乳鼠耳蝸單離外毛細胞胞膜Prestin分布的實驗研究*
2012-01-03 05:24:35尹時華
重慶醫(yī)學 2012年30期
張 博,尹時華
(廣西醫(yī)科大學第一附屬醫(yī)院耳鼻咽喉頭頸外科,南寧 530021)
Prestin是一種特異存在于耳蝸外毛細胞(outer hair cell,OHC)的動力蛋白,該蛋白在解釋哺乳動物耳蝸的高度敏感性和頻率選擇性以及OHC的電動特性機制上具有重要意義。近年來,關于Prestin在OHC的表達、定位以及Prestin與OHC電運動和耳蝸放大效應之間的關系成為研究熱點。早期研究認為采用全細胞電壓鉗技術,在OHC細胞核平面以上的側膜區(qū)域能夠記錄到與電運動相關的非線性電容,在平面以下的側膜區(qū)域則不能記錄到明顯的非線性電容,因此,有人推測OHC細胞膜在細胞核平面以下部位無Prestin分布[1-4]。而近年來有人運用熒光雙染及復合染色等方法觀察發(fā)現(xiàn),Prestin不僅分布于成年大鼠和豚鼠OHC側膜,在細胞底部也有分布[5]。但有關Prestin在乳鼠耳蝸單離OHC細胞膜上的分布尚未見報道。本研究以Wistar乳鼠為研究對象,觀察Prestin在乳鼠耳蝸單離OHC細胞膜上的分布定位。
1 材料與方法
實驗動物與OHC的分離:選擇出生7 d的Wistar乳鼠20只,雌雄不限,迅速斷頭,在解剖顯微鏡下取出耳蝸基底膜。基底膜置于濃度為0.25 mg/mL的木瓜蛋白酶中,37 ℃孵育10~15 min,輕柔吹打后用加有10%胎牛血清的DMEM液終止消化。懸液經(jīng)離心后去上清液,加入1 mL培養(yǎng)液在37 ℃,5%CO2孵箱孵育24 h。4%多聚甲醛固定30 min,封閉液(10%山羊血清和1%BSA)30 min,1∶1 000一抗4 ℃過夜,1∶64 FITC兔抗羊IgG二抗(武漢博士德公司生產(chǎn))室溫下孵育1 h,熒光顯微鏡下觀察。
2 結 果
熒光顯微鏡下顯示,經(jīng)Prestin抗體標記的乳鼠單離OHC的側膜可見明顯的Prestin抗體陽性染色;OHC基底部細胞膜亦見陽性染色,但較弱(圖1);側膜與基底部膜結合部位未見陽性染色。
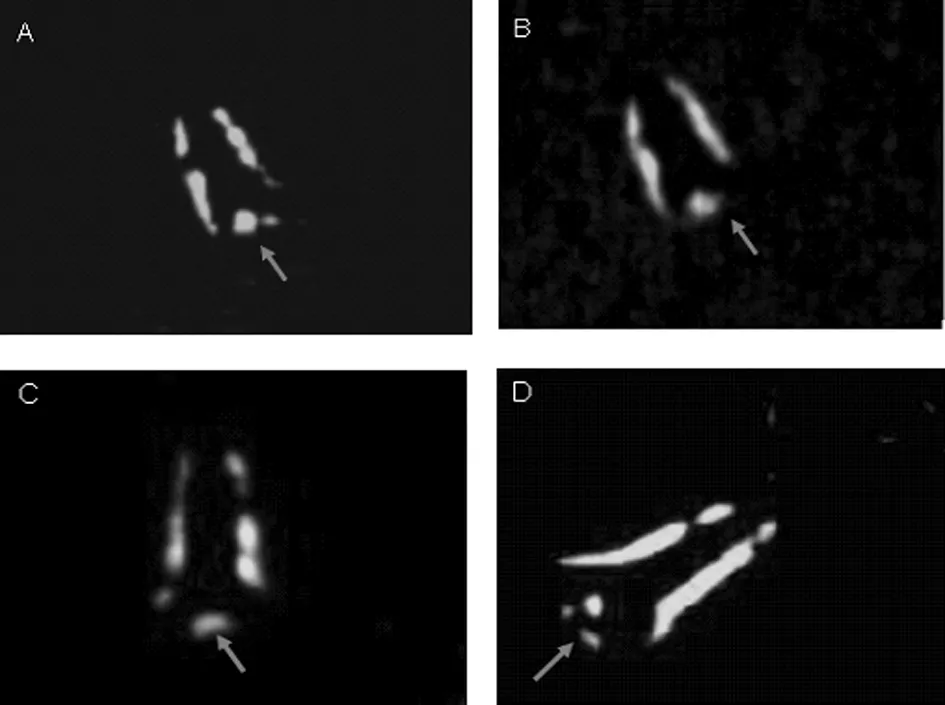
圖1 熒光顯微鏡下基底部細胞膜部位(箭頭所示)
3 討 論
在耳蝸放大機制中,OHC能夠?qū)㈦娦盘栟D(zhuǎn)化為細胞長度的機械性改變[6]。當外界聲音引發(fā)耳蝸內(nèi)液體波動及基底膜機械振動時,內(nèi)毛細胞和OHC表面的纖毛束可將這種機械刺激轉(zhuǎn)換為電信號。其中OHC電位變化除小部分作為聲音信息傳入大腦外,大部分都通過引發(fā)胞體電運動轉(zhuǎn)化為機械能,并反饋給基底膜,進一步增強基底膜的振動,使內(nèi)毛細胞感受的機械刺激得以放大[7]。此過程中,OHC電運動成為關鍵。Prestin作為OHC的運動蛋白,是電運動和耳蝸放大效應的分子基礎,并特異性地在耳蝸OHC側膜上表達,而在耳蝸內(nèi)毛細胞及心臟、腦、腎臟、肺和肌肉等重要組織中均未發(fā)現(xiàn)Prestin表達[8-10]。通過基因敲除動物模型,也進一步證實了Prestin是OHC電運動及耳蝸放大效應所必需的運動蛋白[11]。
近年來,有人運用熒光雙染及復合染色等方法觀察發(fā)現(xiàn),成年大鼠和豚鼠OHC除了側膜有明顯的Prestin抗體陽性染色之外,在OHC基底部的細胞膜上也有較強的陽性染色,同時在細胞核平面以下的側膜即側膜與基底部細胞膜結合處也有陽性染色的存在[5]。
本實驗是以出生7 d乳鼠為研究對象,利用Prestin特異性抗體染色,在熒光顯微鏡下觀察Prestin在乳鼠耳蝸單離OHC細胞膜上的表達與分布。本研究結果顯示:(1)OHC側膜存在較為均一的Prestin抗體陽性染色;(2)OHC底端細胞膜亦見陽性染色,但著色較弱;(3)側膜與底端細胞膜結合部位未見陽性染色。通過本實驗的結果可以推測:成年大鼠OHC基底部細胞膜以及細胞核平面以下的側膜上分布的Prestin蛋白不是一出生就存在的,而是逐步產(chǎn)生增多的。有文獻報道了Prestin的表達可被控制,并且在細胞生命周期中處于不間斷更新過程中,并推測OHC的基底部可能存在Prestin的儲存庫[5,12]。從本實驗結果來看,儲存庫可能存在于OHC側膜,側膜上的Prestin在耳蝸OHC成熟過程中不斷增加,并向基底部膜擴散,這也能比較好地解釋成年Wistar大鼠OHC細胞核平面以上的側膜、基底部膜和細胞核平面以下的側膜上Prestin密度逐漸減少這一現(xiàn)象。
[1]Huang G,Santos-Sacchi J.Mapping the distribution of the outer hair cell motility voltage sensor by electrica1 amputation[J].Biophys J,1993,65(16):2228-2231.
[2]Adler HJ,Belyantseva IA,Merritt RC,et al.Expression of prestin,amembrane motor protein,in the mammalian auditory and vestibular periphery[J].Hear Res,2003,184(1):27-29.
[3]Yu N,Zhai SQ,Yang SM,et al.Distribution of prestin on outer hair cell basolateral surface[J].J Otology,2008,3(2):92-97.
[4]Belyantseva IA,Adler HJ,Curi R,et al. Expression and localization of prestin and the sugar transporter GLUT-5 during development of electromotility in cochlear outer hair cells[J].J Neurosci,2000,20(2):116-118.
[5]于寧,翟所強,趙紅波.外毛細胞基底側膜動力蛋白分布均一性的研究[J].聽力學及言語疾病雜志,2009,17(5):461-463.
[6]Matsumoto N,Kalinec F.Prestin-dependent and prestin-in-dependent motility of guinea pig outer hair cells[J].Hearing Res,2005,208(1):1-13.
[7]陳偉,楊仕明.Prestin-外毛細胞運動蛋白[J].中華耳鼻咽喉頭頸外科雜志,2007,42(5):394-396.
[8]Zheng J,Shen W,He DZ,et al.Prestin is the moter protein of cochlear outer hair cells[J].Nature,2000,40(2):149-153.
[9]Belyantseva IA,Adler HJ,Curi R,et al.Expression and localization of prestin and the sugar transporter GLUT-5 during development of electromotility in cochlear outer hair cells[J].J Neurosci,2000,20(1):116-119.
[10]Zheng J,Long KB,Shen W,et al.Prestine topology:localization of protein epitopes in relation to the plasma membrane[J].Neuroreport,2001,12(8):929-932.
[11]郭運凱,謝鼎華,楊新明.Prestin基因敲除小鼠聽力和毛細胞改變的相關性研究[J].中華耳科學雜志,2004,2(4):245-249.
[12]Yu N,Zhu ML,Zhao HB.Long-term usage of salicylate upregulates prestin expression in the guinea pig cochlea[J].Cell Mo1 Life Sci,2008,65(3):407-409.
