溫度和食物密度對兩種形態型裂足臂尾輪蟲(Brachionus diversicornis)生活史特征和后代形態型的影響*
2011-12-11 08:01:14席貽龍許丹丹葛雅麗
湖泊科學 2011年4期
關鍵詞:差異
馬 杰,席貽龍,許丹丹,葛雅麗
(安徽師范大學生命科學學院,安徽省高校生物環境與生態安全省級重點實驗室,蕪湖241000)
溫度和食物密度對兩種形態型裂足臂尾輪蟲(Brachionus diversicornis)生活史特征和后代形態型的影響*
馬 杰,席貽龍**,許丹丹,葛雅麗
(安徽師范大學生命科學學院,安徽省高校生物環境與生態安全省級重點實驗室,蕪湖241000)
運用單個體培養方法研究了溫度(15℃、20℃、25℃和30℃)和斜生柵藻(Scenedesmus obliquus)密度(0.75×106、1.5 ×106、3.0 ×106和6.0 ×106cells/ml)對等棘刺型和不等棘刺型裂足臂尾輪蟲(Brachionus diversicornis)的凈生殖率、種群內稟增長率、世代時間、生命期望和后代混交率等生活史參數和后代形態型的影響.結果表明:各溫度下,兩種形態型輪蟲的存活率無顯著差異;除20℃外,其余三個溫度下,等棘刺型輪蟲的繁殖率顯著高于不等棘刺型;除30℃下等棘刺型輪蟲的世代時間長于不等棘刺型輪蟲外,各溫度下兩種形態型輪蟲的各生活史參數均無顯著差異.各食物密度下,兩種形態型輪蟲的存活率、繁殖率、各生活史參數也均無顯著差異.兩種形態型輪蟲的種群內稟增長率、世代時間和生命期望對升高的溫度反應不同,但食物密度僅對等棘刺型輪蟲的凈生殖率和內稟增長率有顯著影響.等棘刺型輪蟲的凈生殖率在6.0×106cells/ml食物密度下顯著低于0.75×106和3.0×106cells/ml食物密度下,1.5×106cells/ml食物密度下與上述三食物密度間均無顯著差異;內稟增長率在6.0×106cells/ml食物密度下顯著低于3.0×106cells/ml食物密度下,但兩者均與0.75×106和1.5×106cells/ml食物密度下無顯著差異.在各溫度和食物密度下,兩種形態型輪蟲都能產生等棘刺型和不等棘刺型的后代.25℃下等棘刺型輪蟲所產后代中等棘刺率明顯高于不等棘刺型輪蟲,但各食物密度下兩種形態型輪蟲所產后代中等棘刺率無顯著的差異.溫度對等棘刺型輪蟲所產后代中等棘刺率有顯著的影響,但食物密度對兩種形態型輪蟲所產后代中等棘刺率均無顯著的影響.等棘刺型輪蟲所產后代中等棘刺率在15℃和25℃、20℃和30℃間分別無顯著差異,但前兩者顯著高于后兩者.
裂足臂尾輪蟲;形態型;溫度;食物密度;生活史特征
表型可塑性是生物有機體適應環境變化而改變其表型的能力[1],可塑性的程度和發生時機揭示了生物種群對一系列環境變化的適應性[2-5].多態性在浮游性輪蟲特別是臂尾輪蟲(Brachionus)和龜甲輪蟲(Keratella)中廣泛存在[6-8].自然水體中,萼花臂尾輪蟲(B.calyciflorus)存在幾種形態型[9-10],其中,由晶囊輪蟲(Asplanchna)誘導產生的單一短棘刺型、兩個短棘刺型和兩個長棘刺型的萼花臂尾輪蟲較為常見[7].野外和實驗室內,龜甲輪蟲的多態性也有較多的報道[6,11-12].
溫度、營養狀況和捕食者等環境因子均可以誘導多態性輪蟲的形態發生變化,并改變其生理、生活史和行為等特征[13-15].溫度對短棘刺的尾突臂尾輪蟲(B.caudatus f.apsteini)的后代棘刺長短有顯著的影響,而對長棘刺型的后代影響不顯著;低溫可誘導尾突臂尾輪蟲形成后棘刺[8],或延長一些種類輪蟲的后棘刺[6,16-17].捕食者的存在和低的食物密度均能夠誘導臂尾輪蟲和龜甲輪蟲的后側棘刺和后棘刺的延伸[7,11,18-20].在較高的食物密度下,無棘刺型的龜形龜甲輪蟲(K.testudo)的種群內稟增長率大于有棘刺型的龜形龜甲輪蟲,且前者在培養開始后的3-4d有較高的繁殖率和存活率[21];無棘刺型的萼花臂尾輪蟲比有棘刺型的萼花臂尾輪蟲有較高的種群內稟增長率[22];溫度對兩種形態型萼花臂尾輪蟲的生活史特征和后代形態型有顯著的影響[23].
裂足臂尾輪蟲是典型的、棲息于淺水池塘中的浮游性輪蟲;根據背甲后端左右兩根棘刺長度的不同,可以把裂足臂尾輪蟲分為后端棘刺不等長的不等棘刺型和后端棘刺等長的等棘刺型[24].迄今為止,有關這兩種形態型間的生活史特征和后代形態型差異研究尚未見報道.因此,本文運用單個體培養方法,比較研究不同溫度(15℃、20℃、25℃和30℃)和食物密度(0.75 ×106、1.5 ×106、3.0 ×106和 6.0 ×106cells/ml)下兩種形態型裂足臂尾輪蟲的生活史特征和后代形態型,為不同形態型輪蟲的生活史對策研究積累資料.
1 材料與方法
1.1 實驗材料
實驗所用裂足臂尾輪蟲于2009年4月中旬采自蕪湖市鏡湖.采樣后,隨機挑取等棘刺型和不等棘刺型非混交雌體,于(22±1)℃、自然光照(光照強度約130 lx)條件下進行克隆培養.輪蟲培養液采用Gilbert配方[25](pH 為7.3),所用的餌料為 HB-4 培養基[26]培養的、處于指數增長期的斜生柵藻(Scenedesmus obliquus).
1.2 輪蟲的預培養
實驗前,將兩種形態型輪蟲分別進行預培養.不同溫度下的預培養是將兩種形態型輪蟲分別置于溫度為15℃、20℃、25℃和30℃,斜生柵藻密度為1.5×106cells/ml的條件下進行1周的預培養;不同食物密度下的預培養是用密度分別為0.75 ×106、1.5 ×106、3.0 ×106和6.0 ×106cells/ml的斜生柵藻作為兩種形態型輪蟲的食物,在22℃下進行1周的預培養.預培養過程中,每12h懸浮沉積于試管底部的藻類食物;每24h更換輪蟲培養液并投喂餌料1次,同時通過去除一部分個體使得輪蟲種群始終處于指數增長期.
1.3 單個體培養實驗
將攜帶非混交卵的輪蟲單個培養于帶凹穴的塑料板中,培養液體積為0.5ml.每間隔4h在解剖鏡下檢查輪蟲卵的孵化情況,取剛孵化出的幼體開始實驗.實驗過程中,每間隔12h觀察1次(30℃下每8h觀察一次),記錄輪蟲所產的幼體數及母體的存活情況;并移出所產幼體,在相同的條件下繼續培養待其產卵后確定雌體類型和后代形態型;每間隔24h更換1次培養液并投喂藻類食物.四種溫度下,各形態型輪蟲的克隆數為4個(重復),輪蟲克隆均使用15個齡長小于4h的個體;每種食物密度下,各形態型輪蟲的克隆數為3個(重復),輪蟲克隆也均使用15個個體.實驗條件分別與預培養時相同,實驗至全部母體死亡為止.
1.4 生活史參數及其計算方法
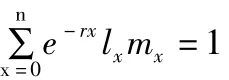
1.5 數據的處理和分析
采用SPSS統計分析軟件對上述數據進行統計分析.對所得的各生活史參數進行正態分布檢驗后,對符合正態分布的數據進行方差分析和多重比較以揭示兩種形態型輪蟲各生命表統計學參數在不同溫度、不同食物密度間的差異.對同一溫度或同一食物密度下兩種形態型輪蟲的各生活史參數的差異顯著性進行兩獨立樣本t檢驗.運用生存分析中的Kaplan-Meier分析不同溫度和食物密度下兩種形態型輪蟲的生存時間的差異顯著性.
2 結果與分析
2.1 溫度對兩種形態型輪蟲生活史特征和后代形態型的影響
兩種形態型輪蟲的存活率均隨著溫度的升高而顯著降低(P<0.05);各溫度下,兩種形態型輪蟲的存活率無顯著差異(P>0.05)(圖1).
等棘刺型輪蟲的繁殖率隨著溫度的升高而顯著升高(P<0.05),不等棘刺型輪蟲的繁殖率在20℃、25℃和30℃下無顯著的差異,但均顯著高于15℃下(P<0.05).除20℃外,其余三個溫度下,等棘刺型輪蟲的繁殖率顯著高于不等棘刺型(P<0.05)(圖1).
兩種形態型輪蟲各生命表統計學參數以及后代等棘刺率之間的差異因溫度的不同而異.15℃和20℃下,兩種形態型輪蟲的各生命表統計學參數和后代等棘刺率均無顯著差異.25℃下,等棘刺型輪蟲所產后代中等棘刺率明顯高于不等棘刺型輪蟲(P<0.05),但兩種形態型輪蟲的各生命表統計學參數均無顯著差異.30℃下,等棘刺型輪蟲的世代時間長于不等棘刺型輪蟲(P<0.05),其余各生命表統計學參數和后代等棘刺率均無顯著差異(表1).
溫度對兩種形態型輪蟲的內稟增長率、世代時間和生命期望均有極顯著的影響(P<0.01),對等棘刺型輪蟲的后代等棘刺率影響也極顯著(P<0.01).兩種形態型輪蟲的內稟增長率總體上均隨溫度的升高而增大,但等棘刺型輪蟲的內稟增長率在15℃和20℃、25℃和30℃下分別無顯著的差異(P>0.05),但后兩者顯著高于前兩者;不等棘刺型輪蟲的內稟增長率在15℃下顯著低于25℃和30℃下,但三者均與20℃下的內稟增長率差異不顯著(P>0.05).兩種形態型輪蟲的世代時間均隨溫度的升高而顯著縮短,但縮短的速率也存在著差異.等棘刺型輪蟲的生命期望在15℃下最長,20℃下顯著長于30℃下,而25℃下與20℃和30℃下均無顯著的差異;不等棘刺型輪蟲的生命期望在15℃下最長,30℃下最短,20℃和25℃下居中且兩者無顯著差異.等棘刺型輪蟲的后代等棘刺率在15℃和25℃、20℃和30℃下分別無顯著的差異,但前兩者顯著高于后兩者(表1).
雙因素方差分析結果表明,溫度對輪蟲的內稟增長率、世代時間、生命期望和后代等棘刺率均有顯著影響(P<0.05),形態型對輪蟲的所有生活史參數及后代等棘刺率均無顯著影響(P>0.05),溫度和形態型的交互作用對輪蟲的后代等棘刺率有顯著影響(P<0.05).四個溫度下,25℃和30℃下輪蟲的內稟增長率較高,而15℃和20℃下較低;15℃下輪蟲的世代時間和生命期望最長,30℃下最短;15℃下輪蟲的后代等棘刺率較高,20℃和30℃下較低.
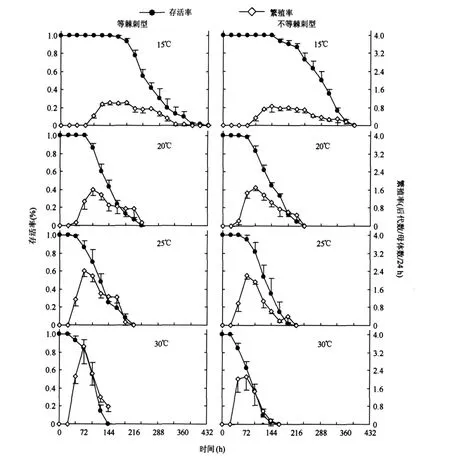
圖1 四種溫度下兩種形態型裂足臂尾輪蟲的存活率和繁殖率Fig.1 Survivorship and fecundity of two B.diversicornis morphotypes at four temperatures
2.2 食物密度對兩種形態型輪蟲的生活史特征和后代形態型的影響
四種食物密度處理間,0.75×106cells/ml食物密度下兩種形態型輪蟲的存活率均較高,3.0×106和6.0×106cells/ml的食物密度下兩種形態型輪蟲的存活率均較低.各食物密度下,兩種形態型輪蟲的存活率均無顯著差異(P>0.05)(圖2).
食物密度對等棘刺型輪蟲的繁殖率有極顯著影響(P<0.01),對不等棘刺型輪蟲的繁殖率無顯著影響(P >0.05).等棘刺型輪蟲的繁殖率在0.75 ×106和3.0 ×106cells/ml食物密度下顯著高于 1.5 ×106和 6.0×106cells/ml食物密度下.各食物密度下,兩種形態型輪蟲的繁殖率均無顯著的差異(P>0.05)(圖2).

表1 四種溫度下兩種形態型裂足臂尾輪蟲的生活史參數和后代等棘刺率*Tab.1 Life history parameters and proportion of equal-spined offspring produced by two B.diversicornis morphotypes at four temperatures
四種食物密度下,兩種形態型輪蟲間的生活史參數和后代等棘刺率均無顯著的差異(表2).

表2 四種食物密度下兩種形態型裂足臂尾輪蟲的生活史參數和后代等棘刺率*Tab.2 Life history parameters and proportion of equal-spined offspring produced by two B.diversicornis morphotypes cultured at four food densities
食物密度僅對等棘刺型輪蟲的凈生殖率和內稟增長率有顯著影響(P<0.05).等棘刺型輪蟲的凈生殖率在6.0 ×106cells/ml食物密度下顯著小于0.75 ×106和3.0 ×106cells/ml食物密度下,1.5 ×106cells/ml食物密度下與上述三食物密度間均無顯著差異;內稟增長率在6.0×106cells/ml食物密度下顯著小于3.0×106cells/ml食物密度下,但兩者均與0.75×106和1.5×106cells/ml食物密度下無顯著差異.

圖2 四種食物密度下兩種形態型裂足臂尾輪蟲的存活率和繁殖率Fig.2 Survivorship and fecundity of two B.diversicornis morphotypes cultured at four food densities
雙因素方差分析結果表明,食物密度對輪蟲的內稟增長率有顯著的影響(P<0.05),形態型對輪蟲的后代等棘刺率有顯著的影響(P<0.05),食物密度和形態型間的交互作用對輪蟲的生活史參數和后代等棘刺率均無顯著的影響.在四個食物密度下,6.0×106cells/ml食物密度下輪蟲的內稟增長率較高,其它三個食物密度下較低;等棘刺型輪蟲所產的后代中等棘刺率顯著高于不等棘刺型.
3 討論
3.1 不同形態型輪蟲的生活史特征
作為防御捕食的器官,后(側)棘刺的形成和生長需要消耗能量.因此,在較高的食物密度下,無棘刺型的龜形龜甲輪蟲比有棘刺型的龜形龜甲輪蟲在培養開始后的3-4d有較高的繁殖率和存活率[21].與其相似的是,本研究中,在3.0×106cells/ml食物密度下,等棘刺型裂足臂尾輪蟲的繁殖率高于不等棘刺型.
在較高的溫度(20℃-30℃)或較低的食物密度下,當萼花臂尾輪蟲或龜形龜甲輪蟲通過攝食所獲取的能量有限或被較多地用于代謝需求時,投入生殖以及后(側)棘刺的形成和生長的能量相對較少,因此它們的種群內稟增長率沒有顯著的差異;但在較低的溫度(15℃)或較高的食物密度下,當上述兩種輪蟲通過攝食所獲取的能量除了滿足代謝需求外,尚有較多的能量可用于生殖以及后(側)棘刺的形成和生長,此時后(側)棘刺的形成和生長消耗了部分能量,由此導致無棘刺型萼花臂尾輪蟲和龜形龜甲輪蟲的種群內稟增長率高于雙棘刺型萼花臂尾輪蟲和具棘刺型龜形龜甲輪蟲的種群內稟增長率[21-23].本研究中,在各溫度和食物密度下,兩種形態型裂足臂尾輪蟲的種群內稟增長率等生活史參數均無顯著差異;這可能與它們在等棘刺和不等棘刺的形成和生長過程中所消耗的能量相似有關.
已有研究結果表明,不同形態型輪蟲的生活史參數對升溫的響應存在著差異.當溫度從20℃升高至30℃時,無后側棘刺型萼花臂尾輪蟲的平均壽命縮短的速率高于雙后側棘刺型;雙后側棘刺型萼花臂尾輪蟲的凈生殖率在30℃下最高,無后側棘刺型萼花臂尾輪蟲的凈生殖率受溫度影響不顯著;當溫度從15℃升高至20℃時,無后側棘刺型萼花臂尾輪蟲的種群內稟增長率下降的速率大于雙后側棘刺型[23].與上述研究結果相似,本研究發現,兩種形態型裂足臂尾輪蟲的內稟增長率、世代時間和生命期望對升高的溫度的反應存在著差異.此外,本研究還首次發現,裂足臂尾輪蟲的凈生殖率和內稟增長率對升高的食物密度的反應也因形態型的不同而異;食物密度僅對等棘刺型輪蟲的凈生殖率和內稟增長率有顯著影響.
3.2 不同形態型輪蟲所產后代的形態型
不同形態型輪蟲所產后代的形態型與晶囊輪蟲的捕食、輪蟲母體的年齡、孤雌生殖世代數和溫度等有關.在沒有晶囊輪蟲誘導的情況下,具棘刺型龜形龜甲輪蟲和螺形龜甲輪蟲通常產出無后棘刺型后代;有些情況下,一些具后棘刺型的龜形龜甲輪蟲也可產出短后棘刺型后代[12,21].然而,一些品系的萼花臂尾輪蟲即使在沒有晶囊輪蟲和其他環境刺激的情況下也形成長后棘刺,后棘刺的長度取決于母體的年齡[28].由休眠卵孵出的萼花臂尾輪蟲和方形臂尾輪蟲(B.quadridentatus)干雌體(stem female)不形成后側棘刺,但隨著孤雌生殖世代數的增加而形成后側棘刺[18,29-30].溫度對短后棘刺型尾突臂尾輪蟲的后代形態型具有顯著影響,而對長后棘刺型的后代形態型無顯著影響;低溫可誘導短后棘刺型尾突臂尾輪蟲產生后棘刺型后代[8].在15℃、20℃、25℃和30℃下,無后棘刺型和雙后棘刺型的萼花臂尾輪蟲均可產生無后棘刺型和雙后棘刺型的后代;15℃下,雙后棘刺型的萼花臂尾輪蟲也可產生單后棘刺型的后代.溫度對雙后棘刺型萼花臂尾輪蟲后代后側棘刺的形成無顯著影響,但無后棘刺型萼花臂尾輪蟲在15℃下產生的具后側棘刺后代的比例高于20℃下[37].與萼花臂尾輪蟲相似,本研究中,在各溫度和食物密度下,兩種形態型裂足臂尾輪蟲都可以產生等棘刺型和不等棘刺型后代;25℃下,等棘刺型裂足臂尾輪蟲所產后代中等棘刺率明顯高于不等棘刺型輪蟲,但在15℃、20℃和30℃下兩種形態型輪蟲所產后代中等棘刺率均無顯著性差異.溫度對等棘刺型裂足臂尾輪蟲的后代形態型有顯著影響,對不等棘刺型輪蟲的后代形態型無顯著影響;等棘刺型輪蟲的后代等棘刺率在15℃和25℃、20℃和30℃間分別無顯著差異,但前兩者顯著高于后兩者.此外,本研究還首次發現,食物密度對兩種形態型裂足臂尾輪蟲的后代形態型均無顯著的影響.
[1]West-Eberhard MJ.Developmental Plasticity and Evolution.Oxford:Oxford University Press,2003.
[2]Hjelm J,Svanb?ck R,Bystr?m P et al.Diet-dependent body morphology and ontogenetic reaction norms in Eurasian perch.Oikos,2001,95(2):311-323.
[3]Dahl J,Peckarsky BL.Induced morphological defenses in the wild:predator effects on a mayfly,Drunella coloradensis.Ecology,2002,83(6):1620-1634.
[4]Trussell GC,Nicklin MO.Cue sensitivity,inducible defense,and trade-offs in a marine snail.Ecology,2002,83(6):1635-1647.
[5]Michimae H.Differentiated phenotypic plasticity in larvae of the cannibalistic salamander Hynobius retardatus.Behavioral Ecology and Sociobiology,2006,60(2):205-211.
[6]Xi YL,Jin HJ,Xie P et al.Morphological variation of Keratella cochlearis(Rotatoria)in a shallow,eutrophic subtropic Chinese lake.Journal of Freshwater Ecology,2002,17(3):447-454.
[7]Yin XW,Niu CJ.Polymorphism and morphotype transformations in the rotifer(Brachionus calyciflorus).Zoological Re-search,2007,28(1):68-72.
[8]Athibai S,Sanoamuang LO.Effect of temperature on fecundity,life span and morphology of long-and short-spined clones of Brachionus caudatus f.apsteini(Rotifera).International Review of Hydrobiology,2008,93(6):690-699.
[9]Koste W.Rotatoria.Die R?dertiere Mitteleuropas.(überordnung monogononta),Bestimmungswerk begründet von Max Voigt.2 vols.Berlin:Gebrüder Borntraeger,1978.
[10]李化炳,席貽龍,程新峰等.輪蟲同域性物種形成:來自萼花臂尾輪蟲克隆間的分子系統發育關系和生殖隔離證據.動物學報,2008,54(2):256-264.
[11]Gilbert JJ,Stemberger RS.Asplanchna-induced polymorphism in the rotifer Keratella slacki.Limnology and Oceanography,1984,29(6):1309-1316.
[12]Stemberger RS,Gilbert JJ.Multiple-species induction of morphological defenses in the rotifer Keratella testudo.Ecology,1987,68(2):370-378.
[13]Saksena DN,Kulkarni N.Polymorphosis in a brachionid rotifer,Brachionus quadridentatus Hermann from Morar channel,Gwalior(India).Proceedings:Animal Sciences,1986,95(3):365-369.
[14]Sanoamuang LO.The effect of temperature on morphology,life history and growth rate of Filinia terminalis(Plate)and Filinia cf.pejleri Hutchinson in culture.Freshwater Biology,1993,30(2):257-267.
[15]Green J.Morphological variation of Keratella cochlearis(Gosse)in Myanmar(Burma)in relation to zooplankton community structure.Hydrobiologia,2007,593(1):5-12.
[16]Bielan?ska-Grajner I.Influence of temperature on morphological variation in populations of Keratella cochlearis(Gosse)in Rybnik Reservoir.Hydrobiologia,1995,313/314(1):139-146.
[17]Diéguez M,Modenutti B,Queimali?os C.Influence of abiotic and biotic factors on morphological variation of Keratella cochlearis(Gosse)in a small Andean lake.Hydrobiologia,1998,387/388:289-294.
[18]Gilbert JJ.Spine development in Brachionus quadridentatus from an Australian billabong:genetic variation and induction by Asplanchna.Hydrobiologia,2001,446/447(1):19-28.
[19]Pavón-Meza EL,Sarma SSS,Nandini S.Combined effects of temperature,food(Chlorella vulgaris)concentration and predation(Asplanchna girodi)on the morphology of Brachionus havanaensis(Rotifera).Hydrobiologia,2007,593(1):95-101.
[20]Sarma SSS,Nandini S.Small prey size offers immunity to predation:a case study on two species of Asplanchna and three brachionid prey(Rotifera).Hydrobiologia,2007,593(1):67-76.
[21]Stemberger RS.Reproduction costs and hydrodynamic benefits of chemically induced defenses in Keratella testudo.Limnology and Oceanography,1988,33(4):593-606.
[22]Gilbert JJ.Females from resting eggs and parthenogenetic eggs in the rotifer Brachionus calyciflorus:lipid droplets,starvation resistance and reproduction.Freshwater Biology,2004,49(11):1505-1505.
[23]Xiang XL,Xi YL,Zhang JY et al.Effect of temperature on survival,reproduction and morphotype in offspring of two Brachionus calyciflorus(Rotifera)morphotypes.Journal of Freshwater Ecology,2010,25(1):9-18.
[24]王家楫.中國淡水輪蟲志.北京:科學出版社,1961:77-78.
[25]Gilbert JJ.Mictic female production in rotifer Brachionus calyciflorus.Journal of Experimental Zoology,1963,153(2):113-124.
[26]章宗涉,黃祥飛.淡水浮游生物研究方法.北京:科學出版社,1991.
[27]胡 柯,席貽龍,王金霞.不同地理區域湖泊中萼花臂尾輪蟲的生活史特征.湖泊科學,2010,22(4):585-590.
[28]Schr?der T,Gilbert JJ.Maternal age and spine development in the rotifer Brachionus calyciflorus:increase of spine length with birth orders.Freshwater Biology,2009,54(5):1054-1065.
[29]Yin XW,Niu CJ.Polymorphism in stem females and successive parthenogenetic generations in Brachionus calyciflorus Pallas.Aquatic Ecology,2008,42(3):415-420.
[30]Gilbert JJ,Schr?der T.Rotifers from diapausing,fertilized eggs:unique features and emergence.Limnology and Oceanography,2004,49(4):1341-1354.
Effects of temperature and algal density on life history characteristics and morphotype in offspring of two Brachionus diversicornis morphotypes
MA Jie,XI Yilong,XU Dandan&GE Yali
(College of Life Sciences,Anhui Normal University,Provincial Key Laboratory of Biotic Environment and Ecological Safety,Wuhu 241000,P.R.China)
Effects of temperature(15℃,20℃,25℃ and 30℃)and Scenedesmus obliquus density(0.75 ×106,1.5×106,3.0×106and 6.0 ×106cells/ml)on life history parameters including net reproductive rate,intrinsic rate of population increase,generation time,life expectancy at hatching and proportion of mictic offspring,and morphotype in offspring of two Brachionus diversicornis morphotypes were studied by means of individual culture.The results showed that at the four temperatures,the survivorship between the two B.diversicornis morphotypes was similar.The fecundities of the equal-spined rotifers at 15℃,25℃ and 30℃ were significantly higher than those of the unequal-spined rotifer.Each of the life history parameters between the two B.diversicornis morphotypes was similar,except that the generation time of the equal-spined rotifers was longer than that of the unequal-spined rotifers at 30℃.The survivorship,the fecundity and each of all the life history parameters between the two B.diversicornis morphotypes were also similar at the four food densities.All the intrinsic rate of population increased,the generation time and the life ex-pectancy at hatching of the two B.diversicornis morphotypes responded differently to increasing temperature,but food density only affected the net reproductive rate and the intrinsic rate of population increase of the equal-spined rotifers significantly.The net reproductive rate of the equal-spined rotifers at algal density of 6.0 ×106cells/ml was lower than that at both 0.75 ×106and 3.0 ×106cells/ml,and the net reproductive rate at 1.5 × 106cells/ml was similar with that at each of all the above-stated three food densities.The intrinsic rate of population increase of the equal-spined rotifers at 6.0 ×106cells/ml was lower than that at 3.0 ×106cells/ml,but both of them were similar with those at 0.75 ×106and 1.5 ×106cells/ml.At each of the four temperatures and food densities,the two B.diversicornis morphotypes produced both equal-and unequal-spined offspring.Only at 25℃,the proportion of equal-spined offspring produced by the equal-spined mothers was higher than that produced by the unequal-spined mothers.However,at each of all the four food densities,the proportion of equal-spined offspring produced by the two B.diversicornis morphotypes was similar.Temperature affected the proportion of equal-spined offspring produced by the equal-spined mothers significantly,but food density did not affect the proportion of equal-spined offspring produced by both the equal-and unequal-spined mothers.The proportions of equal-spined offspring produced by the equal-spined mothers at 15℃ and 25℃,and 20℃ and 30℃ were similar,respectively,but the former two were higher than the latter two.
Brachionus diversicornis;morphotype;temperature;algal density;life history characteristics
* 國家自然科學基金項目(30870369,30499341)、安徽省優秀青年基金項目(08040106904)、安徽省高校生物環境與生態安全省級重點實驗室專項基金項目(2004sys003)和安徽省重點實驗室專項基金項目聯合資助.2010-09-17收稿;2010-10-19收修改稿.馬杰,女,1987年生,碩士研究生;E-mail:jiema_2008@126.com.
** 通訊作者;E-mail:ylxi1965@yahoo.com.cn.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
