干旱對4種幼苗氣體交換參數的影響
2012-01-04 09:25:15楊振意郭淑紅
中南林業(yè)科技大學學報 2012年5期
關鍵詞:植物
楊振意 , 薛 立 , 郭淑紅 , 張 柔
(華南農業(yè)大學 林學院, 廣東 廣州 510642)
干旱對4種幼苗氣體交換參數的影響
楊振意 , 薛 立 , 郭淑紅 , 張 柔
(華南農業(yè)大學 林學院, 廣東 廣州 510642)
對蒲桃Syzygium jambos、南五味子Kadsura longipedunculata、海南紅豆Ormosia pinnata和灰木蓮Mangliet iaglauca幼苗在不同干旱強度下的葉片氣體交換參數進行了研究。結果表明,隨著干旱脅迫時間的延長,4種幼苗的凈光合速率(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)顯著下降;蒲桃的胞間СO2濃度(Ci)顯著降低,氣孔限制值(Ls)下降后上升,南五味子和海南紅豆的Ci都為下降后上升,Ls呈現上升后下降;灰木蓮的Ci持續(xù)上升,而Ls持續(xù)下降。蒲桃的葉片水分利用率(WUE)上升,而其他幼苗則不斷降低;主成分分析表明,幼苗耐干旱性順序為蒲桃>南五味子>海南紅豆>灰木蓮。
干旱脅迫;幼苗;氣體交換參數;主成分分析
光合作用是植物各項生命活動的基礎,直接影響植物的生長和發(fā)育,其大小不僅與自身遺傳特性有關, 而且受眾多環(huán)境因子的影響[1]。干旱脅迫是樹木逆境最普遍的形式[2],水是對光合作用有重要影響的環(huán)境因子之一,干旱條件下植物葉片的光合速率下降,生長受抑制。由于森林多生長在山地,少雨季節(jié)往往出現干旱脅迫,所以干旱脅迫下樹木的光合作用一直是人們研究的熱點。例如,Rouhi等[3]對干旱脅迫下的杏樹Prunus armeniaca、Ogaya等[4]對冬青櫟Quercus ilex和紅景天Phillyrea latifolai、Ranjbarfardooei 等[5]對埃及黃連木Pistacia khinjuk和鈍黃連木P. mutica光合速率等指標進行了研究;劉慧民等[6]研究了干旱脅迫的華北繡線菊和金山繡線菊光合能力,夏江寶等[7]報道了干旱脅迫的常春藤凈光合速率日變化及其影響因子;羅明華[8]等研究了干旱脅迫對丹參葉片氣體交換參數的影響。蒲桃Syzygium jambos葉密濃綠,樹姿優(yōu)美,花果期長,在華南地區(qū)廣泛種植。南五味子Kadsura longipedunculata是具有較高的藥用和食用價值的常綠或半常綠攀援木質藤本[9,10], 葉、花、果均可供觀賞[10],具備優(yōu)良的觀賞特性和生態(tài)學特性[11]。海南紅豆Ormosia pinnata是原產我國廣東和海南島以及越南等國的固氮喬木樹種[12],主干通直, 萌芽更新力強, 耐土壤瘠薄, 冠形優(yōu)雅,且抗風、防火、抗環(huán)境污染, 是優(yōu)良的多用途樹種[13]。灰木蓮Mangliet iaglauca樹形美觀, 枝葉繁茂, 蔭濃, 是優(yōu)良的庭園及行道綠化樹種。目前還未見干旱脅迫對這四種樹種光合特性影響的報道。作者對這些內容進行研究,探討其對不同水分狀況的適應機制, 可以為山地造林時的樹種選擇提供參考。
1 材料與方法
1.1 材 料
2011年4月選擇廣東省林木種苗示范基地的1年生實生苗容器苗,放置華南農業(yè)大學林學院溫室內進行研究。幼苗生長基本情況見表1。
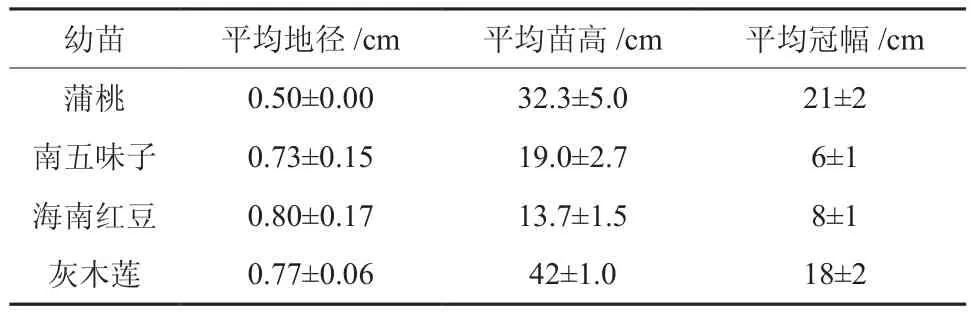
表1 4種幼苗的基本情況(平均值±標準差)Table 1 General characteristics of four seedlings(mean±SD)
1.2 方 法
采用不澆水自然干旱方式進行脅迫試驗。在脅迫處理的0(對照)、2、4和6 d,進行光合氣體交換參數和土壤容積含水量的測定。光合氣體交換參數用美國 L?-СOR 公司生產的 L?-6400便捷式光合測定儀測定。測定參數主要包括:葉片凈光合速率 (Pn,μmol?m-2·s-1)、氣孔導度 (Gs,mmol?m-2·s-1)、 胞 間 СO2濃 度 (Ci,μmol?mol-1)和蒸騰速率 (Tr,mmol?m-2?s-1)。每株植物選擇幼苗頂部開始的第3~8枚葉中的三枚成熟葉進行掛牌標記,測定時間為光合作用比較活躍的9:00~11:00,每葉片記錄3個值,取平均值。測定時用紅藍光源葉室 , 光強 1000 μmol?m-2s-1, 溫度為室溫,空氣流量為500 Lmol?s-1,СO2濃度為空氣中СO2濃度。利用美國產M?2X型土壤水分探頭監(jiān)測土壤容積含水量(RSVWС)的變化。計算水分利用效率RWUE和氣孔限制值Ls計算公式分別見式1和式2:

式(2)中:Ca為大氣СO2濃度。
為綜合評價不同樹種的抗旱性,采用SAS8.1軟件系統(tǒng)對各項指標測定結果進行主成分分析,以確定4個樹種的抗旱性[14]。
2 結果與分析
2.1 干旱脅迫中的土壤水分和幼苗形態(tài)變化
第0 、2、4和6d的土壤體積含水量分別為28%、14%、6%和3%(見圖1)。在第2天時,蒲桃、南五味子、海南紅豆和灰木蓮形態(tài)無明顯變化;第4 天時,蒲桃形態(tài)無明顯變化,南五味子近根部分葉子成棕黃色,大部分葉子變皺并變軟,海南紅豆的小葉稍微閉合,灰木蓮葉子開始萎焉變皺;第6天時,蒲桃葉片變化不大,葉片下垂,南五味子大部分葉子干枯蜷曲,海南紅豆部分小葉基本閉合,有老葉脫落,灰木蓮大部分葉片萎焉卷曲。
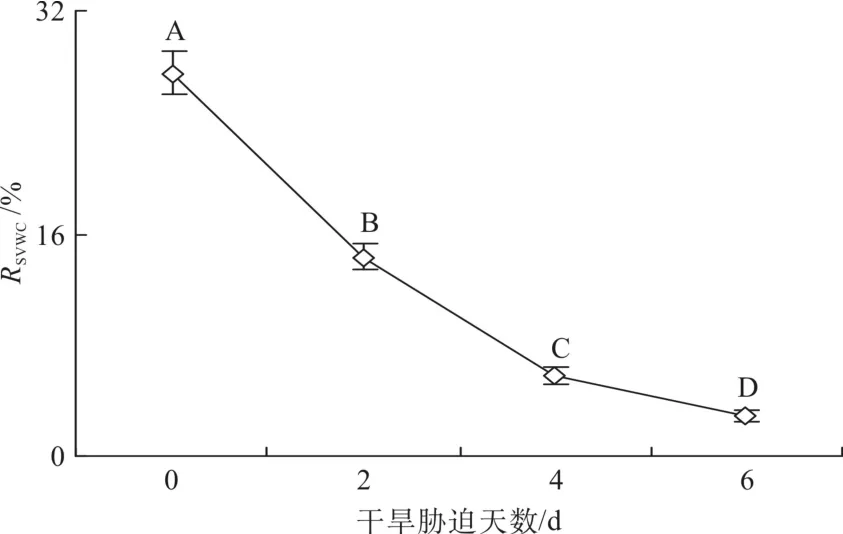
圖1 干旱脅迫中的土壤體積含水量的變化Fig. 1 Changes of soil volumetric water content with time during drought stress
2.2 干旱脅迫中凈光合速率的變化
由圖2可知,第0天時蒲桃和南五味子的凈光合速率(Pn)較高,海南紅豆和灰木蓮的Pn較低。隨著干旱天數的增加,4種幼苗的Pn都不斷降低,在第2d、4d和6d時與對照相比,蒲桃的Pn分別下降6%、85%和90%,南五味子分別下降7%、96%和104%,海南紅豆分別下降6%、61%和90%,灰木蓮分別下降29%、82%和90%。干旱脅迫過程中,蒲桃、南五味子和海南紅豆的Pn在第4天開始下顯著降低(P<0.001),灰木蓮的Pn在第2天就開始顯著降低(P<0.001)。在第6天時蒲桃的Pn相對最高,南五味子降為負值,海南紅豆和灰木蓮總降幅相差不大。
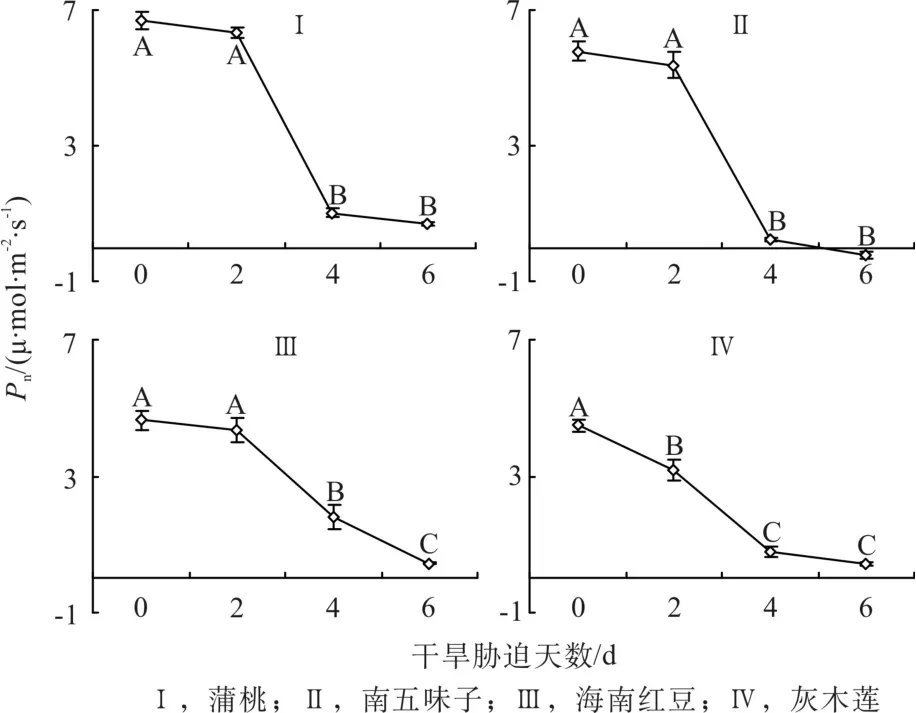
圖2 干旱脅迫中凈光合速率的變化Fig. 2 Changes of net photosynthetic rate during drought stress
2.3 干旱脅迫中氣孔導度的變化
由圖3可知,第0天時,蒲桃的氣孔導度(Gs)最高,南五味子和灰木蓮的Gs相近,海南紅豆的Gs相對最低。隨著干旱天數的增加,4種幼苗的Gs呈現與凈光和速率相似的變化趨勢,在第2天、4天和6天與對照相比,蒲桃的Gs分別下降8 %、88%和95%,南五味子的Gs分別下降25%、91%和92%,海南紅豆的Gs分別下降11%、55%和84%,灰木蓮的Gs分別下降32 %、75%和76%。脅迫過程中,蒲桃和海南紅豆的Gs在第4天開始顯著降低(P<0.001),南五味子和灰木蓮在第2天就開始顯著降低(P<0.001)。蒲桃和南五味子的Gs的總降幅相對較大,海南紅豆次之,灰木蓮最小。
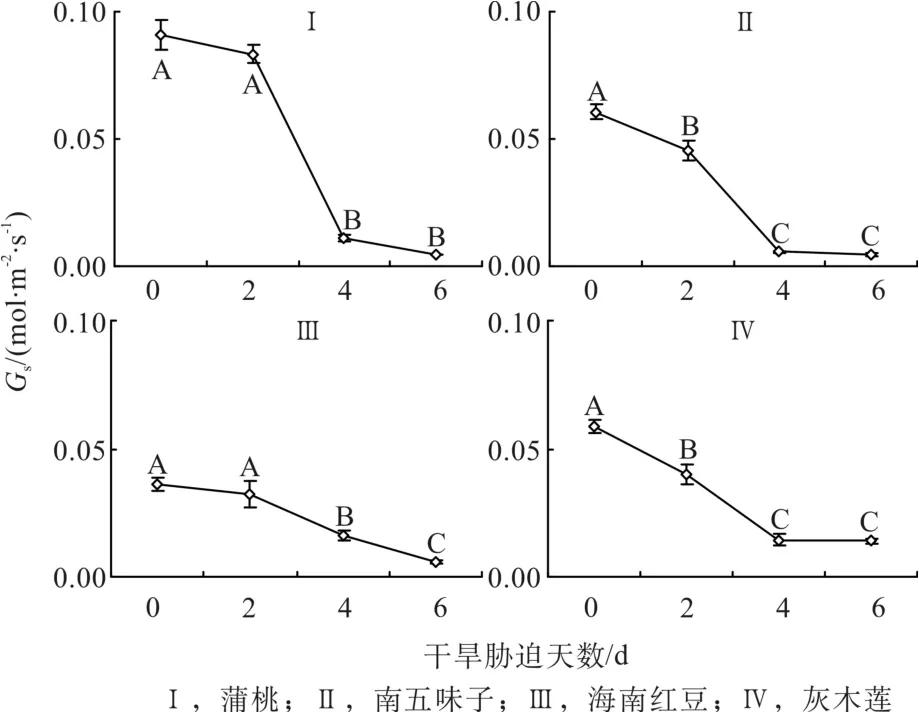
圖3 干旱脅迫中氣孔導度的變化Fig. 3 Changes of stomatal conductance during drought stress
2.4 干旱脅迫中胞間СO2濃度的變化
由圖4可知,在第0天時,蒲桃、南五味子和灰木蓮的蒲桃的胞間СO2濃度(Ci)相差不大。隨著干旱天數的增加,4種幼苗的Ci變化各異。脅迫過程中,蒲桃的Ci一直下降,在第2天、4天和6天 的Ci分別比對照相下降2%、4%和35%,南五味子的Ci分別為降低29%,升高36%和60%;海南紅豆的Ci分別為降低11%,升高43%和56%;灰木蓮的Ci分別為升高3%、18%和25%。南五味子和海南紅豆Ci先降低后升高,灰木蓮的Ci持續(xù)升高。
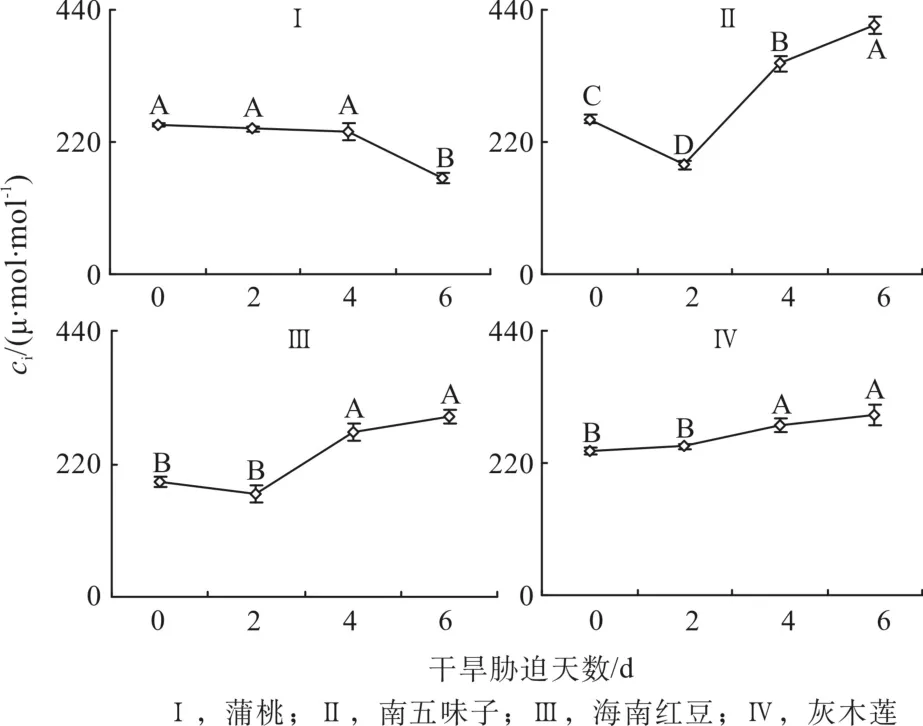
圖4 干旱脅迫中胞間СO2濃度的變化Fig. 4 Changes of intercellular carbon dioxide concentration during drought stress
2.5 干旱脅迫中蒸騰速率的變化
由圖5可知,第0天時,蒲桃的蒸騰速率(Tr)最高,南五味子和灰木蓮的Tr相差近,海南紅豆的Tr相對最低。隨著干旱天數的增加,4種幼苗的Tr變化與氣孔導度(Gs)的變化一致。與對照相比,蒲桃第2、4和6天的Tr分別下降5%、83%和93%,南五味子分別下降15%、88%和90%, 海南紅豆分別下降10%、43%和78%,灰木蓮分別下降27%、69%和70%。脅迫過程中,蒲桃和海南紅豆的Tr在第4天顯著降低(P<0.001),南五味子和灰木蓮在第2天就開始顯著降低(P<0.001)。蒲桃和南五味子的Tr總降幅相對最高,海南紅豆與灰木蓮的Tr總降幅相對較低。最終,蒲桃、南五味子和海南紅豆的Tr較低,灰木蓮的Tr最高。
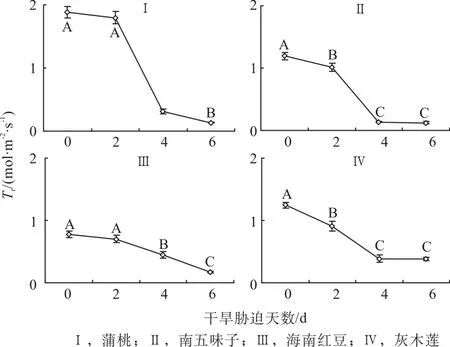
圖5 干旱脅迫中蒸騰速率的變化Fig. 5 Changes of transpiration rate during drought stress
2.6 干旱脅迫中氣孔限制值的變化
由圖6可知,第0天時,蒲桃、南五味子和灰木蓮的氣孔限制值(Ls)相差不大且相對較低,海南紅豆的Ls相對較高。隨著干旱天數的增加,4種幼苗的Ls表現出與胞間СO2濃度(Ci)相反的趨勢。與對照相比,蒲桃第2、4和6d的Ls分別下降2%、上升12%和89%;南五味子分別升高41%,下降48%和94%;海南紅豆分別升高5%,下降31%和46%;灰木蓮分別下降2%、24%和38%。干旱過程中,蒲桃的Ls趨勢為上升,南五味子和海南紅豆Ls先下降后上升,灰木蓮的Ls持續(xù)下降。
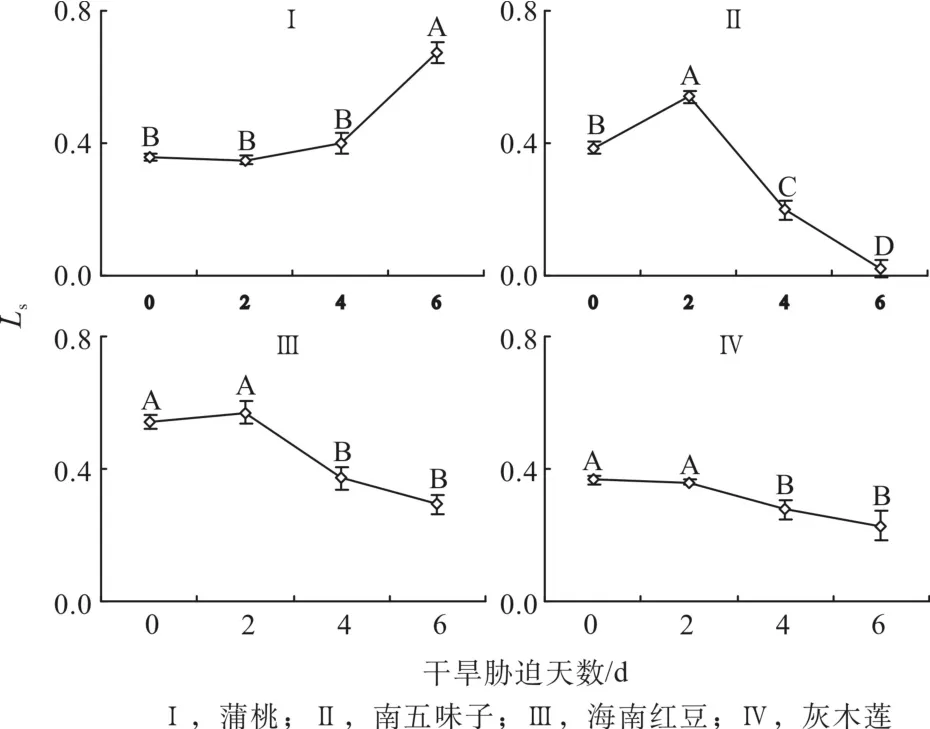
圖6 干旱脅迫中氣孔限制值的變化Fig. 6 Changes of stomatal limit during drought stress
2.7 干旱脅迫中葉片水分利用率的變化
由圖7可知,第0天時,海南紅豆的葉片水分利用率(RWUE)較高,南五味子、灰木蓮和蒲桃相近。隨著干旱天數的增加,4種幼苗的WUE變化不一。干旱脅迫第2天、4天和6 天與對照相比蒲桃的WUE相比分別為上升5%、3%和43%;南五味子的WUE分別下降7%、64%和100%,海南紅豆的WUE分別降低13%、46%和50%,灰木蓮的WUE分別降低7%、41%和53%。
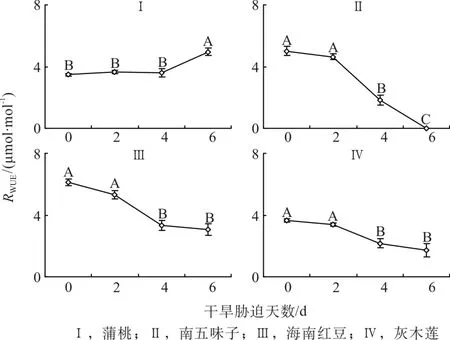
圖7 干旱脅迫中水葉片分利用率的變化Fig. 7 Changes of water use efficiency during drought stress
2.8 4種幼苗抗旱能力的綜合評價
干旱脅迫下不同植物氣體交換的差異體現了植物對干旱環(huán)境的適應能力[7],利用主成分對幼苗生理指標進行分析可以得出其抗旱能力[14]。由主成分的特征向量可計算出4種幼苗的主成分得分順序為蒲桃、南五味子、海南紅豆、灰木蓮,蒲桃的主成分得分遠遠大于其他3種幼苗,表明其抗旱能力強。
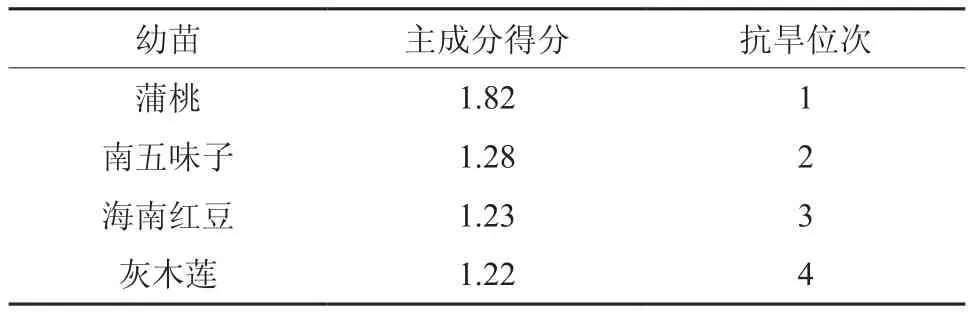
表1 干旱脅迫下4種幼苗的主成分得分Table 1 Score of principal components of 4 seedlings under drought stress
3 討 論
本研究中,蒲桃、南五味子、海南紅豆和灰木蓮的凈光合速率(Pn)、氣孔導度(Gs)和蒸騰速率(Tr)隨著干旱脅迫的加強都不斷下降。植物受到干旱脅迫時,葉片水分散失和葉片水勢下降,導致氣孔開度減小,氣孔阻力增加, 最后引起氣孔關閉。氣孔是光合作用時吸收空氣中СO2的和蒸騰過程中水蒸氣從體內排到體外的主要通道,干旱脅迫造成氣孔阻力增加, СO2進入葉片受阻,同時光合產物輸出變慢,葉片中淀粉水解加強,糖類積累,結果會引起光合速率下降[15,16]。植物干旱脅迫時通過關閉氣孔而減少蒸騰失水,引起蒸騰速率下降,有利于推遲水分虧缺發(fā)展到有害或致死程度的時間,但是因為限制了СO2的進入而減弱光合作用。在干旱脅迫下,光合特性較早出現大幅度降低的幼苗,干旱適應性可能較差。本研究中,蒲桃的凈光合速率(Pn)、氣孔導度(Gs)和蒸騰速率(Tr)均在第4天顯著下降,其他幼苗的這3種生理指標中,有的在第2天就顯著降低,說明抗旱性強。
蒲桃的胞間СO2濃度(Ci)隨著處理天數的增加持續(xù)降低,而氣孔限制值(Ls)下降后升高,葉片形態(tài)未出現明顯變化,說明氣孔限制引起光合作用下降,但是葉片沒有受損。南五味子和海南紅豆的Ci均為下降后上升 ,Ls均呈現上升后下降;灰木蓮的Ci為持續(xù)上升,Ls為持續(xù)下降。依據胞間СO2濃度(Ci)和氣孔限制值(Ls)的變化方向[7]可以判斷是氣孔和非氣孔限制因素[9]。氣孔限制理論用胞間СO2濃度(Ci)和氣孔限制值(Ls)的變化作為判別植物是否受氣孔限制標準,若Ci降低和Ls增大, 則以氣孔限制為主, 若Ci增大和Ls減小, 則以非氣孔限制為主[7]。本研究中,限制蒲桃光合作用的主要是氣孔限制,而海南紅豆和南五味子在干旱2 d內 Ci降低和Ls增大,限制其光合作用的主要因素是氣孔限制,因為輕度干旱脅迫造成根系導水能力下降而使氣孔阻力增加,從而進一步限制СO2吸收,細胞間隙СO2濃度(Ci)下降和氣孔限制值增加。此時光合能力的降低是短暫的可恢復性的, 對植物生理傷害較小[7]。海南紅豆和南五味子干旱后2 d后,以及灰木蓮在整個實驗期間,Ci增大而Ls減小,說明Pn降低主要由氣孔限制轉向非氣孔限制,幼苗木水分失調所引起的其他生理機能紊亂與反應機構的損傷使其物質代謝受阻[15],如葉綠體的電子傳遞速率降低且與光合磷酸化解偶聯(lián),影響同化力的形成,葉綠體變形,片層結構被破壞, 1, 5-二磷酸核酮糖(RuBP)的再生受到阻礙等[17]。海南紅豆、南五味子和灰木蓮的Pn大幅下降,葉肉細胞已經發(fā)生了不能逆轉的變化,南五味子葉片在形態(tài)上已經表現出了巨大的變化,葉片干枯卷曲,光合作用基本停止,而海南紅豆小葉葉片閉合,老葉開始脫落,灰木蓮大部分葉片萎焉卷曲,這與前人對其他植物的研究結論基本一致[7,18-20]。
水分利用效率作為植物氣體交換的一個重要指標,表征了植物在等量水分消耗情況下,固定СO2的能力,它不僅體現了植物自身光合能力大小,也能反映植物有效利用水分的能力[21],許多植物在干旱脅迫下通過改變自身的生理生態(tài)特性來達到高效用水的目的[21-23]。隨著干旱天數的增加,蒲桃葉片的水分利用率持續(xù)增加,而南五味子、海南紅豆和灰木蓮的WUE持續(xù)降低,說明前者消耗同樣數量的水分,可以固定較多的СO2,能在干旱脅迫下高效利用水分。
植物的抗旱性是一個復雜的綜合性狀,它受多種因素影響,不僅與植物的本身的遺傳基因有關,而且還與植物的生長環(huán)境有關, 而各因素的重要性也不同,單一指標常常難以準確地評價某一樹種的抗旱能力。馮慧芳等[14]研究4種苗木的抗旱生理指標和何躍君等[24]研究6種苗木的抗寒生理指標中,發(fā)現主成分分析的苗木抗逆能力與生理指標變化一致。本研究中,根據主成分分析的得分可知,4種幼苗的主成分分析抗旱得分順序為蒲桃、南五味子、海南紅豆、灰木蓮,與其形態(tài)指標的變化一致,說明主成分分析適用于評價樹木抗旱研究。
[1] 步秀芹,徐學選,杜 峰,等. 黃土丘陵區(qū)翻白草光合特性的研究[J]. 水土保持通報,2007, 27(6): 85-89.
[2] 李 燕, 薛 立, 吳 敏. 樹木抗旱機理研究進展[J]. 生態(tài)學雜志,2007,26(11):1857-1866.
[3] ROUH? V, SAMSON R, LEMEUR R, et al. Photosynthetic gas exchange characteristics in three different almond species during drought stress and subsequent recovery[J]. Environmental and Experimental Botany, 2007, 59: 117-129.
[5] RANJBARFORDOE? A, SAMSON R, VAN DAMME P, et al. Effects of drought stress induced by polyethylene glycol on pigment content and photosynthetic gas exchange of Pistacia khinjuk and P. mutica[J]. Photosynthetica, 2000, 38(3): 443-447.
[6] 劉慧民,車艷雙,車代弟,等. 干旱脅迫對華北繡線菊和金山繡線菊光合能力的影響[J]. 應用生態(tài)學報,2010, 21(8):2004-2009.
[7] 夏江寶,張光燦,許景偉,等. 干旱脅迫下常春藤凈光合速率日變化及其影響因子分析[J]. 水土保持通報,2010, 30(3):78-82.
[8] 羅明華,胡進耀,吳慶貴,等. 干旱脅迫對丹參葉片氣體交換和葉綠素熒光參數的影響[J]. 應用生態(tài)學報,2010, 21(3):619-623.
[9] 藍盛芳. 中國南五味子屬分類研究[J]. 華南農學院學報,1984, 5(2): 83-92.
[10] 鄧白羅,謝碧霞,程 張. 中國南五味子屬植物的種質資源及開發(fā)利用[J]. 中南林業(yè)科技大學學報,2008, 28(6): 90-94.
[11] 鄧白羅,謝碧霞,劉 暉,等. 南五味子的地理分布與園林應用[J]. 中南林業(yè)科技大學學報,2009, 29(5): 184-186.
[12] 趙 平,曾小平,彭少麟,等. 海南紅豆(Ormosia pinnata)夏季葉片氣體交換、氣孔導度和水分利用效率的日變[J]. 熱帶亞熱帶植物學報,2000, 8(1): 35-42.
[13] 粟 娟,孫 冰,王德禎. 海南紅豆(Ormosia pinnata)的生物學特性和觀賞價值[J]. 林業(yè)科學研究,1995, 8(6): 677-681.
[14] 馮慧芳,薛 立,任向榮,等. 4種闊葉苗木對PEG模擬干旱的生理響應[J]. 生態(tài)學報,2011,31(2): 371-382.
[15] YORDANOY ?, TSONEY T, GOLTSEY V, et al. ?nteractive effect of water def i cit and high temperature on photosynthesis in sunf l ower and maize plants.1[J]. Photosynthetica, 1997, 33(3/4):391-402.
[16] 劉志媛,黨選民,曹振木. 土壤水分對黃秋葵苗期生長及光合作用的影響[J]. 熱帶作物學報, 2003, 24(1): 70-73.
[17] FLEXAS J, MEDRANO H. Drought-inhibition of photosynthesis in С3 plant stomatal and non-stomatal limitations revisited[J].Annals of Botany, 2002, 89: 183-189.
[18] 竇新永,吳國江,黃紅英,等. 麻瘋樹幼苗對干旱脅迫的響應[J]. 應用生態(tài)學報, 2008, 19(7): 1425-1430.
[19] 徐興友,王子華,龍 茹,等. 干旱對6種野生花卉光合色素含量與氣體交換的影響[J]. 經濟林研究,2008, 26(4): 1-6.
[20] 楊玉珍,張云霞,彭方仁. 干旱脅迫對不同種源香椿苗木光合特性的影響[J]. 北京林業(yè)大學學報,2011, 33(1): 44-48.
[21] 閆海龍,張希明,許 浩,等. 塔里木沙漠公路防護林3種植物光合特性對干旱脅迫的響應[J]. 生態(tài)學報,2010, 30(10):2519-2528.
[22] GE Y, Lu Y J, L?AO J X. et al. Photosynthetic parameters of Mosla hangchowensis and Mosla dianthera as affected by soil moisture[J]. Photosynthetica, 2004, 42(3): 387-391.
[23] 包 卓,孟祥英,張曉松,等. 干旱脅迫對5 種園林綠化植物生理生化的影響[J]. 經濟林研究,2010, 28(1): 46-50.
[24] 何躍君,薛 立,任向榮,等. 低溫脅迫對6種幼苗生理特性的影響[J]. 生態(tài)學雜志,2008,27(4):524-531.
Effects of drought stress on leaf gas exchange parameters of four seedlings
YANG Zhen-yi, XUE Li, GUO Shu-hong, ZHANG Rou
(Сollege of Forestry, South Сhina Agricultural University, Guangzhou 510642, Guangdong, Сhina)
The effects of drought stress on leaf gas exchange of four seedlings ofSyzygium jambos,Kadsura longipedunculata,Ormosia pinnataandMangliet iaglaucawere studied. The results show that net photosynthetic rate (Pn), stomatal conductance (Gs)and transpiration rate (Tr) of the four seedlings continuously decreased with increasing drought time. ?ntercellular carbon dioxide concentration(Ci) ofS. jambosdecreased, whereas its stomatal limits (Ls) fi rst decreased and then increased, whereas intercellular carbon dioxide concentration(Ci) decreased and then increased, theCiofK. longipedunculata,O. pinnatadecreased followed by an increase; theCiofM. iaglaucaincreased and itsLsdecreased during drought experiment period. Water use eff i ciency (WUE) ofS. jambosincreased and that of other seedlings decreased with increasing drought time. The drought resistance of the four seedlings was evaluated by using principal component analysis, and the order of drought resistance wasS. jambos>K. longipedunculata>O. pinnata>M. iaglauca.
drought stress; seedling; gas exchange parameter; analysis of principal component
S72;Q943
A
1673-923X (2012)05-0067-06
2011-11- 15
廣東省林業(yè)局資助項目(F09054);廣東省林業(yè)科技創(chuàng)新專項資金項目(2008KJСX008-01;2009KJСX011-01)
楊振意(1987-),男,碩士研究生,主要從事森林培育和樹木生理學研究;E-mail:691514703@qq.com
薛 立(1958-),男,湖南桃江人, 教授,博士, 主要研究方向為森林生態(tài)和森林培育;E-mail: forxue@scau.edu.cn
[本文編校:歐陽欽]
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
