太湖微囊藻毒素時空分布特征及與環(huán)境因子的關(guān)系*
2011-12-11 08:11:56王經(jīng)結(jié)鮮啟鳴李正魁耿金菊
湖泊科學(xué) 2011年4期
關(guān)鍵詞:環(huán)境
王經(jīng)結(jié),楊 佳,鮮啟鳴,錢 新,李正魁,耿金菊,孫 成
(南京大學(xué)環(huán)境學(xué)院污染控制與資源化研究國家重點(diǎn)實(shí)驗室,南京210093)
太湖微囊藻毒素時空分布特征及與環(huán)境因子的關(guān)系*
王經(jīng)結(jié),楊 佳,鮮啟鳴**,錢 新,李正魁,耿金菊,孫 成
(南京大學(xué)環(huán)境學(xué)院污染控制與資源化研究國家重點(diǎn)實(shí)驗室,南京210093)
采用高效液相色譜法(HPLC)對太湖水體中溶解態(tài)微囊藻毒素(MC-LR;MC-RR)濃度進(jìn)行檢測,探討微囊藻毒素濃度的時空變化及其與氮、磷、總有機(jī)碳、藍(lán)藻生物量以及溫度等湖泊環(huán)境因子之間的關(guān)系.結(jié)果表明:微囊藻毒素表底層濃度基本一致;在一天之內(nèi)的變化沒有明顯規(guī)律;冬季微囊藻毒素濃度較高;位于竺山灣和西部沿岸地區(qū)的兩個點(diǎn)(N5和N1)的微囊藻毒素濃度在一年之中變動較大;MC-LR濃度一般大于MC-RR,但在8,9兩月出現(xiàn)相反的結(jié)果.水中溶解性微囊藻毒素與各環(huán)境因子的相關(guān)性分析表明微囊藻毒素與總氮呈極顯著正相關(guān),與總磷無顯著相關(guān)性.在藍(lán)藻暴發(fā)期,MCLR與總氮、總磷、SS、總有機(jī)碳、藍(lán)藻生物量呈極顯著正相關(guān);MC-RR與所測的環(huán)境因子皆無顯著相關(guān)性.
微囊藻毒素;太湖;環(huán)境因子;相關(guān)性
近年來,隨著經(jīng)濟(jì)的迅猛發(fā)展,水體富營養(yǎng)化程度加劇,有害藍(lán)藻水華發(fā)生日趨頻繁,這引起國內(nèi)外研究人員的普遍關(guān)注.藍(lán)藻水華暴發(fā)不僅造成水質(zhì)下降,而且微囊藻(Microcystis)、魚腥藻(Anabaena)、顫藻(Oscillatoria)等還向水體釋放微囊藻毒素(Microcystins,MCs),危害水生生態(tài)系統(tǒng)的安全[1].MCs是一類單環(huán)七肽化合物,具有多種同分異構(gòu)體[2],主要類型有3種,分別是MC-LR、MC-RR和MC-YR(L、R、Y分別代表亮氨酸、精氨酸和酪氨酸),其中MC-LR和MC-RR所占比例較高,是主要研究對象[3-4].MCs具有肝臟毒性和腫瘤促進(jìn)作用[5],它不僅對水生生物產(chǎn)生毒害作用,還可能通過飲用水和食物鏈的生物富集危害人類健康.影響藍(lán)藻生長及產(chǎn)毒能力的環(huán)境因素眾多,不同時間不同藻的產(chǎn)毒能力不同,產(chǎn)生的毒素種類也不一定相同[6],因此要了解MCs在自然水體中的分布需要綜合考慮各種環(huán)境因子的作用.
太湖是我國第三大淡水湖,現(xiàn)有水面面積2338km2,是典型的淺水湖泊,同時也是藍(lán)藻水華多發(fā)的湖泊.本研究選取太湖作為研究對象,對水中MC-LR和MC-RR進(jìn)行為期一年的監(jiān)測,研究藻毒素在水體中的時空分布特征及影響因素,以探索MCs在自然水體中的形成規(guī)律,為研究藻毒素的生態(tài)風(fēng)險提供理論依據(jù).
1 材料與方法
1.1 采樣點(diǎn)布設(shè)
根據(jù)太湖湖底地形、湖面面積、湖區(qū)地理位置、富營養(yǎng)狀況及主要分布特征等,在太湖選取了九個點(diǎn)(圖1),北湖五個點(diǎn),南湖四個點(diǎn),這九個點(diǎn)覆蓋整個太湖,并具有較好的代表性.
1.2 樣品采集
2009年2月到2009年5月,2009年8月到2009年9每月采樣一次,2009年11月和2010年1月各采樣一次.在七月初藍(lán)藻水華高峰期選擇三天對N1,N2兩點(diǎn)進(jìn)行重點(diǎn)監(jiān)測,每天采樣五次,采樣時間分別為8:00,10:00,12:00,14:00,16:00.使用有機(jī)玻璃采水器,采集水面下 0.5m(表層)和2m(底層)處的水樣,除N1,N2分層儲存外,其他點(diǎn)均將水樣混勻后再儲存.采樣船只采用湖中地物結(jié)合GPS定位儀進(jìn)行定位.待所有的樣品采集完畢后,立即送回實(shí)驗室,根據(jù)不同監(jiān)測指標(biāo)的具體要求保存樣品,待測.
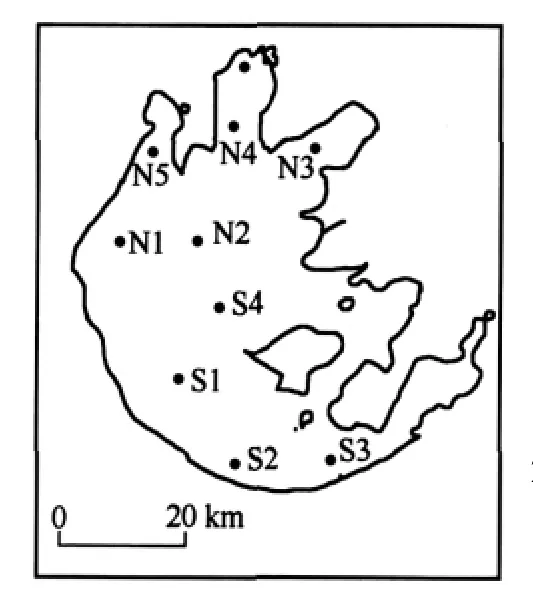
圖1 采樣點(diǎn)位置Fig.1 Sampling sites in Lake Taihu
1.3 水質(zhì)指標(biāo)的測定方法
1.3.1 常規(guī)指標(biāo)的分析測定 懸浮質(zhì)(SS)為103℃-105℃烘干的不可濾殘渣(懸浮物)[7],水溫采用Aaq1183水質(zhì)自動測定儀測定,總磷(TP)、總?cè)芙鈶B(tài)磷(TSP)、溶解態(tài)反應(yīng)磷(SRP)采用過硫酸鉀-鉬銻抗分光光度法,總氮(TN)采用過硫酸鉀氧化-紫外分光光度法,亞硝態(tài)氮(NO2-N)采用N-(1-萘基)-乙二胺光度法,氨氮(NH3-N)采用納氏試劑光度法[7],硝態(tài)氮(NO3-N)采用麝香草酚分光光度法,總有機(jī)碳(TOC)采用過0.45μm濾膜后Aurora1030D濕法(TC-IC模式)測定,UV254采用過0.45μm濾膜在254nm下分光光度法測定[8];藍(lán)藻生物量經(jīng)魯哥試劑固定、沉淀、濃縮后高倍顯微鏡計數(shù)得到[9].
1.3.2 微囊藻毒素的分析測定 量取1L經(jīng)0.45μm濾膜過濾(水體中浮游生物和懸浮物含量較大時可先用500目不銹鋼篩過濾)的水樣置于棕色瓶中.連接SPE固相萃取裝置將水樣通過預(yù)先活化過的Sep-pak小柱(控制流速為8-10ml/min).富集完畢后,依次用10ml純水和10ml體積分?jǐn)?shù)為20%的甲醇溶液淋洗固相萃取小柱.再用10ml含1‰三氟乙酸的甲醇溶液洗脫,洗脫液經(jīng)氮吹后用50%的甲醇定容到500μl,待液相測定.HPLC測定條件:進(jìn)樣量20μl;流動相為57∶43的甲醇與磷酸鹽緩沖液;柱溫為40℃;流速為1ml/min;檢測波長為238nm.MC-LR和MC-RR標(biāo)準(zhǔn)樣品均購自中科院武漢水生生物研究所(純度達(dá)99%).
1.4 統(tǒng)計方法
將所有數(shù)據(jù)在Excel中建立數(shù)據(jù)庫,用SPSS 15.0對水質(zhì)數(shù)據(jù)進(jìn)行Spearman相關(guān)性分析以及逐步回歸分析,借助SigmaPlot 10.0軟件進(jìn)行圖表處理.
2 結(jié)果與討論
2.1 太湖水體中MCs的分布
2.1.1 MCs濃度的深度差異 MCs在表底層的分布具有很好的一致性(圖2).這一規(guī)律在7月份藍(lán)藻暴發(fā)期的連續(xù)監(jiān)測中也得到體現(xiàn).這是由于太湖平均深度較淺(<2m),難以形成分層現(xiàn)象,且面積較大(2338km2),受風(fēng)浪擾動較大,易混合均勻;同時MCs為極性分子,在水中具有較大的溶解度(>1g/L),比較容易擴(kuò)散.
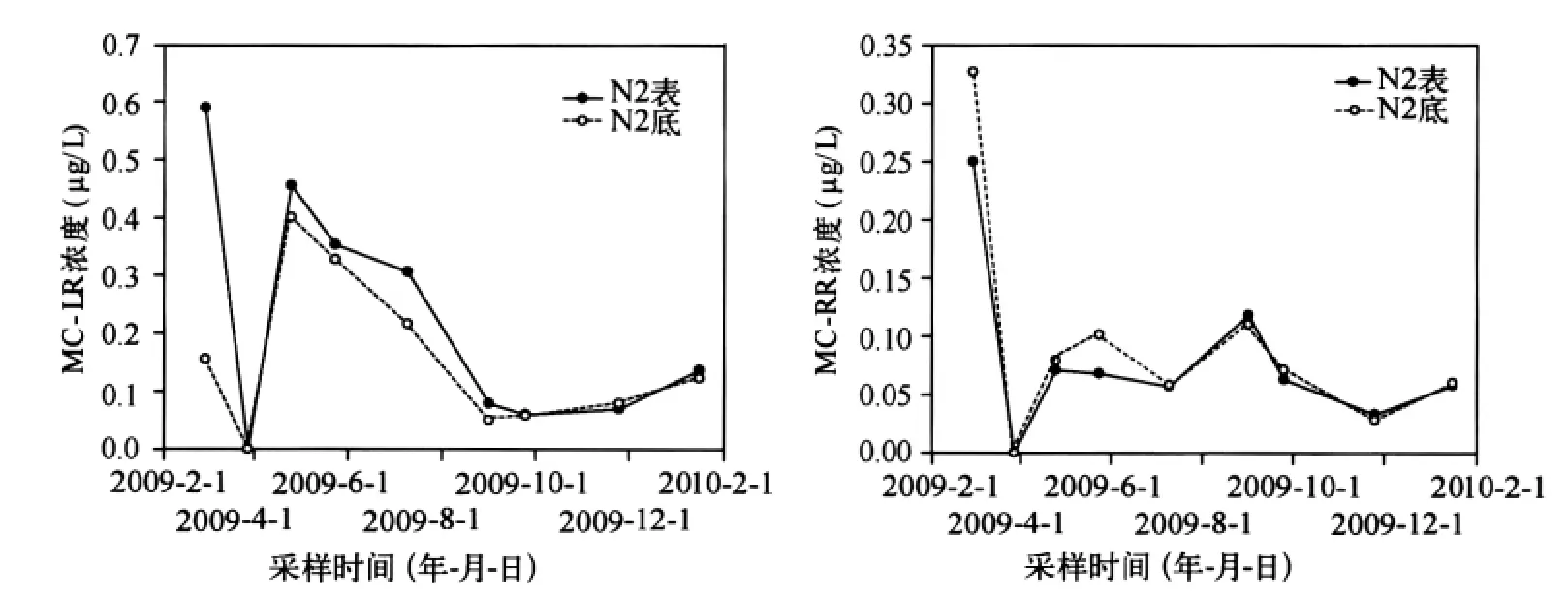
圖2 MCs在水體表底層的濃度變化Fig.2 The distribution of MCs in the bottom and surface of water
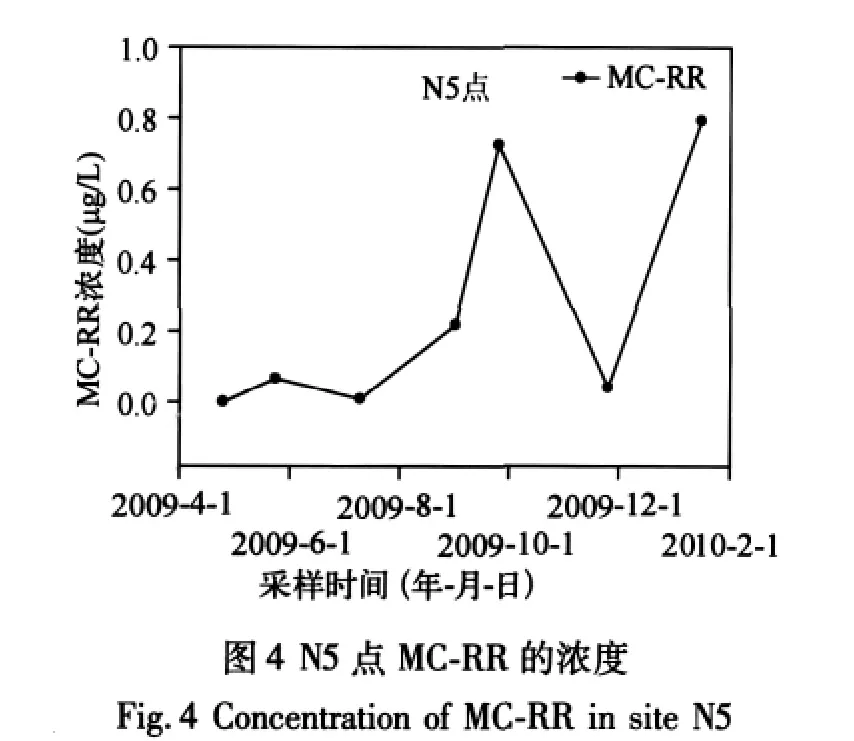
2.1.2 MCs濃度的空間分布 對于 MC-LR,其空間分布特征表現(xiàn)為:S4、S1>S2>S3,N5>N2、N4>N3(圖3).而MC-RR的空間分布特征為:S2>S4>S1、S3,N1>N2、N4>N3(圖3).無論 MC-LR 還是 MC-RR,濃度均低于WHO和我國生活飲用水推薦的標(biāo)準(zhǔn)(1μg/L).MCs在南北湖區(qū)之間未顯示出區(qū)域性分布差異,在N5點(diǎn)顯示出年際波動,MC-RR表現(xiàn)的尤其明顯.09年4月份低于檢測限,而2009年9月份濃度高達(dá)0.7μg/L(圖4).在藍(lán)藻暴發(fā)之前N1點(diǎn)的MC-LR濃度低于N2點(diǎn),暴發(fā)后濃度高于N2點(diǎn),說明藍(lán)藻暴發(fā)促進(jìn)了MC-LR在N1點(diǎn)的積累.這可能是因為N1點(diǎn)靠近西北岸邊,夏季太湖地區(qū)以東南風(fēng)為主,使得藻在N1點(diǎn)聚集,從而導(dǎo)致該點(diǎn)濃度的增加.同時N1點(diǎn)位于東氿湖下方,上游來水的影響較大.所以N1濃度的高低受釋放到水中的藻毒素含量以及水量的雙重影響.九個點(diǎn)中只有N5和N1處于2006年水質(zhì)調(diào)查[10]的劣V類區(qū)域之中,其中是否存在關(guān)系,還需要進(jìn)一步調(diào)查研究.
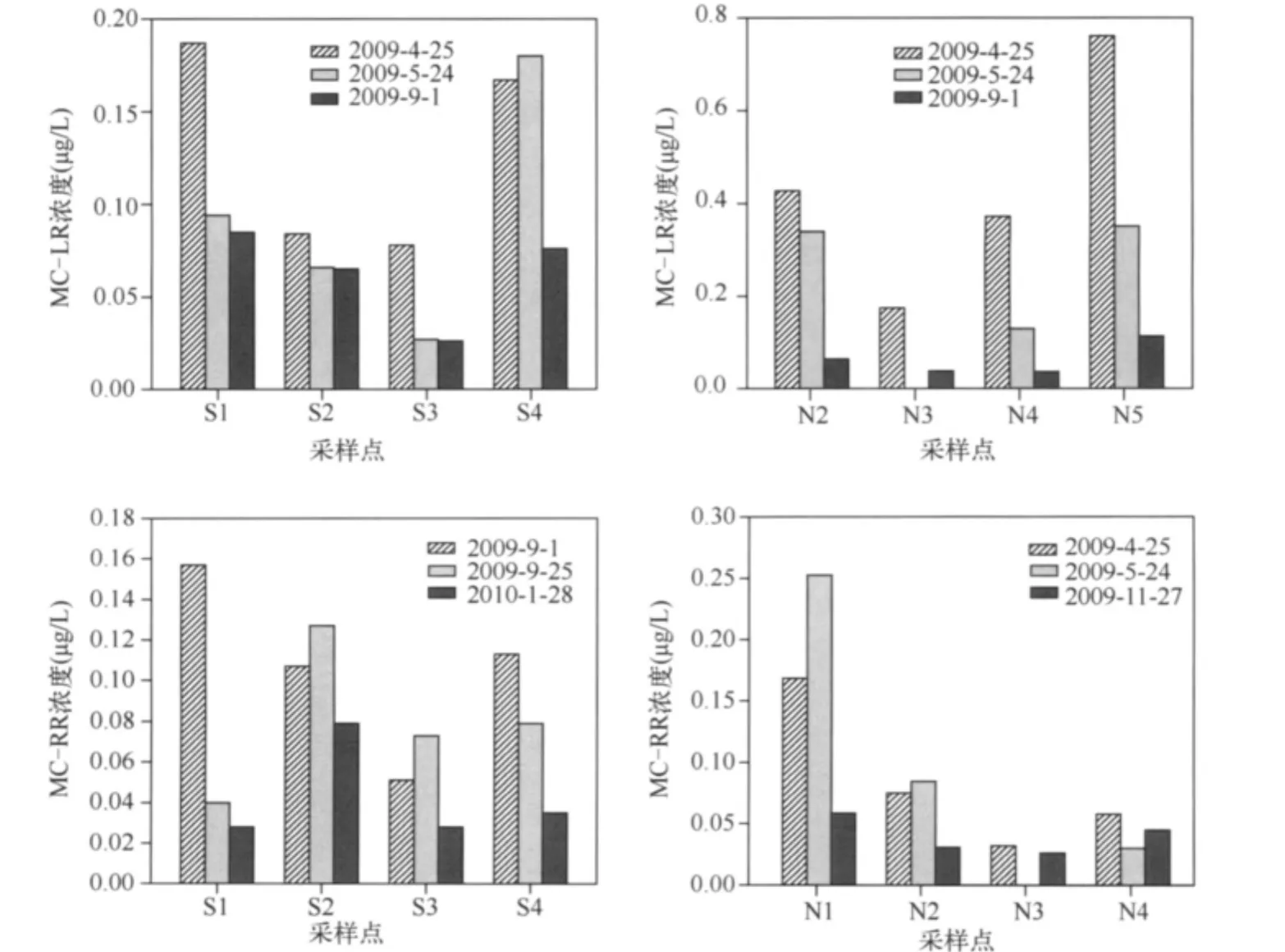
圖3 MCs在太湖的分布狀況Fig.3 The distribution of MCs in Lake Taihu
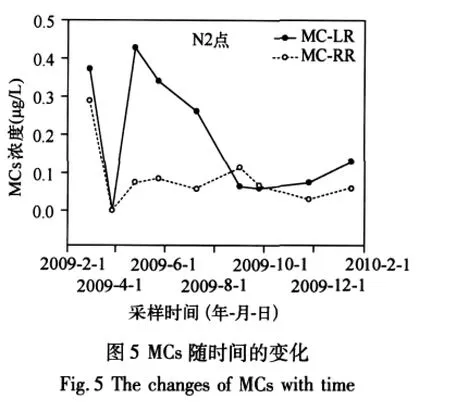
2.1.3 MCs的周年變化 冬季MCs濃度較大,各點(diǎn)的MCs濃度從2009年2月至4月呈增加趨勢;從2009年4月至7月濃度較高但呈減小的趨勢,從2009年11月至2010年4月呈增加趨勢(圖5).藍(lán)藻的數(shù)量從4、5月份開始增加,同時伴隨著藻的死亡,向水體中分泌藻毒素,但是自然水體中微囊藻毒素的去除速度也很快[11],且降水增多,在一定程度上對藻毒素具有稀釋的作用.所以在7月份藍(lán)藻暴發(fā)式生長之前毒素濃度仍然呈增加趨勢.水量對水體中MCs的影響占主要地位.10月份之后藻的密度開始下降,同時太湖流域降水減少,水量的減少對毒素起到一個濃縮的效果.另外,冬季微生物以及水生生物的活性降低,對藻毒素的降解作用減小.有研究表明在自然環(huán)境中底泥沉積物中細(xì)菌對微囊藻毒素的生物降解起到關(guān)鍵的作用[12];淺水湖泊濁度高,光降解的貢獻(xiàn)很小,因此毒素以積累為主,所以到四月為止毒素仍然呈增加的趨勢.從2009年7月至11月,MC-LR濃度緩慢增加,但是MC-RR濃度有一個明顯先增加后減小的過程,并且在8、9月份出現(xiàn)MC-RR濃度大于MC-LR的現(xiàn)象.MCs是一類受產(chǎn)毒基因調(diào)控,在胞內(nèi)代謝產(chǎn)生的毒素.有研究表明在與其它藻類共生的情況下產(chǎn)毒微囊藻更有競爭優(yōu)勢[13],在藍(lán)藻暴發(fā)期微囊藻產(chǎn)毒基因的豐度顯著增加[14-15],因此處于暴發(fā)期的藍(lán)藻可能合成更多的毒素.但微囊藻毒素主要集中在細(xì)胞的類囊體核區(qū),很少存在于細(xì)胞壁和峭壁上[16],只有當(dāng)藻細(xì)胞死亡、破裂時才會釋放到水體中[17].但是即使藻已進(jìn)入穩(wěn)定期或達(dá)到很大的密度也可能只有很少部分的藻細(xì)胞死亡[18-20].自然水體中,微囊藻毒素一般在藍(lán)藻大量衰亡的季節(jié)才會大量釋放到水柱中[11],因此MCs濃度的增加在藍(lán)藻暴發(fā)之后.微囊藻水華發(fā)展過程中,存在多種基因型共存和微囊藻不同基因型的演替[21],可能在藍(lán)藻暴發(fā)前期合成MC-RR的基因種類較多后期較少,或是受其它環(huán)境因子(如光照、溫度、氮、磷等)的影響,出現(xiàn)MC-RR濃度先增加后減小的現(xiàn)象.迄今為止,關(guān)于產(chǎn)生不同藻毒素的原因(是由基因種類決定還是受環(huán)境因子調(diào)控,或是兩者共同作用)鮮有報道.要解釋這種現(xiàn)象,還需要進(jìn)一步研究.
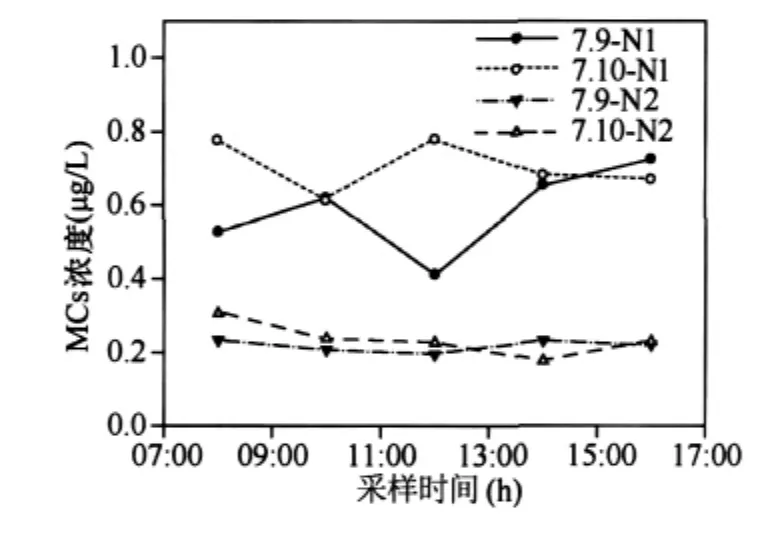
圖6 MCs在一天之內(nèi)的變化Fig.6 The changes of MCs within one day
2.1.4 MCs一天之內(nèi)的變化 對7月份集中采樣的數(shù)據(jù)分析發(fā)現(xiàn)MCs的濃度比較穩(wěn)定,波動較小,一天之內(nèi)的變化規(guī)律不明顯(圖6).MCs主要貯存在細(xì)胞內(nèi),細(xì)胞死亡后毒素才會釋放到水體中.實(shí)驗室研究發(fā)現(xiàn),處于對數(shù)生長期的微囊藻培養(yǎng)物中至多能釋放10%-20%的毒素到培養(yǎng)基中;進(jìn)入穩(wěn)定期時,隨著藻死亡數(shù)量的增加,培養(yǎng)基中毒素的比例會有所增加.但是在藍(lán)藻的生長周期里,藻的死亡數(shù)量只占很少部分[18-20].因此,在藍(lán)藻生長周期里,釋放到水體中的毒素含量是有限的.在自然環(huán)境中,微囊藻毒素的大量釋放一般發(fā)生在水華衰亡的時期.因此在水華暴發(fā)期,一天之內(nèi)釋放到水柱中的藻毒素的量很小且相對較穩(wěn)定.并且,在藍(lán)藻暴發(fā)期,藻密度較大,水體透明度較低,光降解的作用非常有限.底泥中細(xì)菌可能在自然環(huán)境藻毒素的降解過程中起著關(guān)鍵的作用[12],源于底泥中的細(xì)菌對水柱中毒素的降解與水柱中藻毒素的釋放維持著動態(tài)平衡.
2.2 MCs與環(huán)境因子的相關(guān)性分析
氮是藻類生長的必需營養(yǎng)元素,同時氮作為一種必需的元素參與藻毒素的合成[22],因而氮缺乏(<12.53mg/L)不僅限制藻的生長還抑制藻毒素的分泌[23].本研究水體中TN的均值為3.04mg/L,濃度較低,可能抑制藻毒素的分泌.全年相關(guān)性分析結(jié)果(表1)也表明水中溶解性MC-LR與TN呈極顯著正相關(guān),但是MC-RR與TN無顯著相關(guān)性.不同月份MCs與TN相關(guān)性分析表明,大部分時間MC-LR、MC-RR均與TN呈極顯著正相關(guān);MC-LR與TN相關(guān)性偶爾有所變動;MC-RR與TN在7月份藍(lán)藻暴發(fā)到9月份沒有顯著相關(guān)性(表2),這與MC-RR在此期間濃度增加有關(guān)系.總體來說,MCs與TN具有很好的相關(guān)性,這與之前的報道一致[24-25].全年相關(guān)性分析結(jié)果(表1)表明TP與水中溶解性MC-LR呈極顯著正相關(guān),與MC-RR無顯著相關(guān)性.而TP與MC-LR和MC-RR在所有月份均無顯著相關(guān)性.因而TP與MC-LR在全年表現(xiàn)出的極顯著正相關(guān)性可能由二者的季節(jié)變動存在一致性導(dǎo)致的,二者沒有直接的關(guān)系.關(guān)于TP對MCs的影響至今也沒有一致的研究結(jié)論.Sivonen[19]認(rèn)為當(dāng)磷的濃度在0.1-0.4mg/L范圍內(nèi)MCs含量與磷濃度呈正相關(guān),但更高濃度的磷與MCs濃度相關(guān)性不明顯.Utkilen等[26]認(rèn)為磷對微囊藻毒素的產(chǎn)生沒有影響.也有研究表明微囊藻的產(chǎn)毒能力隨著磷濃度的減小而增加[27-28].由于TN/TP波動較大,在1-48之間,MCs與TN/TP的相關(guān)性不顯著.全年監(jiān)測的相關(guān)性分析結(jié)果與不同月份的基本一致.
除藍(lán)藻暴發(fā)期,各個月份的相關(guān)性分析表明,水中溶解性MCs與SS、UV254、TOC、藍(lán)藻密度、水溫均無顯著相關(guān)性(表2).但是全年的相關(guān)性結(jié)果顯示TOC、藍(lán)藻密度、水溫與MC-LR具有統(tǒng)計學(xué)意義上的負(fù)相關(guān)性,與MC-RR無顯著相關(guān)性(表1).因為水中溶解性MC-LR濃度表現(xiàn)為在夏秋兩季濃度較低,春冬兩季濃度較高,MC-RR無此規(guī)律;藍(lán)藻密度、水溫正好相反;同時由于溫度較高、光照強(qiáng)烈、生物活動劇烈導(dǎo)致TOC含量升高[29],所以TOC含量與溫度及藍(lán)藻密度有很好的一致性,即均與MC-LR濃度表現(xiàn)出季節(jié)波動的負(fù)相關(guān)性.藍(lán)藻暴發(fā)期的相關(guān)性分析結(jié)果則表明UV254、TOC、SS、藍(lán)藻數(shù)量與MC-LR呈正相關(guān),與MC-RR無顯著相關(guān)性(表3).MCs主要由藍(lán)藻細(xì)胞死亡破裂后釋放到水體中的[17],因此與藍(lán)藻死亡后水體中的TOC含量及UV254值呈正相關(guān),可能藻細(xì)胞中MCs種類以MC-LR為主[21],所以表現(xiàn)出與MC-LR極顯著相關(guān),與MC-RR不顯著相關(guān).可能由于當(dāng)前死亡藍(lán)藻數(shù)量與藍(lán)藻數(shù)量相當(dāng),并且以生產(chǎn)MC-LR為主[21],所以MC-LR與藍(lán)藻數(shù)量呈極顯著正相關(guān).同時,由于處于藍(lán)藻暴發(fā)期,SS的主要組成部分為藍(lán)藻細(xì)胞,所以MC-LR也與SS呈極顯著正相關(guān).
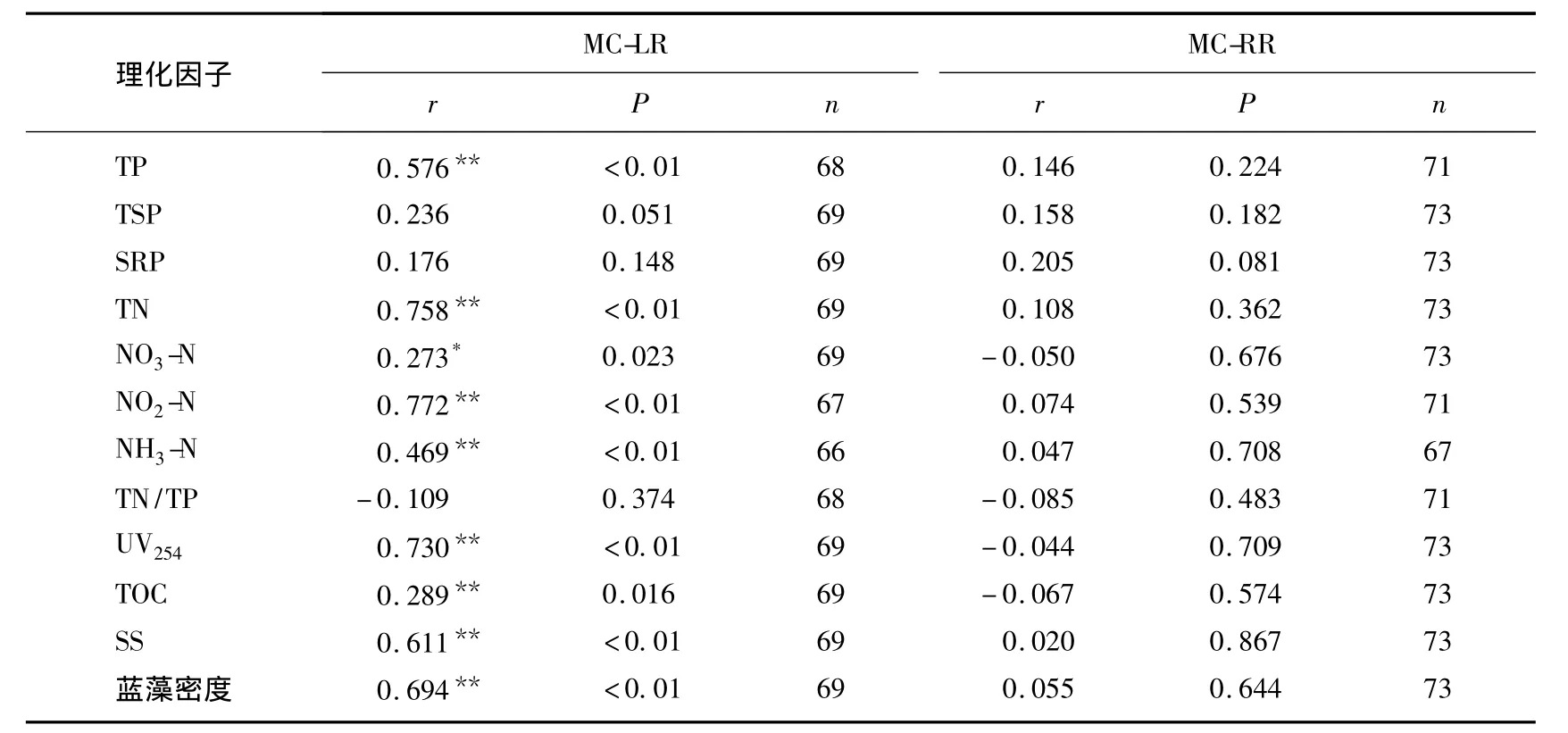
表1 全年MCs與各理化因子相關(guān)性分析Tab.1 Correlation analysis between microcystin and physicochemical factors of the whole year
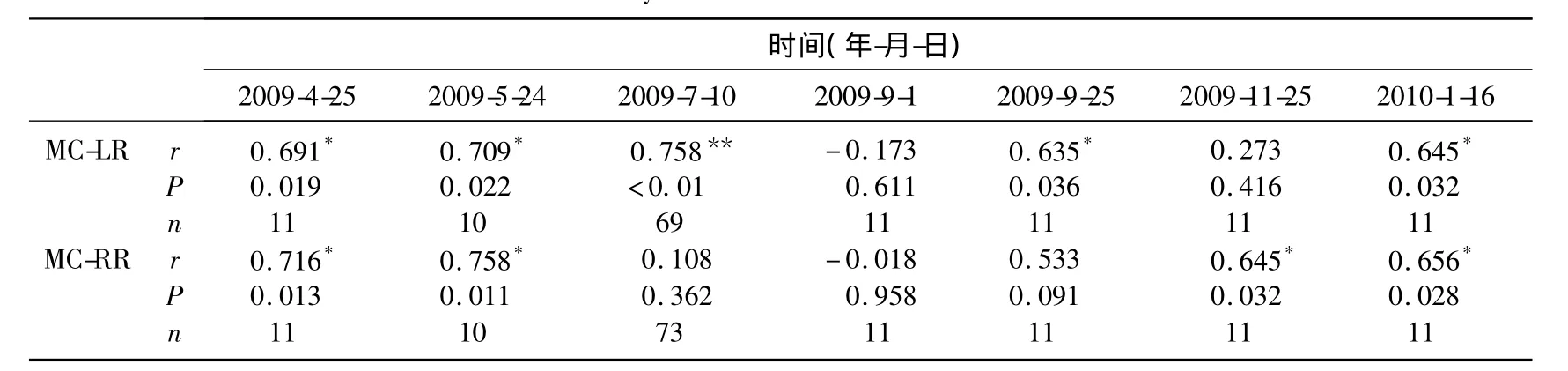
表2 不同時期MCs與TN的相關(guān)性分析Tab.2 Correlation analysis between MCs and TN at different times
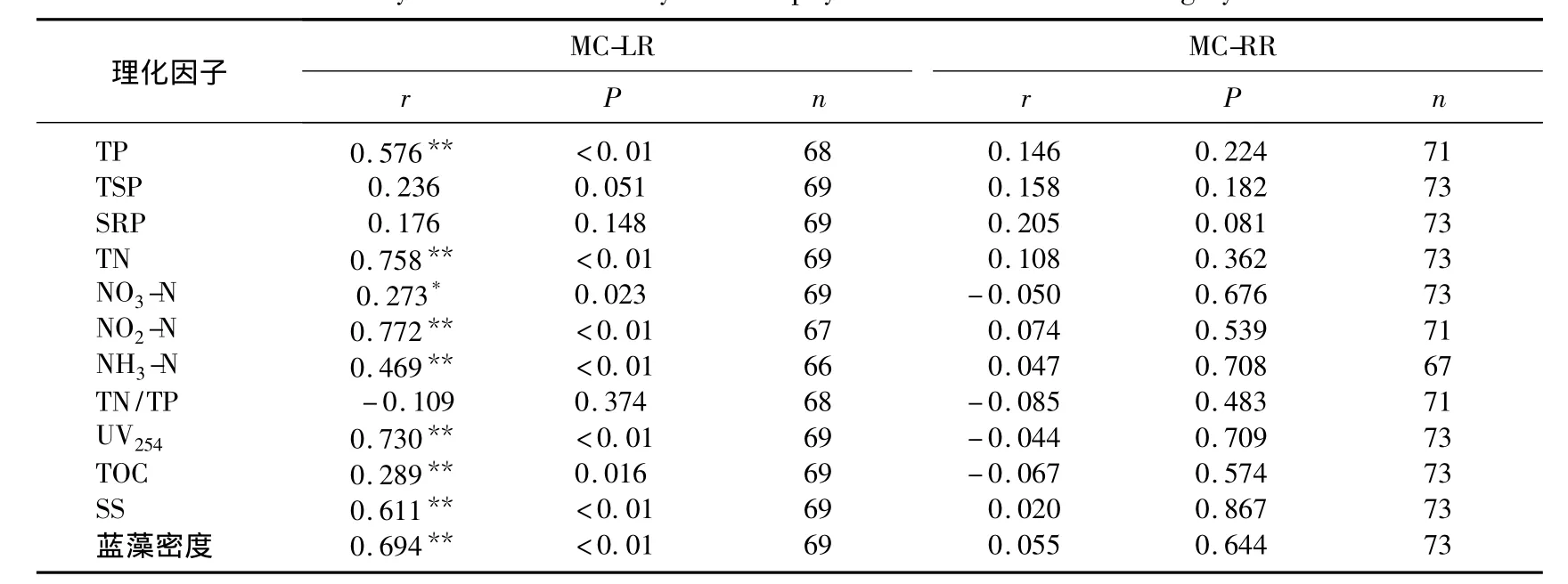
表3 藍(lán)藻暴發(fā)期MCs與各理化因子相關(guān)性分析Tab.3 Correlation analysis between microcystin and physicochemical factors during cyanobacteria blooms
2.2 MCs與環(huán)境因子的回歸分析
全年水中溶解性MCs逐步回歸分析表明,對于MC-LR最好的回歸模型只有TN和SRP進(jìn)入,其R2為0.362,擬合度較差;而MC-RR只有NO2-N進(jìn)入模型,其R2為0.332,擬合度也較差.以現(xiàn)有的環(huán)境因子難以對水體中溶解性MCs的濃度進(jìn)行預(yù)測,有研究表明水體中溶解態(tài)MCs的時空分布受風(fēng)向、風(fēng)速的影響較大[30],增加風(fēng)向、風(fēng)速及水量等物理因子可能能夠預(yù)測全年水體中溶解性MCs的濃度.
3 結(jié)論
對太湖全湖9個點(diǎn)位藻毒素為期一年的監(jiān)測發(fā)現(xiàn)表層與底層藻毒素含量幾乎沒有差別,MC-LR與MCRR的濃度大小則表現(xiàn)為除8、9月份以外MC-LR均大于MC-RR,藻毒素在各點(diǎn)分布不均,在時間上表現(xiàn)為冬季濃度較高,在夏季,一天之內(nèi)藻毒素濃度沒有明顯的變化規(guī)律.北部N1和N5點(diǎn)濃度在一年內(nèi)變化較大,在藍(lán)藻暴發(fā)后藻毒素濃度明顯增加,而其他點(diǎn)均未發(fā)現(xiàn)有此規(guī)律.通過對藻毒素與一些環(huán)境因子的相關(guān)性分析發(fā)現(xiàn),這些環(huán)境因子共同作用影響藍(lán)藻產(chǎn)毒量和藻毒素在環(huán)境中的歸趨從而影響藻毒素的含量,是一個非常復(fù)雜的過程.其中風(fēng)向、風(fēng)速以及水量等物理因素可能對藻毒素的濃度及其時空分布起著非常重要的作用.
致謝:感謝南京大學(xué)水處理與水環(huán)境修復(fù)教育部工程研究中心在水樣采集和樣品分析過程中提供的大力協(xié)助.
[1]Falconer IR.Tumor promotion and liver injury caused by oral consumption of cyanobacteria.Environ Toxic Water,1991,6(2):177-184.
[2]Dowson RD.The toxicity of microcystins.Toxicon,1998,36(7):953-962.
[3]Baldia SF,Conaco MCG,Nishijima T et al.Microcystin production during algal bloom occurrence in Laguna de Bay,the Philippines.Fisheries Sci,2003,69:110-116.
[4]Watanabe MF.Toxic Microcystis.Boca Raton:CRC Press,1996:35-36.
[5]Figueiredo DR,Azeiteiro UM,Esteves SM et al.Microcystin-producing bloom-a serious global public health issue.Ecotox Environ Safe,2004,59(2):151-163.
[6]Vasconcelos VM,Carmichael WW,Croll B et al.Hepatotoxic microcystin diversity in cyanobacterial blooms collected in portuguese freshwaters.Water Res,1996,30(10):2377-2389.
[7]國家環(huán)境保護(hù)總局主編.水和廢水監(jiān)測分析方法(第四版).北京:中國環(huán)境科學(xué)出版社,2002:247-246,254-279,106-108,70-671.
[8]劉約權(quán)主編.現(xiàn)代儀器分析.北京:高等教育出版社,2001:122-159.
[9]全國漁業(yè)自然資源調(diào)查和淡水專業(yè)組.內(nèi)陸水域漁業(yè)自然資源調(diào)查試行規(guī)范.全國漁業(yè)自然資源調(diào)查和淡水專業(yè)組,1980:12-15.
[10]毛新偉,徐 楓,徐 彬等.水質(zhì)及富營養(yǎng)化變化趨勢分析.水資源保護(hù),2009,25(1):48-51.
[11]宋立榮,陳 偉.水華藍(lán)藻產(chǎn)毒的生物學(xué)機(jī)制及毒素的環(huán)境歸趨研究進(jìn)展.湖泊科學(xué),2009,21(6):749-757.
[12]Chen W,Song LR,Peng L et al.Reduction in microcystin concentrations in large and shallow lakes:water andsedimentinterface contributions.Water Res,2008,42:763-773.
[13]Briand E,Yéprémian C,Humbert JF et al.Competition between microcystin-and non-microcystin-producing Planktothrix agardhii(cyanobacteria)strains under different environmental conditions.Environ Microbiol,2008,10(12):3337-3348.
[14]Ye WJ,Liu XL,Tan J et al.Diversity and dynamics of microcystin-Producing cyanobacteria in China's third largest lake,Lake Taihu.Harmful Algae,2009,8:637-644.
[15]Kim SG,Joung SH,Ahn CY et al.Annual variation of Microcystis genotypes and their potential toxicity in water and sediment from a eutrophic reservoir.FEMS Microbiol Ecol,2010,74:93-102.
[16]Shi L,Carmichael WW,Miller I.Immuno-gold localization of hepatotoxins in cyanobacterial cells.Arch Microbiol,1995,163:7-15.
[17]Caemichael WW.The toxicity of cyanobacteria.Sci Am,1994,270(1):64-70.
[18]Young FM,Thomson C,Metcalf JS et al.Immunogold localisation of microcystins in cryosectioned cells of Microcystis.J Struct Biol,2005,151:208-214.
[19]Sivonen K.Effects of light,temperature,nitrate,orthophosphate,and bacteria on growth of and hepatotoxin production by Oscillatoria agardhii strains.Appl Environ Microbiol,1990,56:2658-2666.
[20]Negri AP,Jones GJ,Blackburn SI et al.Effect of culture and bloom development and of sample storage on paralytic shellfish poisons in the cyanobacterium Anabaena circinalis.J Phycol,1997,33:26-35.
[21]Kardinaal WEA,Janse I,Kamst-van Agterveld M et al.Microcystis genotype succession in relation to microcystin concentrations in freshwater lakes.Aquat Microb Ecol,2007,48:1-12.
[22]Botes DP,Wessels PL,Kruger H et al.Structural studies on cyanoginosins-LR,-YR,-YA and-YM,peptide toxins from Microcystis aeruginosa.J Chem Soc Perk T 1,1985,1:2747-2748.
[23]代瑞華,劉會娟,曲久輝等.氮磷限制對銅綠微囊藻生長和產(chǎn)毒的影響.環(huán)境科學(xué)學(xué)報,2008,28(9):1739-1744.
[24]張杭君,張建英,陳英旭等.微囊藻毒素含量與自然水體環(huán)境影響因子的相關(guān)性.環(huán)境科學(xué),2006,27(10):1969-1973.
[25]連 民,陳傳瑋,俞順章等.淀山湖夏季微囊藻毒素分布狀況及其影響因素.中國環(huán)境科學(xué),2000,20(4):323-327.
[26]Utkilen H,Gjolme N.Iron-stimulated toxin production in microcystis aeruginosa.Appl Environ Microbiol,1995,61(2):797-800.
[27]Oh HM,Lee SJ,Jang MH et al.Microcystin production of microcystis aeruginosa in a Phosphorus-limited chemostat.Appl Environ Microbiol,2000,66(1):176-179.
[28]張 瑋,林一群,郭定芳等.不同氮、磷濃度對銅綠微囊藻生產(chǎn)、光合及產(chǎn)毒的影響.水生生物學(xué)報,2006,30(3):318-322.
[29]江志堅,黃小平,張景平.大亞灣海水中總有機(jī)碳的時空分布及其影響因素.海洋學(xué)報,2009,31(1):91-98.
[30]許秋瑾,高 光,陳偉民.太湖微囊藻毒素與湖泊物理因素之間的關(guān)系.中國環(huán)境監(jiān)測,2004,20(1):12-15.
Characteristic of microcystin distributions and its relationships with environmental factors in Lake Taihu
WANG Jingjie,YANG Jia,XIAN Qiming,QIAN Xin,LI Zhengkui,GENG Jinju & SUN Cheng
(State Key Laboratory of Pollution Control and Resource Reuse,School of the Environment,Nanjing University,Nanjing 210093,P.R.China)
In this paper,the high performance liquid chromatography(HPLC)was used to monitor the spatial and temporal distribution of dissolved Microcystin-LR and Microcystin-RR in Lake Taihu,and its relationship with physiochemical factors of the lake were also discussed.The results showed that the concentrations of microcystin(MCs)were not different between surface and bottom water in a day,and the highest concentration of microcystins occurred in winter.Concentrations of the MCs changed much during one year in the sampling sites N5 and N1 which located in Zhushan Bay and the western part of Lake Taihu,respectively.Generally,the concentration of the MC-LR was higher than MC-RR for the year in average while the opposite results appeared in individual month of August and September.Correlation analysis showed that MC-LR was significantly positive correlated with TN,but did not show correlation with TP.During the period of water bloom of cyanobacteria,levels of the MC-LR were significantly positive correlated with TP,TN,UV254,SS,total organic carbon and biomass of cyanobacteria,whereas MC-RR was not significant correlated with all the environmental factors in the statistical test.
Microcystin;Lake Taihu;environmental factors;correlation
* 國家“973”計劃項目(2008CB418003)和國家水體污染控制與治理科技重大專項(008ZX07421-002)項目聯(lián)合資助.2010-08-16收稿;2010-12-21收修改稿.王經(jīng)結(jié),女,1985年生,碩士研究生;E-mail:wangjingjie66@gmail.com.
** 通訊作者;E-mail:xianqm@nju.edu.cn.
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學(xué)學(xué)報(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀(jì)智能(英語備考)(2019年12期)2020-01-13 06:07:18
瘋狂英語·新策略(2019年9期)2019-10-17 01:51:34
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
濰坊學(xué)院學(xué)報(2017年2期)2017-04-20 08:44:31
中國環(huán)境監(jiān)察(2016年5期)2016-10-24 05:25:52
中國商論(2016年33期)2016-03-01 01:59:38
