植物幾丁質酶純化測定及應用研究進展
2011-10-09 02:47:08楊海霞鄧建軍趙廣華
食品工業科技 2011年6期
關鍵詞:植物
楊海霞,鄧建軍,2,張 建,3,趙廣華,*
(1.中國農業大學食品科學與營養工程學院,北京100083;2.西北大學食品科學與工程系,陜西西安710069;3.石河子大學食品學院,新疆石河子832000)
植物幾丁質酶純化測定及應用研究進展
楊海霞1,鄧建軍1,2,張 建1,3,趙廣華1,*
(1.中國農業大學食品科學與營養工程學院,北京100083;2.西北大學食品科學與工程系,陜西西安710069;3.石河子大學食品學院,新疆石河子832000)
植物幾丁質酶是植物體中能夠水解幾丁質多聚體的一種致病性相關蛋白(Pathogenesis-related proteins)。近年來對于幾丁質酶的研究報道中,大量新型的植物幾丁質酶被分離純化,并建立了不同的酶活測定方法,在幾丁質酶的結構及分類方面也逐步有了系統的研究。從幾丁質酶的結構及分類,分離純化以及已建立的酶活測定方法等方面取得的新進展進行了綜述,并展望了幾丁質酶在農業、食品生產及藥用領域的應用前景。
植物,幾丁質酶,分類,純化,酶活測定
植物在受到致病性微生物侵害時,會誘導表達大量不同基因編碼的蛋白質——致病性相關蛋白(Pathogenesis-related proteins)來保護植株免受病原菌的侵害[1-4]。而在這些誘導的致病性相關蛋白質中,幾丁質酶(Chitinase,E.C.3.2.1.14)被證明參與了寄主植物與真菌病原菌的相互作用[5-7]。幾丁質酶又稱殼多糖酶,能夠水解幾丁質內部的β-1,4-糖苷鍵,生產N-乙酰葡萄糖胺寡聚體。它廣泛存在于植物體中,人們已從植物的根、莖、葉、果實以及種子中分離純化得到多種幾丁質酶[8-10],并且發現幾丁質酶可以通過破壞菌絲頂端及其側壁抑制真菌生長,從而達到抗真菌感染的目的;還可以通過釋放寡糖誘導植物抗霉素的產生。抑菌實驗結果表明當幾丁質酶與β-1,3-葡聚糖酶共同作用時,抑菌效果會大大提高[7]。由于幾丁質酶具有很強的抗真菌活性,近年來,幾丁質酶已經成為工業、農業、生物技術以及食品等領域研究的熱點。本文就植物來源的幾丁質酶在結構、分類、分離純化方法、酶活測定方法以及應用前景等方面進行了綜述,為今后植物源幾丁質酶的開發和應用提供技術參考。
1 幾丁質酶的結構及分類
1.1 幾丁質酶的結構
近年來,對多種植物的幾丁質酶基因進行了克隆,研究發現植物幾丁質酶的核苷酸序列與所編碼的氨基酸序列在結構上存在一些規律性的變化,植物幾丁質酶前體一般都含有N-端信號區(Signal peptide)、催化區(Catalytic domain)和 C-端延伸區(C-terminal extension),有的在N-端信號區之后有富含半胱氨酸的幾丁質結合區域(Chitin binding domain,CBD),它與主結構區之間有一段可變交聯區
(Flexible hinge region)[11](如圖 1)。N-端信號區一般由20多個氨基酸組成,為疏水性,作用是調節酶的分泌作用,引導幾丁質酶通過細胞膜運輸到內質網后被信號肽酶裂解掉。有的胞內幾丁質酶是沒有N-端信號區的[12],不同幾丁質酶的N-端信號區也是不同的。CBD是一段由40個氨基酸組成的富含半胱氨酸的高度保守結構[3],6個半胱氨酸殘基位置基本固定,之間形成二硫鍵,可能是維持三維結構和蛋白之間的相互作用。可變交聯區富含甘氨酸和脯氨酸,不同的幾丁質酶的交聯區長度差異很大。催化區的作用是水解底物,序列對比顯示催化區有兩段高度保守的區域,其中SxGG是結合底物部位,DxxDxDxE是催化水解底物區域[13],需要說明的是催化區的底物結合部位和CBD是兩個獨立的區域,研究表明CBD不能夠增加幾丁質水解速率,只能夠增加幾丁質酶的結合能力[14-15]。C-端延伸區的作用是用來引導幾丁質酶進入植物液泡[16]。
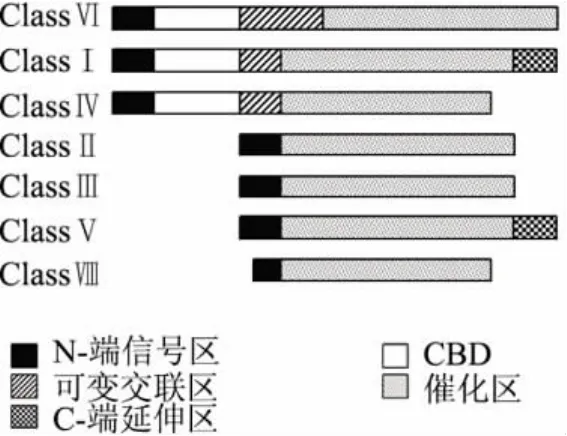
圖1 植物幾丁質酶的結構分類圖
1.2 幾丁質酶的分類
幾丁質酶依據結構特點和氨基酸序列被分為以下幾類[17],如圖1所示:ClassⅠ在 N-端信號區后有幾丁質交聯區(CBD)以及可變交聯區,連接著催化區,一般含有約300個氨基酸,此類幾丁質酶呈堿性,大部分存在于液泡內;ClassⅡ的主要氨基酸序列和ClassⅠ有很強的同源性,只是缺少CBD和可變交聯區,此類幾丁質酶多數呈酸性,存在于胞質外;ClassⅢ的結構和前兩類植物幾丁質酶的結構很不相同,但是和細菌、真菌幾丁質酶具有較高的同源性,同樣不存在CBD區[18],它是胞外水解酶,在酶活性方面和溶菌酶有較大的相似性;ClassⅣ和ClassⅠ具有序列同源性,也同樣含有CBD和保守的主要結構,ClassⅣ和ClassⅠ可用血清學方法來區分,ClassⅣ的成熟蛋白質基本含有241~255個氨基酸,分子量比ClassⅠ略小是由于有部分片段缺失[3]。Melchers el從煙草分離到一種新的幾丁質酶,此酶的氨基酸序列與幾種細菌的幾丁質外切酶相似,但與已報道的植物幾丁質酶不同,故將其定義為ClassesⅤ[10]。目前在煙草、蕁麻、甜菜、豆科植物以及裸子植物中均已發現 ClassesⅤ幾丁質酶[19-20]。ClassⅥ幾丁質酶的N端序列含有兩個重復排列的可變交聯區域,而ClassⅦ則缺失CBD區域,與ClassⅣ有較高的同源性,已從小麥、棉花中提取得到此類酶[21-22]。
如果依據糖基水解酶系統分類,植物幾丁質酶可以被分為兩個家族,即Family 18和Family 19。所有的真菌、動物、細菌幾丁質酶以及植物ClassⅢ、Ⅳ幾丁質酶均為Family 18,植物 ClassⅠ、Ⅱ、Ⅴ、Ⅵ、Ⅶ為Family 19[23]。二者在氨基酸序列、三維結構以及分子機制等方面的差異表明它們分別起源于不同的祖先[21]。
2 植物幾丁質酶的分離純化方法
植物幾丁質酶的分子量介于25~35kD之間。在以幾丁質為底物時,最適反應pH一般低于7,其等電點介于3~10之間,植物幾丁質酶有著較好的熱穩定性(有的在60℃仍很穩定),且抗蛋白酶降解[24]。
植物幾丁質酶可以通過不同的方法分離純化,主要有硫酸銨鹽析、離子交換層析、疏水作用層析、凝膠過濾層析、幾丁質-親和層析以及電泳等方法。分離純化過程最好在低溫(4℃)下進行。幾丁質酶的酶活會隨著純化步驟的增加有所降低。提取植物幾丁質酶一般按如下方法進行:將植物原料用適當pH的緩沖液勻漿,攪拌后過濾,得到的濾液離心去除不溶性淀粉及雜質,上清液中加入適當濃度的硫酸銨,使幾丁質酶沉淀,靜置一段時間,離心取沉淀,用緩沖液重新溶解成溶液狀態,透析去除鹽分等雜質,即得到幾丁質酶的粗提液。純化植物幾丁質酶的方法有很多,依據不同幾丁質酶的生物化學性質選擇合適的純化方法。Bloch and Burger利用幾丁質與幾丁質酶結合的原理,將酶粗提液過幾丁質親和柱,收集洗脫液即為純化的幾丁質酶[25]。Takeshi Watanabe等人通過離子交換層析和凝膠過濾層析對幾丁質酶進行純化[26],純化過程中要選擇適當的pH、緩沖液、洗脫鹽濃度、流速等條件,也可利用電泳(Polyacrylamide Gel Electrophoresis)的方法制備得到純化的幾丁質酶[27]。
3 植物幾丁質酶的酶活測定方法
植物幾丁質酶主要是水解幾丁質多聚體中的β-1,4-糖苷鍵,幾丁質酶的水解活性主要有三種方式,即內切幾丁質酶(Endochitinase)、外切幾丁質酶(Exochitinase)以及 N-乙酰氨基己糖苷酶(GlcNAcase)[13,27]。內切幾丁質酶水解位點為同聚物任意中間位點的β-1,4-糖苷鍵,生成可溶的、低分子量的 N-乙酰氨基葡糖多聚體,如:chitotetraose、chitotriose以及diacetylchitobiose,其作用底物大多為四糖以上的低聚糖,而且酶與底物親和性也隨著底物聚合度提高而增大[28];外切幾丁質酶則作用于非還原端口依次切下單糖,它生成的產物只有diacetylchitobiose,沒有單糖或低聚糖;N-乙酰氨基己糖苷酶利用外切形式將diacetylchitobiose和幾丁質多聚體裂解成N-乙酰氨基葡糖單體[13]。離體實驗表明,幾丁質酶的活性底物還有其它物質;有的幾丁質酶具有溶菌酶活性可作用于細菌肽聚糖、N-乙酰胞壁酸和N-乙酰葡萄糖之間的β-1,4-糖苷鍵;有的還能催化轉糖基反應[29]。一般幾丁質酶的內切作用和外切作用是同時進行的[12]。從許多植物包括大麥、玉米、煙草、馬鈴薯、大豆和大米中提取的幾丁質酶都屬于內切幾丁質酶,主要產物為低聚糖[30]。
3.1 還原糖測定方法
目前,關于檢測幾丁質酶酶活的方法報道很多。最常用的方法是用膠體幾丁質(Colloidal chitin)做底物,通過檢測酶解后的產物還原糖進行酶活測定。膠體幾丁質制備方法如Imanaka T等人所述[31],選擇合適的酶解條件進行酶解反應后,生成產物還原糖利用Imato和Yagishita的方法測定,1U等于釋放1μg 還原糖所需的酶量[32]。Nitoda T.K.等人發現用膠狀幾丁質的顆粒均一大小的粉末作底物,靈敏度比用膠狀幾丁質上清液高4倍[33]。幾丁質的可溶性衍生物乙二醇幾丁質(Glycolchitin)以及脫乙酰殼聚糖(Chitosan)被用作十分重要的測定酶活底物,酶解后通過紫外吸光值的測定反應酶活[34]。還有報道認為用膠狀幾丁質做底物測定還原糖的含量只測出外切幾丁質酶的活性,如要測內切幾丁質酶活性還需加入蝸牛腸酶酶解,再測定還原糖含量[35]。
3.2 生色底物測定方法
為了更好地檢測溶液中幾丁質酶的酶活,生色測定方法被建立并逐漸成熟起來。底物N-acetyl-D- glucosaminide(GlcNAc)、β - D - N,N′-diacetylchitobiose[(GlcNAc)2]以 及 β-D-N,N′,N″-triacetylchitotrios[(GlcNAc)3]可用生色基團p-nitrophenyl(pNP)分別標記為 pNP-GlcNAc,pNP-(GlcNAc)2以及 pNP-(GlcNAc)3。內切幾丁質酶、外切幾丁質酶以及N-乙酰氨基己糖苷酶分別酶解pNP-(GlcNAc)3、pNP-(GlcNAc)2以及 pNP-GlcNAc后釋放pNP,通過熒光測定反應酶活。Nagel R.等人提出利用pNP-GlcNAc作底物比固體幾丁質靈敏,幾丁質酶對pNP-GlcNAc的水解速率是固體幾丁質的三倍,1U等于釋放1nmol pNP所用的酶量[36]。同樣,底物被4-methylumbelliferyl(4-MU)修飾的測定酶活的方法也被建立起來。4-MU-(GlcNAc)3、4-MU-(GlcNAc)2和4-MU-GlcNAc酶解后可釋放4-MU,通過熒光(激發波355nm,發射波460nm)分別測定反應內切幾丁質酶、外切幾丁質酶以及N-乙酰氨基己糖苷酶的酶活,1U即釋放1nmol 4-MU所用的酶量。
3.3 染料底物測定方法
有些學者利用染料修飾膠狀幾丁質,利用酶解釋放的染料具有特異吸光值來測定幾丁質酶的活性。早在1964年Hackman和Goldberg就提出用有色幾丁質衍生物Chitin Azure作底物,酶解后通過550nm處測定吸光值反映幾丁質酶酶活,1U酶活被定義為吸光值增加0.001所需的酶量[37]。M.Go′mez Ram?′rez提出了一種快速靈敏的方法,利用Remazol Brilliant Blue RR(RBB)染色的膠狀幾丁質作底物,培養酶解后通過在595nm下測定吸光值反映幾丁質酶的活性[38]。Ostazin Brilliant Red用作標記幾丁質底物,酶解后釋放的被染料標記的GlcNAc在530nm測 定 吸 光 值[39]。Mojtaba Mamarabadi將 底 物 用Remazol Brilliant Violet(RBV)修飾,酶解后在550nm下測定吸光值反映酶活[40]。
4 植物幾丁質酶在食品中的應用前景
近些年來,人們從幾丁質酶的酶學特性、抗菌機理以及分子克隆方面已經有了系統的研究,但是對于幾丁質酶防御反應的機理還有待闡明;此外,目前學者的興趣主要集中于尋找或重組有更好動力學特性的新型幾丁質酶,以及對幾丁質酶活性位點殘基的識別[27]。由于植物幾丁質酶的重要作用,其在食品及藥物方面有著廣泛的應用。
4.1 食品添加劑
由于幾丁質酶具有良好的抑菌活性[7],所以在食品生產中,幾丁質酶與其它抗微生物酶(葡萄糖氧化酶和乳過氧化物酶)或與傳統的防腐措施(如山梨酸鹽、乙醇、溫度和低pH)聯合使用,可以提高食品微生物的安全性,在糕點或飲料等食品中適當添加,可防止微生物的繁殖,起到防腐作用。
另外,幾丁質對動植物而言是一種較難吸收利用的氮素來源,幾丁質酶可將食品中的幾丁質酶解,作用后所產生的N-乙酰幾丁寡糖和殼聚幾丁寡糖都具有清爽的甜味,吸濕性和保濕性好,而且在水中溶解度比單糖低,有助于調整食品的水活性,增進保水性,兼具調味和改良食品質構的功能。
4.2 降解產物的利用
幾丁質酶的降解產物寡聚糖、葡糖胺以及N-乙酰氨基葡糖在醫藥及食品方面有著廣闊的應用前景,我國漁業食品工業生產中的廢棄物中含有大量幾丁質,而傳統工藝用酸水解蝦殼等生產幾丁寡糖成本昂貴且對容器腐蝕性大,利用幾丁質酶水解法制備其降解產物,不僅能徹底有效的處理廢殼,減少對環境的污染,還可以變廢為寶,使降解產物得以利用,這種環保、經濟、快速的方法無疑為企業開拓一條新的途徑。
4.3 食品基因工程中的應用
由于幾丁質酶具有顯著的抗菌特性[7],近年來,通過轉基因技術,將水稻、大麥、煙草等幾丁質酶基因轉入水稻、番茄等植物中用于病蟲害防治,效果十分顯著,不僅獲得了高效而廣譜的抗性,還克服了轉基因植物的表達水平低的問題,應用前景十分廣闊。
5 小結
總之,無論從幾丁質資源的開發利用、減輕污染和保護環境的角度來看,還是幾丁質原料及幾丁質酶在農業、工業、食品及藥物中的重要作用,充分研究幾丁質酶,利用幾丁質酶的特性開發新的應用前景將會給社會以及人類帶來巨大幫助。
[1]Kitajima S,Sato F.Plant pathogenesis- related proteins:molecular mechanisms of gene expression and protein function[J].The Journal of Biochemistry,1999,125(1):1-8.
[2]Tyler B M.Entering and breaking:virulence effector proteins of oomycete plant pathogens[J].Cell Microbiology,2009,11(1):13-20.
[3]Collinge D B,Kragh K M,Mikkelsen J D,et al.Plant chitinases[J].Plant Journal,1993,3(1):31-40.
[4]Ellis J G,Rafiqi M,Gan P et al.Recent progress in discovery and functional analysis of effector proteins of fungal and oomycete plant pathogens[J].Current Opinion in Plant Biology,2009,12(4):399-405.
[5]Saito A,Fujii T,Shinya T,et al.The msiK gene,encoding the ATP- hydrolysing component of N,N′- diacetylchitobiose ABC transporters,is essential for induction of chitinase production in Streptomyces coelicolor A3(2) [J].Microbiology,2008,154(Pt 11):3358-3365.
[6]Shinya T,Hanai K,Galis I,et al.Characterization of NtChitIV,a class IV chitinase induced by beta-1,3-,1,6-glucan elicitor from Alternaria alternata 102:Antagonistic effect of salicylic acid and methyl jasmonate on the induction of NtChitIV[J].Biochemical and Biophysical Research Communication,2007,353(2):311-317.
[7]El G A,Wilson C L,Callahan A M.Induction of Chitinase,beta-1,3- Glucanase,and Phenylalanine Ammonia Lyase in Peach Fruit by UV- C Treatment[J].Phytopathology,2003,93(3):349-355.
[8]Graham L S,Sticklen M B.Plant chitinases[J].Canadian Journal of Botany,1994,72:1057-1083.
[9]Mauch F,Mauch-Mani B,Boller T.Antifungal hydrolases in pea tissue:II.Inhibition of fungal growth by combinations of chitinase and beta-1,3-glucanase[J].Plant Physiology,1988,88(3):936-942.
[10]Melchers L S,Apotheker-De G M,van der Knaap J A,et al.A new class of tobacco chitinases homologous to bacterial exochitinases displays antifungal activity[J].Plant Journal,1994,5(4):469-480.
[11]歐陽石文,趙開軍,馮蘭香,等.植物幾丁質酶的研究進展[J].生物工程進展,2001,21:30-34.
[12]Seidl V,Huemer B,Seiboth B,et al.A complete survey of Trichoderma chitinases reveals three distinct subgroups of family 18 chitinases[J].Federation of European Biochemical Societies Journal,2005,272(22):5923-5939.
[13]Duo- Chuan L.Review offungalchitinases[J].Mycopathologia,2006,161(6):345-360.
[14]Kuranda M J,Robbins P W.Chitinase is required for cell separation during growth of Saccharomyces cerevisiae[J].Journal of Biological Chemistry,1991,266(29):19758-19767.
[15]Limon M C,Margolles-Clark E,Benitez T,et al.Addition of substrate-binding domains increases substrate-binding capacity and specific activity of a chitinase from Trichoderma harzianum[J].FEMS Microbiology Letters,2001,198(1):57-63.
[16]Neuhaus J M,Sticher L,Meins F J,et al.A short C-terminal sequence is necessary and sufficient for the targeting of chitinases to the plant vacuole[J].Proceedings of the National Academy of Sciences of the United States of America,1991,88(22):10362 -10366.
[17]Shinshi H,Neuhas J M,Ryals J,et al.Structure of a tobacco endochitinase gene:evidence that different chitinase genes can arise by transposition of sequences encoding a cysteine-rich domain[J].Plant Molecular Biology,1990,14(3):357-368.
[18]Kuo C J,Liao Y C,Yang J H,et al.Cloning and characterization ofan antifungalclass III chitinase from suspension- cultured bamboo(Bambusa oldhamii)cells[J].Journal of Agricultural and Food Chemistry,2008,56(23):11507-11514.
[19]Van Damme E J,Culerrier R,Barre A,et al.A novel family of lectins evolutionarily related to class V chitinases:an example of neofunctionalization in legumes[J].Plant Physiology,2007,144(2):662-672.
[20]Taira T,Hayashi H,Tajiri Y,et al.A plant class V chitinase from a cycad(Cycas revoluta):biochemical characterization,cDNA isolation,and posttranslational modification [J].Glycobiology,2009,19(12):1452-1461.
[21]Henrissat B,Bairoch A.New families in the classification of glycosyl hydrolases based on amino acid sequence similarities[J].Biochemical Journal,1993,293:781-788.
[22]Suzuki K,Taiyoji M,Sugawara N,et al.The third chitinase gene(chiC)of Serratia marcescens 2170 and the relationship of its product to other bacterial chitinases[J].Biochemical Journal,1999,343:587-596.
[23]Powning R F,Irzykiewicz H.Studies on the chitinase system in bean and other seeds[J].Comparative Biochemistry and Physiology,1965,14:127-133.
[24]Bloch R,Burger M M.Purification of wheat germ agglutinin using affinity chromatography on chitin[J].Biochemical and Biophysical Research Communications,1974,58(1):13-19.
[25]Watanabe T,Kobori K,Miyashita K et al.Identification of glutamic acid 204 and aspartic acid 200 in chitinase A1 of Bacillus circulans WL-12 as essential residues for chitinase activity[J].Journal of Biological Chemistry,1993,268(25):18567-18572.
[26]Dahiya N,Tewari R,Hoondal G S.Biotechnological aspects of chitinolytic enzymes:a review[J].Applied Microbiology and Biotechnology,2006,71(6):382-773.
[27]Molano J,Polacheck I,Duran A,et al.An endochitinase from wheat germ[J].Journal of Biological Chemistry,1979,254(11):4901-4907.
[28]Song H K,Suh S W.Refined structure of the chitinase from barley seeds at 2.0 a resolution[J].Acta Crystallographica Section D-biological Crystallography,1996,52(Pt 2):289-298.
[29]Imanaka T,FukuiT,Fujiwara S.Chitinase from Thermococcus kodakaraensis KOD1[J].Methods in Enzymology,2001,330:319-329.
[30]Imoto T,Yagishita K.A simple activity measurement of lysozyme[J].Agricultural and Biological Chemistry,1971,35:1154-1156.
[31]Nitoda T,Kurumatani H,Kanzaki H,et al.Improved bioassay method for Spodoptera litura chitinase inhibitors using a colloidal chitin powder with a uniform particle size as substrate[J].Pesticide Science,1999,55(5):563-565.
[32]Liau C Y,Lin C S.A modified coomassie brilliant blue G 250 staining method for the detection of chitinase activity and molecular weight after polyacrylamide gel electrophoresis[J].Journal of Bioscience and Bioengineering,2008,106(1):111-113.
[33]Boller T,Vogeli U.Vacuolar localization of ethylene-induced chitinase in bean leaves[J].Plant Physiology,1984,74(2):442-444.
[34]Kovacs K,Szakacs G,Pusztahelyi T,et al.Production of chitinolytic enzymes with Trichoderma longibrachiatum IMI 92027 in solid substrate fermentation[J].Applied Biochemistry and Biotechnology,2004,118(1-3):189-204.
[35]Nagel R,Birch R G,Manners J M.Detection of bacterial chitinase activity in transformed plant tumour cells using a specific exochitinase substrate[J].Plant Cell Reports,1990,8(12):729-732.
[36]Hackman R H,Goldberg M.New substrates for use with chitinases[J].Analytical Biochemistry,1964,8:397-401.
[37]Gomez R M,Rojas A L,Rojas A N,et al.Colloidal chitin stained with Remazol Brilliant Blue R,a useful substrate to select chitinolytic microorganisms and to evaluate chitinases[J].Journal of Microbiological Methods,2004,56(2):213-219.
[38]Shirazi F,Kulkarni M,Deshpande M V.A rapid and sensitive method for screening of chitinase inhibitors using Ostazin Brilliant Red labelled chitin as a substrate for chitinase assay[J].Letters in Applied Microbiology,2007,44(6):660-665.
[39]Mamarabadi M,Jensen B,Lubeck M.Three endochitinaseencoding genes identified in the biocontrol fungus Clonostachys rosea are differentially expressed[J].Current Genetics,2008,54(2):57-70.
Review on purification,enzyme assay and application of plant chitinases
YANG Hai-xia1,DENG Jian-jun1,2,ZHANG Jian1,3,ZHAO Guang-hua1,*
(1.College of Food Science and Nutritional Engineering,China Agricultural University,Beijing 100083,China;2.Department of Food Science and Engineering,Northwest University,Xi’an 710069,China;3.Food College,Shihezi University,Shihezi 832003,China)
Plant chitinases which hydrolyze the chitin is one of pathogenesis-related proteins and can be induced in resistance of plants to fungal pathogens.Thus,plant chitinases has a wide range of application as a antisepticise material.Recently,chitinases from different plants have been purified and their enzymatic activities have been assayed with varied methods.The structures and classifications,different methods on enzymatic activity assay and purification were summarized.Moreover,applications of chitinases such as in food and medicine field were prospected.
plant;chitinases;classification;purification;enzymatic activity assay
TS201.2+5
A
1002-0306(2011)06-0431-05
2010-05-13 *通訊聯系人
楊海霞(1984-),女,博士研究生,研究方向:蛋白質化學與營養。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
