沙生植物差巴嘎蒿光合特性及其模擬研究
2011-08-20 04:06:08李熙萌盧之遙馬帥馮金朝
草業學報 2011年6期
李熙萌,盧之遙,馬帥,馮金朝
(中央民族大學生命與環境科學學院,北京100081)
差巴嘎蒿(Artemisia halodendron)為高蒸騰、高耗水型沙地旱生植物,主要分布于科爾沁沙地和呼倫貝爾沙地,為流動、半固定沙地植被建群種,在生物固沙中起著重要作用[1]。目前,國內外對差巴嘎蒿的研究主要集中于根系動態[2]、種群生態特征及生態位[3]和種子散布格局等方面[4],對自然狀態下差巴嘎蒿光合特性的深入研究報道較少。光合作用是植物重要的生理過程,研究植物的光合特性,有利于了解植物對光能的利用效率,闡明植物光合的生態學特征[5]。不同物種以及同一物種的不同品種、甚至同一品種在不同地區的光合特性各有差異,因此,有必要在大范圍對物種展開多層次、多角度的科學研究[6]。本試驗對科爾沁沙地自然條件下差巴嘎蒿葉片光合作用進行了觀測和模擬研究,旨在揭示自然條件下差巴嘎蒿葉片光合作用特征,闡明其光合作用變化規律及其與環境之間的相互關系,為防治沙漠化、維持沙地生態平衡提供科學依據。
1 材料與方法
1.1 樣地與材料
試驗觀測在中國科學院奈曼沙漠化研究站進行。試驗地點位于科爾沁沙地中南部奈曼旗境內,地理位置為42°58′N,120°44′E,海拔約為360m。觀測地點的沙地類型為半固定沙地,植被蓋度為20%~40%。該地區氣候特征屬溫帶大陸性半干旱氣候,輻射較強烈,水熱同期;年均氣溫6.51℃,最熱月(7月)平均氣溫23.5℃,最冷月(1月)平均氣溫-13.2℃;多年平均降水量為360mm左右,主要集中在6-8月,年蒸發量1 935mm;年太陽輻射總量為5 200~5 400MJ/m2,全年≥10℃的年積溫為3 200~3 400℃,無霜期151d[7]。
供試材料為半固定沙地自然生長的差巴嘎蒿。選用當年生枝條上部成熟葉片為試驗材料。
1.2 試驗方法
1.2.1 測定方法 試驗觀測于2009年7月下旬進行。差巴嘎蒿葉片光合速率及光響應曲線的測定采用美國LICOR-6400型便攜式氣體分析系統(LICOR,美國)。
光合作用日變化的測量選擇晴朗無云的天氣進行。從早晨6:00開始,每隔2h測定1次,全天觀測共7次;每次觀測有3~5個重復。測定因子包括:凈光合速率(net photosynthetic rate,Pn,μmol/m2·s)、氣孔導度(stomatal conductance,Gs,mmol/m2·s)、胞間 CO2濃度(intercellular CO2concentration,Ci,μmol/mol)、蒸騰速率(evaporation rate,E,μmol/m2·s)、光合有效輻射(photosynthetic active radiation,Par,μmol/m2·s)、大氣CO2濃度(ambient CO2concentration,Ca,μmol/mol)、環境溫度(ambient temperature,Ta,℃)、空氣相對濕度(relative humidity,RH,%)等。
光響應曲線的測量選擇晴朗天氣,在上午10:00-11:00進行,每次觀測3個重復。光強由LICOR-6400紅藍光源進行控制,在不同光合有效輻射通量密度下(photosynthetic flux density,PFD:2 000,1 500,1 200,1 000,800,600,400,200,100,50,0μmol/m2·s)測定其凈光合速率。測量過程中葉室溫度控制在25℃,CO2濃度控制在380μmol/mol,相對濕度控制在50%。
氣孔限制值(stomatal limition,Ls)的計算參照Berry和Downton[8]的方法:Ls=1-Ci/Ca。
植物水分利用效率(water use efficiency,WUE)采用下式計算:WUE=Pn/E[9]。
1.2.2 模型選擇 有很多模型能夠描述植物光合作用與光強之間的關系,包括二項式回歸法及各種光響應模型,其中光合模型又分為雙曲線模型和指數模型2類。本研究采用二項式回歸法和常用的5種光響應模型對差巴嘎蒿光響應曲線進行擬合[10-14]。氣孔導度模型也有多種不同的表達形式。本研究中選用Ball等[15]提出的半經驗模型及葉子飄和于強[16]提出的氣孔導度修正模型對數據進行擬合。
2 結果與分析
2.1 Pn、E及 WUE的日變化
從6:00開始,差巴嘎蒿凈光合速率Pn隨Par升高而增大,8:00時達到峰值,為10.37μmol/(m2·s)。此后,Par繼續升高,Pn呈下降趨勢,14:00最低;此后,Pn逐漸上升,16:00時出現第2峰值,為4.85μmol/(m2·s)。16:00后,Pn隨Par的降低而降至最低。
差巴嘎蒿葉片E的日變化曲線為單峰型,其峰值出現在10:00,較Pn峰值出現時間晚(圖1);差巴嘎蒿葉片WUE日變化曲線為雙峰型,變化規律與Pn相似,在8:00和16:00達到峰值,14:00WUE為最小值(圖1)。
2.2 光合作用的氣孔調節
差巴嘎蒿Gs的日變化曲線呈雙峰型(圖2),一天中的高峰出現在8:00,為0.16mmol/(m2·s),午后峰值出現在14:00,但此峰并不明顯。Pn與Gs的變化并不同步,但第1峰值出現時間相同。
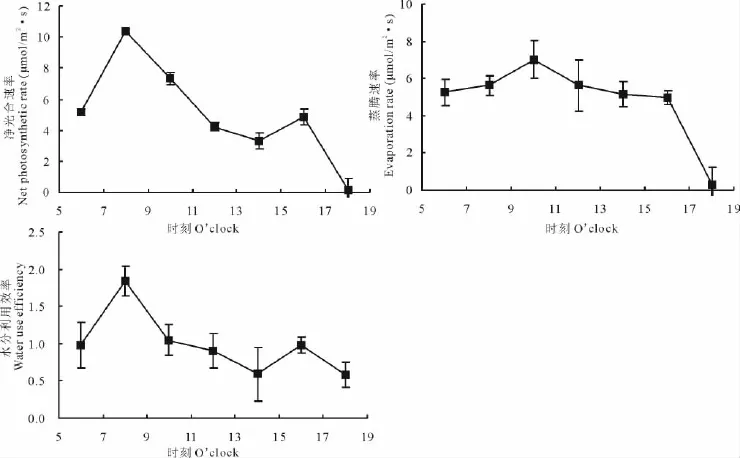
圖1 差巴嘎蒿葉片Pn、E及WUE的日變化Fig.1 The diurnal courses of Pn、E and WUE of A.halodendron
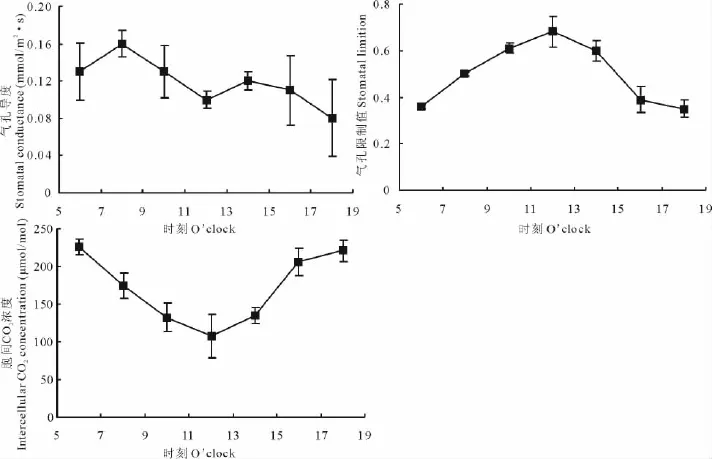
圖2 差巴嘎蒿Gs、Ci和Ls的日變化Fig.2 The diurnal courses of Gs、Ci and Ls in leaves of A.halodendron
Ls的日變化規律呈單峰曲線,從6:00開始逐漸增大,12:00達到峰值,此后逐步下降。Ci日變化同Ls相反,其最大值出現在6:00,12:00最低,之后開始回升,到18:00時基本恢復到清晨水平。
2.3 光響應曲線的模擬
決定系數和殘差標準差均可反映模型擬合的精度,前者隨精度升高而增大,后者則相反。本研究使用決定系數和殘差標準差共同判斷擬合結果的優劣。光響應曲線的直角雙曲線修正模型和氣孔導度修正模型決定系數最大,同時殘差標準差最小,具有最高的擬合精度(表1,2,圖3)。

表1 光響應模擬及擬合結果Table 1 Light-response curve model and fitting results

表2 氣孔導度模型及擬合結果Table 2 Stomatal conductance model and fitting results
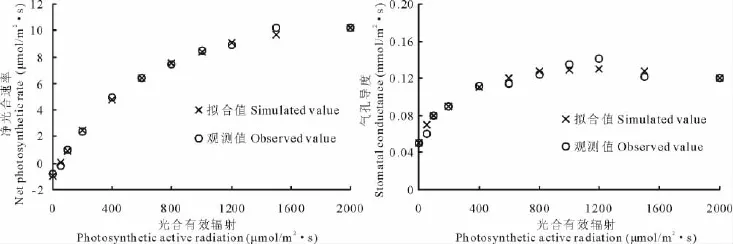
圖3 光響應修正模型及氣孔導度修正模型擬合結果Fig.3 The fitting effect of modified light response model and stomatal conductance model
3 討論
植物的光合日動態變化反映了植物一天中光合作用隨著環境變化而發生的變化,是植物對特定環境條件的適應結果[17]。觀測結果表明,自然條件下差巴嘎蒿光合速率日變化曲線為雙峰型,具有明顯的“午休”現象。光合“午休”是植物在長期進化過程中形成的一種適應干旱環境的方法[18]。一般認為,導致光合“午休”的原因大致可以歸結于氣孔因素和非氣孔因素2個方面。氣孔因素主要指光合作用的氣孔調節,具體表現在Gs和Ls的大小,前者反映氣孔的開放程度,后者反映氣孔對CO2的阻力,而非氣孔因素則指葉肉細胞的羧化能力。為了區分光合作用的氣孔限制與非氣孔限制,Farquhar和Sharkey[19]提出了一個判別方法,即當Ci降低,同時Ls增大時,才可以判定光合速率降低是由Gs降低引起的,即Pn的降低是由氣孔因素造成的。相反,如果葉片Pn的降低伴隨著Ci的升高,那么Pn的下降要歸因于非氣孔因素,即葉肉細胞羧化能力的下降。由此可知,從6:00開始,隨著光照強度及溫度的升高,差巴嘎蒿葉片氣孔逐步打開,凈光合速率和蒸騰速率也隨之升高,到8:00時氣孔導度最大,葉片氣體交換能力最強,光合速率達峰值,此后,光強和溫度進一步升高,為了避免過度失水,氣孔逐漸關閉,導致氣體交換能力降低,CO2供應不足,凈光合速率開始下降,也即凈光合速率在8:00-12:00期間下降是由氣孔因素造成的[20,21]。14:00后,光強與溫度降低,差巴嘎蒿凈光合速率有所回升,到16:00時到達第2峰值。此峰值低于上午,注意到此時溫度較第1峰值出現時高,故推測第2峰值的降低可能是高溫導致的葉片光、暗呼吸加強的結果。此后,Ls降低,Ci升高,凈光合速率的降低則是由于非氣孔因素造成的。
水分利用效率(WUE)是由植物的Pn和E兩方面決定的,即消耗單位重量的水,植物所固定的CO2量。植物為維持水分平衡,在選擇優先光合和優先蒸騰之間達到適度的調節[22]。本研究中,差巴嘎蒿的 WUE上午和下午各有1個高峰,在午間強光和高溫的環境條件下氣孔關閉,最大程度減少蒸騰失水,顯示出較強的抗旱能力。WUE峰值與Pn峰值同時出現,暗示WUE與Pn之間的關系更為密切,即WUE的增大在更大程度上依賴于Pn的上升,同時也說明光合作用對其適應干旱環境的重要性。
光響應曲線反映了在一定范圍光照強度下植物潛在的光合能力以及植物對生長環境的適應性[6],對植物葉片光響應曲線的測量可以確定光飽和點、光補償點、飽和光強和表觀量子效率等指標,對了解光反應過程的效率非常重要。迄今為止,國內外學者已經建立了許多光響應模型以描述光強同凈光合速率及氣孔導度之間的關系。由于不同模型的意義不完全相同,而不同植物種及同一植物不同部位葉片的生理特性也有差別,因而應用同一模型進行不同植物種的光響應曲線擬合必將產生一定的誤差[23]。基于此,采用不同的光合-光響應曲線和氣孔導度-光響應曲線進行了擬合。結果顯示,使用直角雙曲線修正模型和氣孔導度修正模型擬合,決定系數大于其他模型,而殘差標準差也最小,表明對于差巴嘎蒿,這2種模型具有較高的擬合精度,對于其光合特性有較強的預測性。
[1] 周海燕,趙愛芬.科爾沁草原主要牧草冷蒿和差不嘎蒿的生理生態學特性與競爭機制[J].生態學報,2002,22(6):894-900.
[2] 趙瑋,張銅會,劉新平,等.差巴嘎蒿灌叢土壤和根系含水量對降雨的響應[J].生態學雜志,2008,27(2):151-156.
[3] 樸順姬,王振杰,顏秀靈,等.科爾沁沙地差巴嘎蒿種群生態位適宜度分析[J].植物生態學報,2006,30(4):593-600.
[4] 趙麗婭,李兆華,趙錦慧,等.流動沙丘先鋒灌木差巴嘎蒿的種子散布格局[J].西北植物學報,2005,25(9):1821-1826.
[5] 楊秀芳,玉柱,徐妙云,等.2種不同類型的尖葉胡枝子光合-光響應特性研究[J].草業科學,2009,26(7):61-65.
[6] 韋蘭英,曾丹娟,張建亮,等.巖溶石漠化區四種牧草植物光合生理適應性特征[J].草業學報,2010,19(3):212-219.
[7] 李玉霖,張銅會,崔建垣.科爾沁沙地農田玉米耗水規律研究[J].中國沙漠,2002,22(4):354-358.
[8] Berry J A,Downton J S.Environmental regulation of photosynthesis[A].Photosynthesis:Development,Carbon Metabolism and Plant Productivity[M].New York:Academic Press,1982:263-343.
[9] Fischer R A,Turner N C.Plant productivity in the arid and semiarid zones[J].Annual Reivew of Plant Physiology,1978,29:227-317.
[10] Baly E C.The kinetics of photosynthesis[J].Proceedings of the Royal Society Ser B(Biological Sciences),1935,117(104):218-239.
[11] Ye Z P.A new model for relationship between light intensity and the rate of photosynthesis in Oryza sativa[J].Photosynthetica,2007,45:637-640.
[12] Thornley J H M.Mathematical Models in Plant Physiology[M].London:Academic Press,1976:86-110.
[13] Prado C H B A,Moraes J A P V.Photosynthetic capacity and specific leaf mass in twenty woody species of Cerrado vegetation under field condition[J].Photosynthetica,1997,33(1):103-112.
[14] Bassman J,Zwier J C.Gas exchange characteristics of Populus torichocarpa,Populus deltoids and Populus torichocarpa×P.deltoids clones[J].Tree Physiology,1991,8(2):145-159.
[15] Ball J T,Woodrow I E,Berry J A.A model predicting stomatal conductance and its contribution to the control of photosynthesis under different environmental conditions[A].In:Biggens J.Progress in Photosynthesis Research[M].Dordrecht:Martinus Nijhoff Publishers,1987:221-224.
[16] 葉子飄,于強.植物氣孔導度的機理模型[J].植物生態學報,2009,33(4):772-782.
[17] 高鶴,宗俊勤,陳靜波.7種優良觀賞草光合生理日變化及光響應特征研究[J].草業學報,2010,19(4):87-93.
[18] 張利平,王新平,黃子琛,等.沙漠地區沙冬青的氣體交換特征研究[J].西北植物學報,1998,18(2):207-213.
[19] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[20] 黃立華,梁正偉,馬紅媛.蘇打鹽堿脅迫對羊草光合、蒸騰速率及水分利用效率的影響[J].草業學報,2009,18(5):25-30.
[21] 賈洪濤,趙成義,盛鈺,等.干旱地區紫花苜蓿光合日變化規律研究[J].草業科學,2009,26(7):56-60.
[22] Robert E S,Mark A,John S B.Kok effect and the quantum yield of photosynthesis-light partially inhibits dark respiration[J].Plant Physiology,1984,75:95-101.
[23] 王秀偉,毛子軍.7個光響應曲線模型對不同植物種的實用性[J].植物研究,2009,29(1):44-48.
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
光學精密工程(2016年6期)2016-11-07 09:07:19
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
核科學與工程(2015年4期)2015-09-26 11:59:03
