3種荒漠植物群落物種組成與豐富度的季節變化及地上生物量特征
2011-06-08 08:22:14陶冶張元明
草業學報 2011年6期
陶冶,張元明
(1.中國科學院新疆生態與地理研究所 干旱區生物地理與生物資源重點實驗室,新疆 烏魯木齊830011;2.中國科學院研究生院,北京100049)
群落結構與物種組成是群落演替的結果,它與生態系統的穩定性有密切關系[1,2]。研究發現,多數群落內的物種組成與多樣性均隨季節而變化,只是幅度不同而已[1,3-7]。荒漠是世界陸地生態系統的重要組成部分,約占陸地總面積的18.5%[8]。相比于森林生態系統,荒漠生態系統具有生物多樣性和生態系統穩定性較低的特點,因而極易受到外力干擾,而且擾動破壞后很難恢復[9]。因此,加強荒漠生態系統相關研究,對于人類深入認識和保護荒漠景觀具有重要而深遠的意義。
生物量是生態系統物質循環(碳循環)的重要載體,是植被生產力的體現,也是評價生態系統結構和功能的重要指標[10-12],越來越受到重視[13]。雖然全球荒漠地區的平均生物量較低(約7Mg/hm2),但其在世界碳循環中的重要地位逐漸被人們認識并引起重視[8]。以北美Sonoran沙漠為例,其面積約占世界荒漠面積的1%,而生物量儲量卻占4.4%,表現出了較強的、潛在的碳匯能力[13]。草地綜合順序分類法(comprehensive and sequential classification system of rangeland,CSCS)預測表明,到2050年,世界森林面積將有一定的減小,而草地和荒漠面積會明顯增加[16]。我國荒漠化土地約占國土面積的27.46%,其潛在面積更是達到47.1%[17],因而,荒漠地區生物量的研究對了解荒漠生態系統的生產力和承載力也十分重要。
準噶爾荒漠位于天山(南)和阿爾泰山(北)之間,東西分別以北塔山和塔爾巴哈臺山為界,屬于典型溫帶荒漠類型[18,19]。與天山南部的塔克拉瑪干沙漠及世界同緯度地區的其他荒漠相比,該荒漠植物物種最為豐富,因而構成了多樣的荒漠植物群落類型[20]。而且該荒漠還具有我國其他荒漠區沒有的植物生活型——短命和類短命植物(即短營養期植物),其在早春時期還能形成短命植物層片,表現出短暫的分布優勢。因而,它們的存在使準噶爾荒漠物種組成及其季節變化更具特色[21,22]。此外,準噶爾荒漠中心的古爾班通古特沙漠也是我國幾大沙源地之一,故人們對防沙治沙的重視也使得該地區成為研究的熱點區域[23]。近年來,關于荒漠地區植物群落結構與多樣性的研究較多[5,24-27],但對其季節動態的研究較少;對荒漠植被生物量分配與估算的研究雖有不少[12,28-30],但將生物量與物種豐富度相結合的研究極少[31]。為此,本研究選擇古爾班通古特沙漠3種典型植物群落,即蛇麻黃(Ephedradistachya)、白莖絹蒿(Seriphidiumterrae-albae)和準噶爾沙蒿(Artemisiasongarica)群落,通過研究其物種組成、豐富度的季節變化及不同生活型植物生物量特征,以探知不同季節群落結構變化特點,維持群落結構穩定的基礎及群落生物量分配格局,為深入了解該地區生態系統穩定機制奠定基礎,并為完善干旱荒漠區不同植被類型碳循環和碳儲量相關研究提供可靠的數據支持。
1 材料與方法
1.1 研究區概況
研究區位于古爾班通古特沙漠(44°11′~46°20′N,84°31′~90°00′E),處于準噶爾荒漠的中心。古爾班通古特沙漠面積約4.88×104km2,是我國第二大沙漠,也是我國最大的固定和半固定沙漠。年均蒸發量>2 000 mm,而年均降水量僅70~150mm,且主要集中于冬季(降雪)和春季(降雨);年均溫5~8℃,極端高溫為40℃以上,極端最低氣溫小于-40℃,≥10℃的年積溫可達3 000~3 500℃,空氣相對濕度平均50%~60%,5-8月通常<45%;該沙漠分布有各種形態的沙壟和一些新月型沙丘鏈、蜂窩狀沙丘,高度10~50m,沙丘主要為南北方向延伸;控制本區域的風系主要是西風氣流和蒙古高壓形成的西北和東北風系,起沙風(≥6m/s)集中于4-9月,其中以4-6月最為強盛[23]。
該沙漠共計208種植物,有18種植物群落類型和11種植物生活型,其中沙漠腹地主要由白梭梭(Haloxylonpersicum)、蛇麻黃、白莖絹蒿、準噶爾沙蒿和準噶爾無葉豆(Eremospartonsongoricum)等群落類型組成[22]。短命和類短命植物廣泛分布,成為早春荒漠生態系統的重要組成部分。此外,沙漠地表發育有良好的生物土壤結皮(包括藻結皮、地衣結皮、苔蘚結皮等),成為維持地表穩定的重要生物因子[18,32]。
1.2 研究方法
1.2.1 植被調查與草本植物生物量獲取 由于古爾班通古特沙漠(準噶爾荒漠)不同類型物種的花果期(生長期)具有明顯的不同時性[20-22],因此,本研究在時間尺度上分為初夏和秋初2個季節進行。初夏調查時(2010年5月底-6月初),在古爾班通古特沙漠中東部設置若干采樣點(圖1),位于昌吉回族自治州北部和阿勒泰地區(主要是福海縣)南部區域。每個采樣點附近調查1~3種群落。共調查蛇麻黃樣地36個、白莖絹蒿樣地28個、準噶爾沙蒿樣地13個,樣地大小均為20m×20m,以2條對角線上的8個5m×5m小樣方記錄樣地內的物種數、植株數,并在樣地內采集混合土樣(5次混合)1份。所有樣地四角均用木樁標記,然后用GPS定位。上述數據作為計算群落物種豐富度的原始數據。同時在對角線樣方內各設置1個1m×1m草本小樣方(共8個),用剪刀齊地面刈割獲取小樣方內所有短命和類短命植物地上部分,按不同物種分裝在布袋內,標記好后帶回實驗室沖洗干凈,于70℃烘48h,然后用精度0.000 1g天平稱重。小樣方內的草本數據用于估算樣地內初夏草本地上生物量。本研究中植物地上生物量均指現存量(standing biomass)。
秋季調查時(2010年8月),利用GPS定位信息,準確找到原始樣地,以5m×5m為小樣方詳細測量所有灌木的高度、冠幅(長軸長和短軸長),以估算灌木地上生物量。在每個樣地的8個對角線樣方內各設置1個1m×1m小樣方,收獲草本植物并分裝,帶回實驗室沖洗干凈后烘干稱重。小樣方內的草本數據用于估算秋季時樣地內的草本植物地上生物量。
1.2.2 灌木地上生物量估測方法 前期植被調查發現,3種群落共出現5種灌木,即蛇麻黃、白莖絹蒿、準噶爾沙蒿、準噶爾無葉豆和淡枝沙拐棗(Calligonumleucocladum)。除矮小灌叢蛇麻黃外,其他4種均可利用地上形態指標(株高、冠幅等)進行估算,這樣也減少了對植被的破壞。異速生長方程廣泛應用于樹木和灌木的生物量估算,其變型眾多[33]。本研究采用Y=aCVb模型估算灌木生物量[33,34],其中,Y為地上生物量,a為異速生長常數,b為異速生長指數,CV(cm3)為冠幅面積(cm2)×高(cm)。4種灌木地上生物量的擬合方程見表1。
因蛇麻黃為叢片狀分布,而且同一個樣地內的蛇麻黃高度分布較為均勻,因而本研究通過蓋度估算其地上生物量。首先測量得到樣地內蛇麻黃灌叢的總面積,隨后在樣地內隨機獲取3叢(0.5m×0.5m)蛇麻黃的蓋度和地上生物量,最后結合樣地內蛇麻黃灌叢總蓋度,進而估算出樣地內的蛇麻黃地上生物量。所有灌木樣品均置于70℃烘箱內烘72h后稱重(天平精度0.01g)。
1.2.3 物種生活型的確定及豐富度計算 物種生活型的判別有很多標準[35],本研究以張立運和陳昌篤[22]對古爾班通古特沙漠208種植物的劃分方法為準。為方便起見,本研究將灌木、半灌木和小半灌木統稱為灌木。物種豐富度以樣地內的物種數表示,分初夏和秋初2個季節計算和比較。
圖1 研究區及樣點分布Fig.1 Study area and the collection sites distribution in Gurbantunggut Desert

表1 古爾班通古特沙漠4種灌木地上生物量的擬合方程Table 1 Mathematical fitting equations of aboveground biomass for four shrubs in Gurbantunggut Desert
1.3 統計分析
對不同季節物種豐富度間進行Pearson相關分析,對物種豐富度與其地上生物量間進行二次曲線擬合;對不同群落類型物種豐富度、地上生物量間及初夏時3種群落中不同生活型植物物種數間進行One-way ANOVA分析,當方差為齊性時,采用LSD法進行多重比較,否則采用Tamhane’s T2方法。上述分析均采用SPSS 13.0統計軟件完成,常規數據處理和作圖在Excel 2003軟件上完成。
2 結果與分析
2.1 群落物種組成特征
3種灌木群落生境存在一定差異,蛇麻黃、白莖絹蒿群落主要存在于沙丘中下部和丘間平地,地表多存在以地衣和藻結皮為主的生物結皮,而準噶爾沙蒿群落主要存在于流動沙地、車轍和人工固沙區(沙障),其自然生境中的地表鮮見生物結皮。在夏季物種最豐富時期,3種群落分別記錄到63,61和40個物種,分屬于23,23和14科(表2),準噶爾沙蒿群落的物種數及科數明顯少于其他2種群落;秋季物種數及科數均迅速減少,主要存留一些菊科、藜科、禾本科和豆科植物,此時3種群落物種數(分別為15,14和16種)和科數(分別為9,8和7科)較為接近。夏季時,3種群落內菊科植物種數最多,所占比例均在21%以上;菊科、藜科、十字花科、禾本科、紫草科及豆科6個主要科的物種數分別占各群落總物種數的66.67%,63.93%和75%。總體來講,蛇麻黃和白莖絹蒿群落物種組成較為豐富,結構較為復雜,而準噶爾沙蒿群落物種組成和結構相對較簡單。
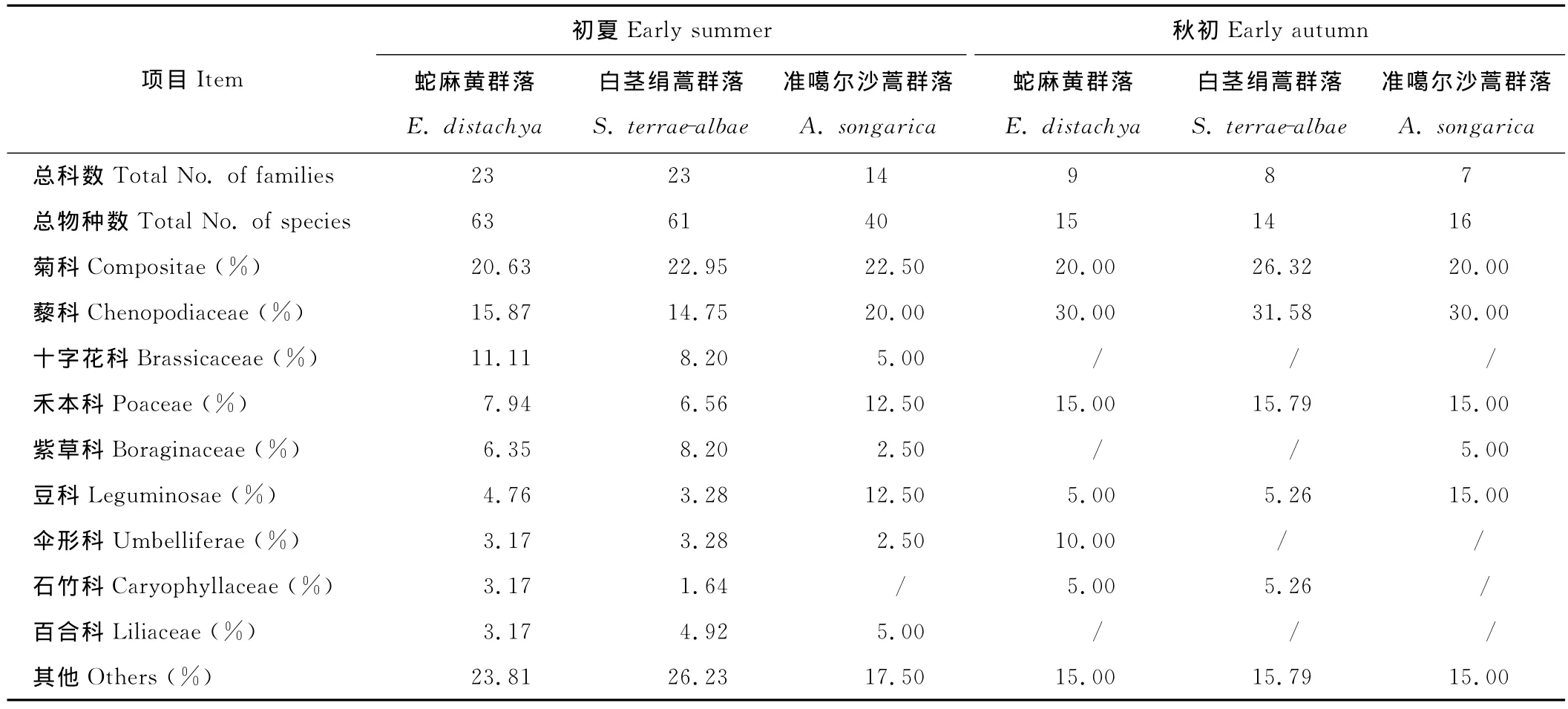
表2 初夏和秋初3種群落的科數和物種數及主要科內物種數的貢獻率Table 2 Total number of families and species and the relative contribution of species in main families of the three community types at early summer and early autumn
從植物生活型角度來看,各群落的物種均可劃分為多年生草本、一年生草本(二者屬于長營養期草本)、類短命植物、短命植物(二者屬于短營養期草本)和灌木共5種生活型。不同季節物種生活型組成不同(表3),但不同群落間變化趨勢一致。以蛇麻黃群落為例,夏季植物生活型組成最豐富(5種),以短命植物和一年生草本植物居多;而到了秋季,短命和類短命植物均已凋亡,僅剩3種生活型,此時一年生草本植物占據最大比例。3種群落中,不同生活型植物的數量比例也有所不同,其中蛇麻黃群落和白莖絹蒿群落較為相似,即灌木多草本少;而準噶爾沙蒿群落內的灌木和多年生草本的數量比例相對較高。

表3 初夏和秋初3種群落中植物生活型組成及其貢獻率Table 3 Life-form composition and relative contributions of the three community types at early summer and early autumn%
2.2 物種豐富度特征
各類群落的物種豐富度值均表現為夏季高于秋季,其中蛇麻黃和白莖絹蒿群落的物種豐富度在2個季節之間均差異顯著,而準噶爾沙蒿群落則無明顯差異(圖2),表明后者短命和類短命植物貢獻度較小(表3)。夏季時,3種群落間物種豐富度(物種數)總體上差異顯著(F=10.75,P<0.000 1),其中蛇麻黃和白莖絹蒿群落的物種豐富度均顯著高于準噶爾沙蒿群落;秋季時,3類群落間物種豐富度總體差異不顯著(F=1.12,P=0.332),此時,3類群落的物種豐富度均較低。同一季節,蛇麻黃和白莖絹蒿群落的物種豐富度均無顯著差異,表明二者物種組成和變化規律較為相似。
對初夏不同生活型植物物種數對比發現,在蛇麻黃和白莖絹蒿群落中,短命植物個數在各群落類型中均最大,其次是一年生草本,而多年生草本的物種數最少(圖3)。準噶爾沙蒿群落內的短命植物、一年生植物及灌木種類相當,而多年生草本和類短命植物種類最少。不同群落類型之間對比發現,蛇麻黃群落內的短命植物平均物種數最多,顯著高于白莖絹蒿群落,后者又顯著高于準噶爾沙蒿群落。
此外,各群落類型的物種豐富度在夏季和秋季之間均具有極顯著的正相關關系(表4)。這表明,群落內物種數的多少在季節上呈正相關,即如果一個群落內的夏季物種豐富度大,那么秋季也大,避免出現物種數在不同季節上的突然增多或減少的現象,因而體現了荒漠植物群落在物種組成和結構上也具有較強的穩定性。
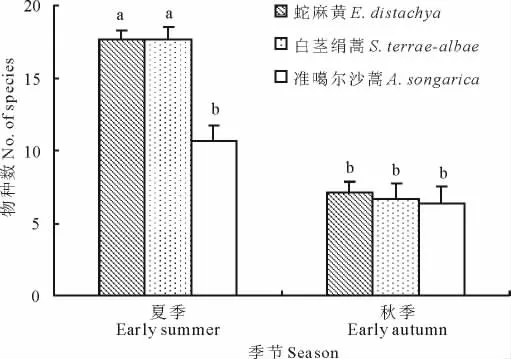
圖2 初夏和秋初3種群落不同季節的物種豐富度(平均值±SE)對比Fig.2 Richness(mean±SE)of the three community types at early summer and early autumn
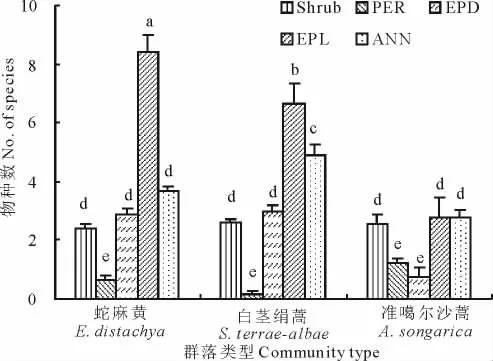
圖3 初夏3種群落不同生活型植物物種數(平均值±SE)比較Fig.3 Number of species(mean±SE)of different life-forms of the three community types at early summer

表4 3種群落物種豐富度在季節間的相關系數Table 4 Pearson correlation coefficients among species richness at early summer and early autumn
2.3 地上生物量特征
初夏時期,蛇麻黃群落中草本層植物(指短命和類短命植物)生物量最高(為6.99g/m2),白莖絹蒿群落次之(6.10g/m2),準噶爾沙蒿群落最低,僅為0.67g/m2(表5)。不同群落短命和類短命植物生物量所占比例不同,蛇麻黃和白莖絹蒿群落以短命植物為主(70%左右),而準噶爾沙蒿群落以類短命植物為主(占67.09%)。
秋季短命和類短命植物已凋亡,僅剩灌木、一年生和多年生草本。3類群落中,總地上生物量以白莖絹蒿群落最大(85.38g/m2),蛇麻黃群落次之(73.19g/m2),準噶爾沙蒿群落最低(54.21g/m2)(表6);3類群落的地上生物量均以灌木為主(均在95%以上),草本植物(指一年生和多年生草本)生物量比例很小(5%以下),其中以準噶爾沙蒿群落最小,其草本植物生物量僅為0.89g/m2,占1.65%。建群種生物量在各群落中均占絕對優勢,其分別占總地上生物量的86.03%,84.07%和89.65%。
與初夏相比,秋季蛇麻黃和白莖絹蒿群落內的草本層地上生物量均呈降低趨勢(分別降低57.65%和31.15%),而準噶爾沙蒿群落則有所增加(32.84%),但由于該群落夏季和秋季草本植物生物量均低于1g/m2,因而這種增長意義不大。調查顯示,3種群落在該沙漠的分布范圍為蛇麻黃>白莖絹蒿>準噶爾沙蒿,故總體來講,3種群落夏季草本層地上生物量大于秋季。

表5 初夏時期3種群落內短命和類短命植物的地上生物量及其比例 (平均值±標準誤)Table 5 Aboveground biomass of ephemeral plant and ephemeroid plant and their percentages of the three community types at early summer(mean±SE)
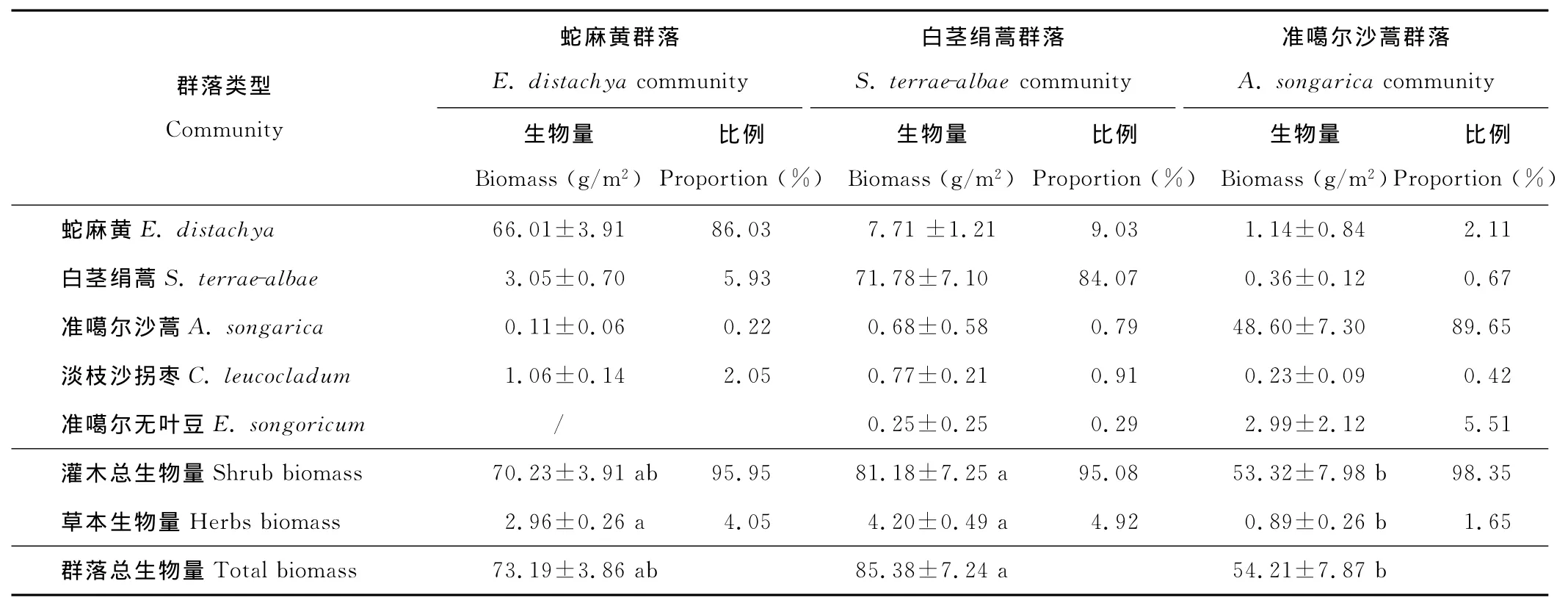
表6 秋初3種群落的總地上生物量、灌木和草本植物地上生物量及其貢獻率 (平均值±標準誤)Table 6 Aboveground biomass of total species,shrubs and herbs and their relative contributions of the three community types at early autumn(mean±SE)
2.4 地上生物量與物種豐富度的關系
無論夏季還是秋季,77個研究群落內的草本植物物種豐富度與其地上生物量之間均呈顯著的“單峰型關系”(圖4),說明起初草本植物豐富度隨著草本植物生物量的增加而增加,但到一定程度時反而呈降低趨勢,也說明中等生物量時草本植物豐富度最高。該結論與陳生云等[36]、Chalcraft等[37]及Oba等[38]的研究結果一致。與秋季相比(P=0.0121),這種規律在初夏時最為顯著(P<0.000 1),主要歸因于該時期較豐富的草本植物種類(主要是短命植物)。但是,若把灌木生物量考慮進來,那么無論夏季還是秋季,物種豐富度與生物量間均無規律可循,這可能是灌木生物量較大但種類稀少的緣故。
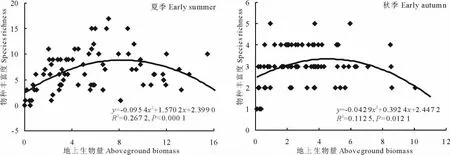
圖4 初夏和秋初草本植物物種豐富度與其地上生物量的關系Fig.4 Relationships between herbaceous richness and their aboveground biomass at early summer and early autumn
3 討論
3.1 植物生活型與豐富度的季節變化及其生態學意義
本研究中,初夏植物生活型有5種,秋季僅3種;同時,物種豐富度在3類群落內也表現出夏季大、秋季小的規律,這與草本植物,尤其是短命和類短命植物的出現與消失密切相關。可見,草本植物對荒漠植物群落物種生活型和豐富度的變化起到至關重要的作用,而且這種作用與群落類型無關。早春短命和類短命植物是第三紀末第四紀初,古地中海退卻以后,由熱植物區系衍生出來的年輕植物類群。其分布中心為中亞,地中海沿岸、西亞、北非等地也有。我國僅新疆北部有分布(約205種),為其分布區的最東端[21]。在古爾班通古特沙漠,由于冬季有一定的降雪和早春一定的降雨,使早春時期土壤較為濕潤[22]。此時,雖然溫度較低,但短命和類短命植物已開始萌發(3月底或4月初),一般在5-6月開花結果,隨即完成生活史[39]。王雪芹等[23]報道,該沙漠起沙風(≥6 m/s)集中于4-9月,其中以4-6月最為強盛,該時期正是短命植物大量生長的時期。因而,無論沙塵暴發生的頻率還是強度,該沙漠都要比我國其他沙塵暴源區小得多。那么,短命和類短命植物的存在無疑成為沙塵暴危害降低的一個主要原因。本區短命和類短命植物還能形成層片或小群落,以致枯死后其生存的一些地帶性群落才逐漸顯露出本來面目[21,22]。這表明短命和類短命植物不僅能有效維持地表穩定,還能參與和改變荒漠植物群落的物種構成,對維持生物多樣性和荒漠生態系統穩定性均具有重要意義。
此外,當夏末短命植物消亡后,一年生草本便迅速生長,并逐漸占據群落下層的優勢地位,填補了短命和類短命植物死亡后留下的資源位空缺。可見,短營養期草本植物和長營養期草本植物存在的這種時間上的交錯,不僅使它們在資源利用上盡可能減少競爭,也促使它們形成了不同時間段上的“相似功能群”。因而,一年生草本植物同樣對維持荒漠生態系統穩定起重要作用。
3.2 幾種荒漠類型生物量對比及其影響因素
荒漠是世界生態系統的重要組成部分。在世界眾多生態系統類型中,荒漠生態系統平均生物量(地上+地下)為7Mg/hm2[8],其中以北美典型熱荒漠——Sonoran沙漠最高,達到32Mg/hm2,該地區主要建群種為仙人掌(Opuntia)類植物[13]。相比之下,北美的2個溫帶荒漠——Mojave和Chihuahuan沙漠生物量較低,僅為1.8~2.47Mg/hm2[40]和1.6~3.2Mg/hm2[41]。對比我國幾種溫帶荒漠發現,烏蘭布和東南緣沙漠梭梭(Haloxylon)群落(6.42~16.41Mg/hm2)[28,29]和準噶爾荒漠東南緣沙漠的梭梭群落(6.69~19.39Mg/hm2)[30]的生物量均接近或高于世界平均水平,鄂爾多斯荒漠的油蒿(Artemisiaordosica)群落也較高(4.7~10.9Mg/hm2)[42],但毛烏素沙地的油蒿群落(0.12~2.35Mg/hm2)[34]卻遠低于世界平均值。由于本研究未測定地下生物量,因此,準確估計這3類群落生物量還比較困難。但是,即使按照根冠比1∶1(實際值可能小于1∶1)計算地下生物量[13],3類群落秋季總生物量(地上+地下)也僅接近于毛烏素沙地的油蒿群落,但仍低于已知其他荒漠類型和世界荒漠生物量的平均值。
影響生物量的可能因素主要為:1)在全球尺度上,由氣候條件所決定的生態系統(群落)類型。生物量表現為熱帶森林>溫帶森林>地中海灌叢>北方森林>薩王納群落>溫帶草地≥荒漠=北極苔原>農田的規律[8]。可見,荒漠地區的生物量遠遠小于其他絕大部分生態系統類型。2)中小尺度上,除氣候條件外,小地形、微氣候等環境因子可能直接影響小尺度上的植被結構與物種組成,進而影響植被生物量。降水、溫度、地下水位、土壤類型等因素在小尺度上對植物多樣性及生物量均有不同影響,這在各類控制或野外試驗中均得到證實[43-47]。對于短命植物類群來講,冬季降雪、早春降水、氣溫、土壤含水量等可能是最主要影響因素[39]。3)人為干擾,如放牧、刈割、工程活動等。眾多研究認為,中度刈割和放牧有利于提高草地地上生物量產量及物種多樣性,而高強度干擾則顯著降低生物量和多樣性[38,48]。就本研究而言,3類群落處在相同的氣候和土壤類型條件下,那么影響群落生物量差異的主要原因可能是坡位、地表穩定性、土壤水分等因素引起的群落結構的差異。
3.3 荒漠植物群落中草本植物生物量貢獻度
本研究中,秋季蛇麻黃群落、白莖絹蒿群落和準噶爾沙蒿群落中草本植物生物量分別為2.96,4.20和0.89 g/m2,即使夏季時,其生物量也僅為6.99,6.10和0.67g/m2。從比例上看,3類群落中草本植物生物量在群落總生物量中的比例均<5%。王春玲等[30]、宋朝樞和賈昆峰[29]對吉木薩爾地區及烏蘭布和地區的梭梭群落生物量研究發現,其林下層植物(包含林下小灌木、多年生草本、一年生草本)生物量比例僅占1.13%~2.39%和2.01%~4.21%;那么,僅就林下草本層而言,其生物量比例在梭梭群落中更小。因而認為,在大尺度生物量估算時,草本植物生物量可以忽略不計。但即便如此,草本植物尤其是短命和類短命植物的生態作用是不可忽視的。
致謝:中國科學院新疆生態與地理研究所周曉兵博士、吳林碩士及新疆農業大學趙偉、丁陽、劉星等同學在野外數據采集方面給予大力幫助,新疆農業大學邱東女士在數據錄入與處理上給予無私幫助,新疆額爾齊斯河流域開發工程建設管理局沙漠一站也提供了諸多便利,在此一并感謝!
[1]婁彥景,趙魁義,馬克平.洪河自然保護區典型濕地植物群落組成及物種多樣性梯度變化[J].生態學報,2007,27(9):3883-3891.
[2]祝燕,趙谷風,張儷文,等.古田山中亞熱帶常綠闊葉林動態監測樣地——群落組成與結構[J].植物生態學報,2008,32(2):262-273.
[3]陳文河,劉學東,盧伙勝.南沙群島海域魚類群落結構的季節性變化研究[J].熱帶海洋學報,2011,29(4):118-124.
[4]陳瑩,馬鳴,李維東,等.新疆阜康荒漠區不同生境鳥類群落的季節變化[J].生態學雜志,2011,30(2):273-280.
[5]杜茜,馬琨.寧夏荒漠草原恢復演替過程中物種多樣性與生產力的變化[J].生態環境,2007,16(4):1225-1228.
[6]費永俊,魯文俊,韓烈保.荊江大堤護坡草本植物物種多樣性及其季節動態[J].草業科學,2007,24(10):1-5.
[7]李步杭,張健,姚曉琳,等.長白山闊葉紅松林草本植物多樣性季節動態及空間分布格局[J].應用生態學報,2008,19(3):467-473.
[8]Houghton R A,Hall F,Goetz S J.Importance of biomass in the global carbon cycle[J].Journal of Geophysical Research,2009,114:G00E03,doi:10.1029/2009JG000935.
[9]李博.生態學[M].北京:高等教育出版,2000.
[10]韓龍,郭彥軍,韓建國,等.不同刈割強度下羊草草甸草原生物量與植物群落多樣性研究[J].草業學報,2010,19(3):70-75.
[11]李士美,謝高地,張彩霞.典型草地地上現存生物量資產動態[J].草業學報,2009,18(4):1-8.
[12]再努拉·熱和木都拉,陳亞寧,艾里西爾·庫爾班,等.準噶爾盆地梭梭群落地上生物量及季節變化研究——以克拉瑪依農業綜合開發區為例[J].干旱區地理,2006,29(3):375-380.
[13]Búrquez A,Martínez-Yrízar A,Núez S,etal.Aboveground biomass in three Sonoran Desert communities:Variability within and among sites using replicated plot harvesting[J].Journal of Arid Environments,2010,74:1240-1247.
[14]Fang J Y,Chen A P,Peng C H,etal.Changes in forest biomass carbon storage in China between 1949and 1998[J].Science,2001,292:2320-2322.
[15]方精云,唐艷鴻,Son Y.碳循環研究:東亞生態系統為什么重要[J].中國科學:生命科學,2010,40(7):561-565.
[16]任繼周,梁天剛,林慧龍,等.草地對全球氣候變化的響應及其碳匯潛勢研究[J].草業學報,2011,20(2):1-22.
[17]吳波,蘇志珠,陳仲新.中國荒漠化潛在發生范圍的修訂[J].中國沙漠,2007,27(6):911-917.
[18]張元明,王雪芹.準噶爾荒漠生物結皮研究[M].北京:科學出版社,2008.
[19]趙白.準噶爾盆地的形成與演化[J].新疆石油地質,1992,13(3):192-196.
[20]張新時.天山北部山地-綠洲-過渡帶-荒漠系統的生態建設與可持續農業范式[J].植物學報,2001,43(12):1294-1299.
[21]毛祖美,張佃民.新疆北部早春短命植物區系綱要[J].干旱區研究,1994,11(3):1-26.
[22]張立運,陳昌篤.論古爾班通古特沙漠植物多樣性的一般特點[J].生態學報,2002,22(11):1923-1932.
[23]王雪芹,蔣進,雷加強,等.古爾班通古特沙漠短命植物分布及其沙面穩定意義[J].地理學報,2003,58(4):598-605.
[24]柴永青,曹致中,蔡卓山.肅北地區裸果木荒漠群落構成特征及物種多樣性研究[J].草業學報,2010,19(1):21-27.
[25]Zhu W B,Lv A F,Jia S F.Spatial distribution of vegetation and the influencing factors in Qaidam Basin based on NDVI[J].Journal of Arid Land,2011,3(2):85-93.
[26]Qian Y B,Zhang H Y,Wu Z N,etal.Vegetation composition and distribution on the northern slope of Karlik Mountain to Naomaohu Basin,East Tianshan Mountains[J].Journal of Arid Land,2011,3(1):15-24.
[27]何明珠.阿拉善高原荒漠植被組成分布特征及其環境解釋V.一年生植物層片物種多樣性及其分布特征[J].中國沙漠,2010,30(3):528-533.
[28]李鋼鐵,張密柱.梭梭林生物量研究[J].內蒙古林學院學報,1995,17(2):35-43.
[29]宋朝樞,賈昆峰.烏拉特梭梭林自然保護區科學考察集[M].北京:中國林業出版社,2000.
[30]王春玲,郭泉水,譚德遠,等.準噶爾盆地東南緣不同生境條件下梭梭群落結構特征研究[J].應用生態學報,2005,16(7):1224-1229.
[31]鄭曉翾,靳甜甜,木麗芬,等.呼倫貝爾草原物種多樣性與生物量、環境因子的關系[J].中國草地學報,2008,30(6):74-81.
[32]Zhang Y M,Wu N,Zhang B C,etal.Species composition,distribution patterns and ecological functions of biological soil crusts in the Gurbantunggut Desert[J].Journal of Arid Land,2010,2(3):180-189.
[33]Djomo A N,Ibrahima A,Scaborowski J,etal.Allometric equations for biomass estimation in Cameroon and pan moist tropical equations including biomass data from Africa[J].Forest Ecology and Management,2010,260:1873-1885.
[34]Li C P,Xiao C W.Above-and belowground biomass ofArtemisiaordosicacommunities in three contrasting habitats of the Mu Us desert,northern China[J].Journal of Arid Environments,2007,70:195-207.
[35]Cain S A.Life-forms and phytoclimate[J].Botanical Review,1950,16(1):1-32.
[36]陳生云,劉文杰,葉柏生,等.疏勒河上游地區植被物種多樣性和生物量及其與環境因子的關系[J].草業學報,2011,20(3):70-83.
[37]Chalcraft D R,Wilsey B J,Bowles C,etal.The relationship between productivity and multiple aspects of biodiversity in six grassland communities[J].Biodiversity and Conservation,2009,18:91-104.
[38]Oba G,Vetaas O,Stenseth N S.Relationships between biomass and plant species richness in arid-zone grazing lands[J].Journal of Applied Ecology,2001,38:836-845.
[39]蘭海燕,張富春.新疆早春短命植物適應荒漠環境的機理研究進展[J].西北植物學報,2008,28(7):1478-1485.
[40]Rundel P W,Gibson A C.Ecological Communities and Processes in a Mojave Desert Ecosystem,Rock Valley Nevada[M].Cambridge:Cambridge University Press,1996.
[41]Huenneke L F,Schlesinger W H.Patterns of net primary production in Chihuahuan Desert ecosystems[A].In:Havstad K M,Huenneke L F,Schlesinger W H.Structure and Function of a Chihuahuan Desert Ecosystem[M].New York:Oxford University Press,2006.
[42]Jin Z,Qi Y,Dong Y.Storage of biomass and net primary productivity in desert shrubland ofArtemisiaordosicaon Ordos Plateau of Inner Mongolia,China[J].Journal of Forestry Research,2007,18:298-300.
[43]何丹,李向林,萬里強,等.施用尿素當年對退化天然草地物種地上生物量和重要值的影響[J].草業學報,2009,18(3):154-158.
[44]馮燕,王彥榮,胡小文.水分脅迫對兩種荒漠灌木幼苗生長與水分利用效率的影響[J].草業學報,2011,20(4):293-298.
[45]蘇丹,孫國峰,張金政,等.水分脅迫對費菜和長藥八寶生長及生物量分配的影響[J].園藝學報,2007,34(5):1317-1320.
[46]王麗,胡金明,宋長春,等.水分梯度對三江平原典型濕地植物小葉章地上生物量的影響[J].草業學報,2008,17(4):19-25.
[47]蔡仕珍,李璟,潘遠智,等.不同種植密度對烏頭生長發育的影響[J].草業學報,2011,20(2):278-282.
[48]呼格吉勒圖,楊劼,寶音陶格濤,等.不同干擾對典型草原群落物種多樣性和生物量的影響[J].草業學報,2009,18(3):6-11.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
成才之路(2017年21期)2017-07-27 20:52:40
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
安徽農學通報(2017年9期)2017-05-19 23:03:05
現代農業科技(2016年22期)2017-03-24 12:21:57
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
學周刊·下旬刊(2015年10期)2015-07-22 12:36:17
