與地徑相近的野生桃葉杜鵑菌根侵染情況測(cè)定計(jì)算。結(jié)果表明,兩地桃葉杜鵑根系均完全形成菌根,侵染率為100%(見(jiàn)表2)。從圖2可以看出,LCG-T菌根春夏兩季其它侵染指標(biāo)顯著大于BL-T菌根(P<0.05)。LCG-T1全根系及侵染根系的侵染強(qiáng)度、侵染根系的菌絲結(jié)豐度指標(biāo)分別比BL-T1高出32.64%、10.93%、31.9% 。LCG-T2全根系及侵染根系的侵染強(qiáng)度、侵染根系的菌絲結(jié)豐度指標(biāo)分別比BL-T2高出39.2%、25.77%、44.54%。

表1 不同分布區(qū)桃葉杜鵑菌根形態(tài)?Table 1 Mycorrhizal morphology of R. annae from different distribution regiongs
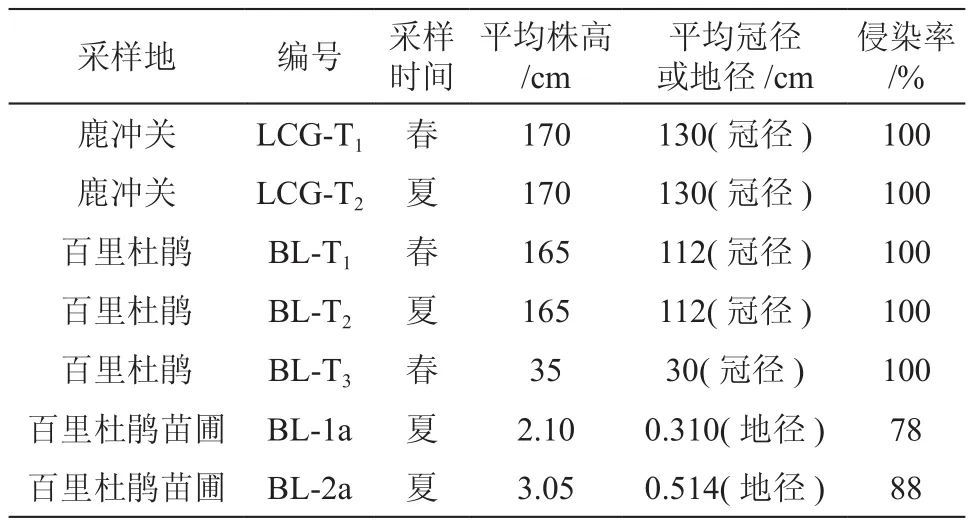
表2 桃葉杜鵑菌根侵染率Table 2 Comparison of mycorrhiza infection of R. annae

圖2 不同分布區(qū)菌根侵染比較Fig.2 Mycorrhizal colonization in different distribution area
2.3.2 不同季節(jié)野生桃葉杜鵑菌根真菌侵染比較
分 析 春 季 (LCG-T1、BL-T1)和 夏 季(LCG-T2、BL-T2)兩地野生桃葉杜鵑根系侵染情況。結(jié)果為:在春夏兩季桃葉杜鵑根系均完全形成菌根,侵染率為100%(見(jiàn)表2);其它侵染指標(biāo)兩地春季幾乎全部高于夏季(見(jiàn)圖2)。從圖中可以看出,LCG菌根春夏兩季侵染變化小,而B(niǎo)L菌根春夏兩季侵染則變化較大。經(jīng)方差分析,菌根侵染率受季節(jié)影響差異不顯著(P<0.05)。
2.3.3 成年開(kāi)花植株與幼樹(shù)菌根侵染比較
春季采集于百里杜鵑方家坪成年開(kāi)花植株菌根(BL-T1)與幼樹(shù)(BL-T3)菌根侵染情況測(cè)定。結(jié)果表明,不論植株高低,野生桃葉杜鵑根系均完全形成菌根,侵染率為100%(見(jiàn)表2)。其它指標(biāo)BL-T3明顯高于BL-T1菌根,BL-T3根系及侵染根系的侵染強(qiáng)度、菌絲結(jié)豐度指標(biāo)分別比BL-T1高出43.64%、29.47%、53.3% (見(jiàn)圖3)。經(jīng)方差分析,幼樹(shù)與成熟開(kāi)花樹(shù)菌根侵染率差異顯著(P<0.05)。

圖3 成年開(kāi)花樹(shù)與幼樹(shù)菌根侵染比較Fig.3 Mycorrhizal colonization with different standard
2.3.4 播種苗根系菌根侵染情況
1a、2a播種苗的侵染率相對(duì)較低,且差異不顯著(見(jiàn)表2)。從圖4中可能看出,1a、2a苗總侵染強(qiáng)度、菌根菌絲結(jié)豐度指標(biāo)很低,但侵染根段的菌絲結(jié)豐度相對(duì)其它指標(biāo)高很多,這說(shuō)明桃葉杜鵑菌根浸染首先形成ERM的潛力很大。2a苗侵染指標(biāo)較1a呈上升趨勢(shì),侵染根段菌絲結(jié)豐度已近50%。

圖4 1a、2a播種苗侵染比較Fig.4 Seedlings colonization of 1a, 2a
3 討 論
3.1 桃葉杜鵑菌根結(jié)構(gòu)
杜鵑花類(lèi)菌根是一類(lèi)復(fù)雜的菌根[8],一株植株根系常會(huì)結(jié)合多種類(lèi)型菌根菌[19]。吳重華等[20]在冷杉林下的金背杜鵑(Rh. clementinae subsp.aureodorsale)根系中發(fā)現(xiàn)了3種與其形成外生菌根的真菌。Vohník和Albrechtová(2011年)對(duì)歐洲的六種杜鵑花菌根進(jìn)行研究,發(fā)現(xiàn)它們的根內(nèi)同時(shí)存在有ERM真菌、深色有隔的內(nèi)生真菌和其它真菌[21]。Setaro等[22]對(duì)生長(zhǎng)于厄瓜多爾南部熱帶高山雨林的杜鵑花科植物Cavendishia nobilis var. capitata Sebacinales菌根進(jìn)行了研究,指出其菌根為特殊的內(nèi)外生菌根,其菌根中的蠟殼耳目(Sebacinales)是一類(lèi)既能形成菌絲鞘、同時(shí)在細(xì)胞間穿越和侵入細(xì)胞內(nèi)形成菌絲結(jié)結(jié)構(gòu)的真菌。通過(guò)染色觀察,發(fā)現(xiàn)桃葉杜鵑菌根具有ERM特有的、典型的菌絲結(jié),屬于ERM菌根。但在桃葉杜鵑菌根(主要為L(zhǎng)CG-T菌根)中還出現(xiàn)了叢枝菌根(AM)及外生菌根(ECM)具有的一些結(jié)構(gòu)特點(diǎn),這與賈銳等觀察興安杜鵑菌根的結(jié)果相似[23],但形成這些菌根結(jié)構(gòu)是哪類(lèi)或哪幾類(lèi)真菌,還需要進(jìn)一步研究。
3.2 桃葉杜鵑復(fù)雜菌根結(jié)構(gòu)形成的原因
鄭鈺等[7]認(rèn)為杜鵑花植物菌根形成外生菌根是因?yàn)槌L幱诹窒聦拥亩霹N花類(lèi)植物,通過(guò)與外生菌根植物共生的同一種真菌菌絲體整合到森林的公共菌絲體網(wǎng)絡(luò),成為森林內(nèi)部以公共菌絲體網(wǎng)絡(luò)為中介的資源再分配活動(dòng)的參與者,作者指出這可能對(duì)于它們適應(yīng)多樣的生境具有重要意義。鹿沖關(guān)森林公園為桃葉杜鵑模式產(chǎn)地,生物種類(lèi)繁多,樹(shù)林郁閉較高,桃葉杜鵑生長(zhǎng)于林下為中下層灌木。桃葉杜鵑復(fù)雜的菌根結(jié)構(gòu)由多種真菌侵入引起,可能是維持該物種在復(fù)雜環(huán)境中生存的一種方式。
有學(xué)者認(rèn)為杜鵑花菌根內(nèi)同時(shí)存在ERM和其它內(nèi)生菌根的原因是因?yàn)榉植嫉木暥扰c海拔不同[21]。BL-T分布海拔較高,各年齡階段桃葉杜鵑菌根內(nèi)多見(jiàn)菌絲結(jié)結(jié)構(gòu);LCG-T分布海拔較低,菌根真菌為復(fù)雜的菌根結(jié)構(gòu)。從試驗(yàn)結(jié)果來(lái)看,兩者之間是有一定差異,但造成分布地菌根結(jié)構(gòu)的差異原因是否因緯度與海拔不同造成,還需要進(jìn)一步研究。
3.3 桃葉杜鵑菌根真菌的侵染
ERM真菌的侵染受到地域環(huán)境的綜合影響[1],菌根多樣性與地上部分生物的多樣性有相關(guān)性[24]。Grime等實(shí)驗(yàn)發(fā)現(xiàn)由于菌絲網(wǎng)絡(luò)作用,人工小群落中的亞優(yōu)勢(shì)種能夠在優(yōu)勢(shì)種的存在下得以生存,接種后的植物群落比不接種的對(duì)照組具有更高的植物多樣性[25]。分布于鹿沖關(guān)森林公園的桃葉杜鵑菌根侵染率高,侵染強(qiáng)度大,原因可能與地上部分植被豐富有關(guān),機(jī)理可能是通過(guò)森林菌根網(wǎng)絡(luò)在同種或不同種植物間實(shí)現(xiàn)資源的重新分配和共享[26]。分布于百里杜鵑的桃葉杜鵑菌根侵染率及侵染強(qiáng)度稍低可能與其分布區(qū)幾乎為純林有關(guān)。通過(guò)對(duì)幼樹(shù)菌根觀察,發(fā)現(xiàn)菌根幾乎為菌絲體占據(jù),這與金背杜鵑純林菌根侵染率很低的結(jié)果有所不同[20]。
幼樹(shù)的侵染指標(biāo)較成年樹(shù)高,原因除了菌根能顯著提高根系吸收營(yíng)養(yǎng)的能力供其生長(zhǎng),還可能是方便幼樹(shù)在更大范圍內(nèi)選擇適合共生真菌[27]。
菌根在不同季節(jié)侵染率及侵染強(qiáng)度不同,生態(tài)環(huán)境條件是主要原因之一。如在西澳大利亞州的南部,由于季節(jié)性地中海氣候,杜鵑花菌根真菌明顯活動(dòng)僅限于較冷濕的月份,在冬天到春天的一段時(shí)間菌根的侵染達(dá)到了最高[1,28]。桃葉杜鵑菌根侵染是否受到貴州亞熱帶高原季風(fēng)濕潤(rùn)氣候的影響需要作進(jìn)一步的研究。百里杜鵑的桃葉杜鵑菌根春夏季侵染變化較大的原因,可能與生長(zhǎng)在高海拔、晝夜溫差大、長(zhǎng)時(shí)間處于空氣與土壤濕度均較大的環(huán)境中有關(guān)系。影響侵染率及侵染強(qiáng)度的另外一個(gè)原因是植物自身生長(zhǎng)發(fā)育過(guò)程影響。桃葉杜鵑在春季開(kāi)花,集中花期在5月,開(kāi)花期需要更多的菌根來(lái)吸收營(yíng)養(yǎng)物質(zhì),特別是對(duì)磷元素的獲取[29]。桃葉杜鵑春季菌根侵染普遍高于夏季的情況及原因與梅(Prunus mume Sieb. et Zucc.)和蘭(Orchis L. )類(lèi)似[30-31]。
1a、2a播種苗培養(yǎng)基質(zhì)中帶有部分原生境真菌,因此播種苗具有一定的侵染,雖然侵染率相對(duì)野生較低,但說(shuō)明桃葉杜鵑人工栽培條件下能形成ERM,這為桃葉杜鵑菌根化育苗提供了事實(shí)依據(jù)。2a苗侵染指標(biāo)較1a呈上升趨勢(shì),這可能與ERM真菌選擇、侵染時(shí)間和定居時(shí)間有關(guān),然而是否與小苗的生長(zhǎng)適應(yīng)性有關(guān)需進(jìn)一步研究。
從試驗(yàn)結(jié)果來(lái)看,幼苗—幼樹(shù)—成年菌根侵染指標(biāo)呈現(xiàn)低—高—低的變化趨勢(shì),變化產(chǎn)生的原因及對(duì)宿主的影響還需要更多的探索。
致謝:貴州大學(xué)生命科學(xué)學(xué)院江龍教授,林學(xué)院謝雙喜教授;1a、2a桃葉杜鵑小苗由百里杜鵑林業(yè)局黃家涌提供。
[1] Dixon K W, Sivasithamparam K, Read D J. Ericoid Mycorrhizas in Plant Communities[J].Microorganisms in Plant Conservation and Biodiversity, 2004,227-239.
[2] Read D J.The structure and function of the ericoid mycorrhizal root[J].Annals of Botany,1996,77:365-374.
[3] Cairney J W G.Evolution of mycorrhiza systems[J].Naturwissens chaften,2000,87:467-475.
[4] Cairney J W G,Meharh A A. Ericoid mycorrhizal:A partnership that exploits harsh edaphic conditions[J].European Journal of Soil Science,2003,54:735-740.
[5] Sokolovski S G, Meharg Y A, Maathuis F J M. Calluna vulgaris root cells show increased capacity for amino acid uptake when colonized with the mycorrhizal fungus Hymenoscyphus ericae[J].New Phytologist.2002,155(3):525-530.
[6] Starrett M C,Blazich F A,Shafer S R, et al. In vitro colonization of micropropagated Pieris floribunda by ericoid mycorrhizae.Establishment of mycorrhizae on microshoots[J].Hort Science,2001,36(2):353-356.
[7] Strandberg M,Johannsson M.Uptake of nutrients in Calluna vulgaris seed plants growth with and without mycorrhizae[J].Forest Ecology and Management, 1999,114:129-135.
[8] 張春英,侯奕敏,戴思蘭.云錦杜鵑菌根顯微結(jié)構(gòu)觀察[J].園藝學(xué)報(bào),2008,35(11):1641-1646.
[9] 鄭 鈺,高 博,孫立夫,等.銀葉杜鵑和繁花杜鵑根部真菌的多樣性[J].生物多樣性,2010,18(1):76-82.
[10] Ashford A E,Allaway W G, Reed M L. A possible role of Lysinema ciliatum R. Br.&other Epacridaceae[J].Annals of Botany,1996,77:375-381.
[11] Massicotte H B,Melville L H,Peterson R L.Structural characteristics of root-fungal interactions for five ericaceous species in eastern Canada [J].Canadian Journal of Botany,2005,83:1057-1064.
[12] Usuki F, Junichi A P, Kakishima M.Diversity of ericoid mycorrhizl fungi isolated from hair roots of Rhododendron obtusum var. Kaempferi in a Japanese red pine forest[J].Mycoscience,2003,44:97–102.
[13] 徐小蓉,張習(xí)敏,牛曉娟,等.赤霉素+2,4-D及赤霉素+丁酰肼對(duì)馬纓杜鵑光合作用日變化的影響[J].中南林業(yè)科技大學(xué)學(xué)報(bào),2011,31(11):131-136.
[14] 李朝嬋,趙云龍,張冬林,等.長(zhǎng)蕊杜鵑扦插內(nèi)源激素變化及解剖結(jié)構(gòu)觀察[J].林業(yè)科學(xué)研究,2012,25(3):360-365.
[15] 高貴龍,龍秀琴,胡小京,等.赤霉素對(duì)兩種高山杜鵑種子發(fā)芽的影響[J].種子,2010,29(5):22-25
[16] 苗永美,王永清,莊 平,等.桃葉杜鵑組織培養(yǎng)技術(shù)研究[J].生物學(xué)雜志,2006,23(6):29-31.
[17] Komanik P P,Bryon W C,Schultz R C. Procedures and equipment for staining large numbers of plant root samples for endomycorrhizal assay[J].Canadian Journal of Microbiology,1980,26(4):536-538.
[18] Trouvelot A, Kough J L, Gianinazzi-Pearson V. Mesure du taux de mycorhization VA d′un système radiculaire.Recherche de méthodes d′estimation ayantune signification fonctionnelle.In:Physiological and Genetical Aspects of Mycorrhizoe,V.Gianinazzi-Pearsin and S.Gianinazzi(eds.).INRA Press, Paris,1986,217-221.
[19] Allen T R,Miller T,Berch S M, et al. Culturing and direct DNA extraction find different fungi from the same ericoid mycorrhizal roots[J].New Phytologist, 2003,160:255-272.
[20] 吳重華,王吉忍,楊俊秀,等.太白山自然保護(hù)區(qū)金背杜鵑菌根調(diào)查研究[J].西北林學(xué)院學(xué)報(bào),2000,15(3):68-70.
[21] Vohník M and Albrechtová J.The Co-occurrence and Septate Endophytes in Roots of Six European Rhododendron Species,Folia Geobotanica, 2011, 15 March. published online(springer).
[22] Setaro S, Wei? M, Oberwinkler F, et al. Sebacinales form ectendomycorrhizas with Cavendishia nobilis, a member of the Andean clade of Ericaceae, in the mountain rain forest of southern Ecuador[J]. New Phytologist,2006,169: 355–365.
[23] 賈銳,楊秀麗,閆 偉.興安杜鵑菌根形態(tài)特征和土壤理化性質(zhì)的關(guān)系研究[J].內(nèi)蒙古農(nóng)業(yè)大學(xué)學(xué)報(bào),2011,32(3):63-66.
[24] Peterson T A, Mueller W C, Englander L.Anatomy and ultrastructure of a Rhododendron root fungus association[J].Canadian Journal of Botany,1980,58:2421-2433.
[25] Grime J P,Mackey J M L,Hiller S H,et al. Floristic diversity in a model system using experimental microcosms[J].Nature,1987,328:420-422.
[26] 梁 宇,郭良棟,馬克平.菌根真菌在生態(tài)系統(tǒng)中的作用[J].植物生態(tài)學(xué)報(bào),2002,26(6):739-745
[27] Zettler L W,Pixkin K A,Stewart S I, et al.Protocom mycobionts of the federally threatened eastein prairie fringed orchid,Platanthera leucophaea(Nutt.)Lindley and a technique to prompt leaf elongation in seedlings[J]. Studies in Mycology, 2005,53:163-171.
[28] Kemp E, Adam P,Ashford A E.Seasonal changes in hair roots and mycorrhizal colonization in Woollsia pungens (Cav.) F. Muell.(Epacridaceae) [J].Plant and Soil,2003,250: 241-248.
[29] Dighton J, Coleman D C. Phosphorus relations of roots and mycorrhizas of Rhododendron maximum L. in the southern Appalachians North Carolina[J]. Mycorrhiza,1992,1:175-184.
[30] 蔡邦平,陳俊愉,張啟翔,等.中國(guó)梅叢枝菌根侵染的調(diào)查研究[J].園藝學(xué)報(bào),2008,35(4):599-602.
[31] 侯天文,金 輝,劉紅霞,等.四川黃龍溝優(yōu)勢(shì)蘭科植物菌根真菌多樣性及其季節(jié)變化[J].生態(tài)學(xué)報(bào),2010,30(13):3424-3432.
Study on microstructure and infections of Rhododendron annae mycorrhiza
OU Jing1, LIU Ren-yang1, CHEN Xun2
(1.Forestry College , Guizhou University, Guiyang 550025, Guizhou, China; 2.Guizhou Provincial Dept. of Science and Technology,Guiyang 550001, Guizhou ,China)
By using the acid fuchsin staining, the roots of Rhododendron annae Franch were dyed and squashed, which grows in Guizhou province, and the microstructure and mycorrhizal colonization were observed. The results show that the lateral roots of ferine R. annae have complex structure, were enwrap by dense hypase to form hyphal sheath on the outside of the roots and there was no root hair; There were intracellular hyphal knots which are the unique and typical structure of ericoid mycorrhiza, there also were a few septate hypha and fasciculate hyphae structure. Wild plant infection index was higher, the indexes differences of the infected roots in different regions were remarkable, the indexes of Luchonghuang mycorrhiza of adult flowering plant were higher than Fangjiaping’s; In different seasons, the rates in spring were higher than that in summer; The indexes of flowering adult tree were obviously less than that young tree. Sowing seedlings’ index was lower. The infection indexes of 2-year-old sowing seedlings showed a rising trend compared with that of 1-year-old.
Rhododendron annae mycorrhiza;ericoid mycorrhiza;microstructure;infection index
S718.8
A
1673-923X(2012)11-0028-06
2012-10-10
貴州省農(nóng)業(yè)科技攻關(guān)項(xiàng)目(黔科合NY字[2011]3076號(hào))
歐 靜(1974-),女,貴州威寧人,副教授,主要從事森林培育與種苗培育研究工作, E-mail:coloroj@126.com
陳 訓(xùn)(1956-),男,貴州赫章人,研究員,博導(dǎo),主要從事森林培育與種苗培育研究工作;E-mail:chenxunke1956@163.com
[本文編校:吳 毅]
