NF-κB抑制劑PDTC對小鼠肝癌細胞生長的影響
2011-06-01 01:03:44何順亮周海華余文娟宋瑞祥施廣霞郭連英
大連醫科大學學報 2011年4期
何順亮,司 敏,周海華,余文娟,宋瑞祥,施廣霞,郭連英,郝 冰
(大連醫科大學 1.臨床醫學七年制2006級,2.臨床醫學2007級,3.病理生理教研室,遼寧 大連 116044)
核轉錄因子NF-κB是首先由Sen、Baltimore、Mercurio等,在1986年從成熟B 淋巴細胞和漿細胞核的抽提物中發現的核蛋白因子,因能與免疫球蛋白的κ輕鏈基因內含增強子的特異性序列(即κB序列,10bP : 5’-GGGACTTTCC-3’)特異結合并促進κ鏈基因表達而被稱為核因子κB ( nuclear factor - κB,NF -κB)[1,2],該因子是一個由復雜的多肽亞單位組成的蛋白家族,包括Rel ( c - Rel) ,RelA(p65) ,RelB,NF -κB1 (p50)和NF -κB2 (p52、p49、p50B)。NF -κB是真核生物中廣泛存在的一種多向性、多功能的核轉錄因子,可調控細胞凋亡、增殖的相關基因,在細胞癌變過程中發揮重要作用。抗氧化劑吡咯烷二硫代氨基甲酸鹽 (Pyrro1idine dithiocarbamate ,PDTC) 是NF-κB的特異性抑制劑,PDTC可抑制淋巴瘤、乳腺癌和卵巢癌等腫瘤細胞的增殖,促進細胞凋亡,抑制促炎性因子的分泌[3-5]。本實驗用體外細胞培養和體內抑瘤實驗研究PDTC 對小鼠腹水型肝癌Hca-F的抗腫瘤作用。
1 材料和方法
1.1 動 物
Balb/c小鼠,無特定病原體, 雌雄各半,共20只,體重雌鼠18~32 g,雄鼠27~39 g。
1.2 藥品、試劑與器材
PDTC購于sigma公司; MTT購于Amresco公司,生產批號:0793;DMSO購于天津市光復精細化工研究所,生產批號:060831,丙二醛(Maleic Dialdehyde MDA)試劑盒購于南京建成生物工程研究所。低溫高速離心機(EVOLUTION TMKC,美國),酶標儀(BIO-RADmodel550,日本)。
1.3 腫瘤細胞株
小鼠Hca-F細胞株為源自小鼠肝癌細胞H22的高淋巴道轉移亞系,常規傳代保種。
1.4 體外實驗
1.4.1 實驗方法[6]:(1) 標本處理:無菌條件下取小鼠腹水0.1 mL加于預盛有PBS緩沖液3 mL的離心管中,離心棄上清,用PBS緩沖液2 mL沖洗后離心3遍,加含有10%胎牛血清的RPMI1640培養基(1640全培基,下同)吹打成細胞懸液,計數并調濃度至1×106細胞/mL。(2)細胞培養:細胞懸液加入96孔培養板中(200 μL /孔),設空白對照組(1640全培基)、細胞對照組(腫瘤細胞+1640全培基)、實驗組(腫瘤細胞+含PDTC全培基)。每組設3個復孔,對照孔不加藥物。PDTC 用藥組分別加入4種不同的濃度的PDTC,從低到高依次為12.5 μg/mL、 25 μg/mL、50 μg/mL、100 μg/mL。37℃,5%CO2下培養培養24 h或8 h,每孔加入20 μL 5%MTT,繼續培養4 h,離心去上清,加入DMSO 150 μL/孔,于微量振蕩器上振蕩10 min,使藍紫色結晶完全溶解,最后放入酶標儀上以570 nm波長檢測吸光度值(absorbance,A)。重復3遍。
1.4.2 計算抑制率:按照實體瘤體外敏感標準[7],抑制率>30%為敏感,<30%為耐藥。以抑制率≥70% 為體外高度敏感,50% ≤抑制率<70% 為中度敏感,30% ≤ 抑制率<50%為低度敏感。抑制率=[(細胞對照組平均A值-實驗組平均A值)/(細胞對照組平均A值-空白對照組平均A值)]×100% 。
1.5 體內實驗
(1)動物分組:20只小鼠,雌雄各半,每只鼠右側腋下皮下接種Hca-F癌細胞1×106/0.2 mL·只,隨機分為攻擊對照組、PDTC治療組。(2)給藥方法:PDTC治療組腹腔注射0.25 g/mL PDTC溶液0.2 mL,攻擊對照組腹腔注射生理鹽水 0.1 mL/25 g。(3)指標觀察:給藥后每日測量腫瘤體積和體重,用藥結束后處死小鼠后取小鼠的腫瘤并記錄體重和瘤重。(4)去除腫瘤中的脂肪和纖維組織,采取10倍稀釋制備腫瘤組織勻漿,按照丙二醛(malondialchehyche,MDA)試劑盒說明書操作測定MDA含量,采用放射免疫法檢測腫瘤壞死因子α(tumor necrosis factor α,TNF-α)含量。
1.6 統計學方法
2 結 果
2.1 PDTC對肝癌細胞的體外增殖影響
用含有PDTC的完全培養基培養Hca-F細胞24 h或8 h后用MTT法檢測細胞的增殖活性,結果顯示PDTC對Hca-F細胞的增殖有抑制作用,濃度為25 μg/mL對Hca-F細胞的抑制作用較強, PDTC濃度作用曲線見圖1。
2.2 PDTC治療對小鼠腫瘤體積和瘤重的影響
PDTC治療接種腫瘤的小鼠10 d,每天測量腫瘤體積,活殺小鼠測瘤重,結果表明PDTC對腫瘤生長沒有明顯的抑制作用,與對照組比較差異無顯著性意義,結果見圖2和3。
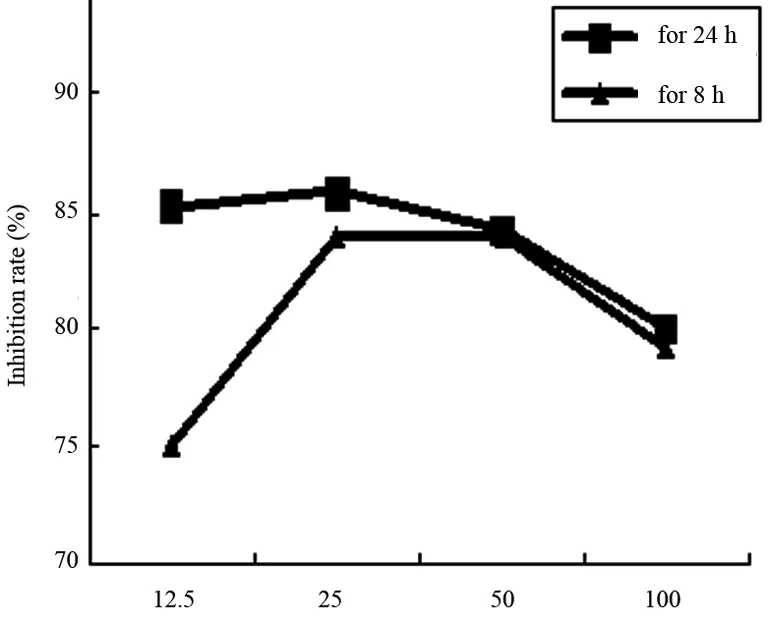
圖1 PDTC 對Hca-F細胞增殖的抑制作用
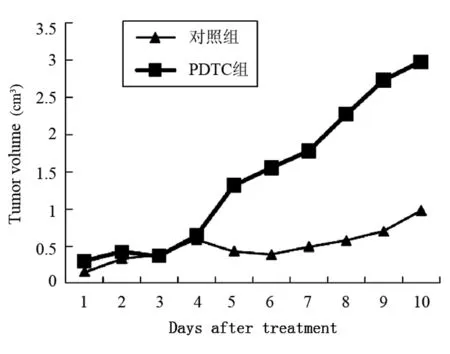
圖2 PDTC治療后Hca-F瘤體積的變化
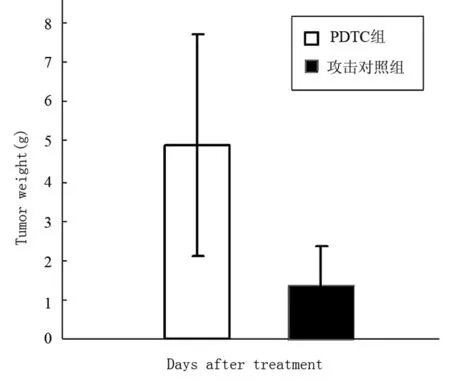
圖3 PDTC治療后Hca-F瘤重的變化
2.3 MDA檢測結果
取小鼠腫瘤組織進行MDA試劑盒檢測,PDTC組小鼠腫瘤組織勻漿MDA含量為(0.041889±0.00219)μmol/g,攻擊對照組為(0.039111±0.00388)μmol/g,兩組之間的差異無顯著性意義(P>0.05)。
2.4 TNF-α放射免疫檢測結果
注射PDTC組的小鼠腫瘤組織勻漿中TNF-α的含量(1.809±2.315) ng/mL低于生理鹽水的小鼠腫瘤組織(2.233±2.561)ng/mL,但兩組間比較,差異無顯著性意義(P>0.05)。
3 討 論
一些體內外的實驗研究表明,抑制腫瘤細胞NF-κB的活性將引起腫瘤細胞的凋亡,從而抑制腫瘤的生長,增加腫瘤細胞對放療和化療的敏感性,促進腫瘤細胞凋亡[8]。作為NF-κB的抑制劑PDTC如能應用于臨床的腫瘤治療,則能減少化療藥物的嚴重毒副作用和化療耐藥的發生率,延長化療耐藥產生時間從而進一步提高化療效果[9,10]。而本實驗為探索NF-κB的抑制劑PDTC對于某些腫瘤細胞是否的確存在抑制作用,其作用的體內外差異是否存在,這些問題對于未來的腫瘤化療有著重要的意義。
體外MTT法發現PDTC對于Hca-F的增殖有明顯的抑制作用,在濃度為25 μg/mL時抑制率明顯較高。而體內實驗以小鼠為模型,觀察瘤重及其體積可發現藥物對腫瘤并無明顯的抑制作用,反而大部分注射PDTC個體的瘤體積有增大的趨勢。MDA是細胞脂質過氧化的產物,可以反映氧化損傷的程度[11],本實驗中作為藥物療效的指標,TNF-α是NF-κB的靶基因之一,檢測腫瘤組織勻漿中TNF-α水平以反映NF-κB活化情況,在本實驗中PDTC治療組和攻擊對照組之間的差異無統計學意義,不能排除藥物劑量低的可能性,也可能因為腫瘤組織的氧化應激水平不明顯。體內抗腫瘤實驗與體外細胞毒實驗的結果差別很大,還可能與NF-κB的多功能和PDTC作用機制有關,因為PDTC是通過抗氧化作用抑制氧化應激誘導的NF-κB活化,對其他因素引起的NF-κB活化的抑制作用可能有限。NF-κB活化有其抑制腫瘤細胞凋亡和促進增殖的作用,抑制其活化與功能在體外環境中可能有抗腫瘤作用,但NF-κB活化可促進多種具有抗腫瘤作用的細胞因子產生,從而增強機體的抗腫瘤免疫,抑制NF-κB的功能對抗腫瘤免疫不利。確切機制有待深入研究。
參考文獻:
[1] Shinohara T,Miki T,Nishimura N,et al.Nuclear factor-κB-dependent expression of metastasis suppressor KAI1/CD82 gene in lung cancer cell lines expressing mutant p53[J].Cancer Res,2001,61 (2): 673-678.
[2] Bours V,Bentires-Alj M,Hellin AC,et al.Nuclear factor-κB,cancer and apoptosis[J].Biochem pharmacol,2000,60(8):1085-1090.
[3] Nuutinen U,Simelius N,Ropponen A,et al.PDTC enables type I TRAIL signaling in type II follicular lymphoma cells[J].Leukemia Res,2009,33(6):829-836.
[4] 楊春蓉,張徽,黃偉,等.PDTC聯合紫杉醇降低MDA-MB-231細胞增殖侵襲能力[J].第三軍醫大學學報,2010,32:570-579.
[5] Liu GH,Wang SR,Wang B,et al.Inhibition of nuclear factor-κB by an antioxidant enhances paclitaxel sensitivity in ovarian carcinoma cell line[J].Int J Gynecol Cancer,2006,16(5):1777-1782.
[6] 晏馬成,林厚文,沈陽,等.噻唑藍法檢測5種抗癌藥物對人癌細胞的體外抑制作用[J].中藥材,2001,24(6):418-419.
[7] 辛華雯,陳肖嘉,杜光祖,等.十種抗癌藥物敏感性試驗方法與臨床相關性研究[J].中國醫院藥學雜志,1994,14(12):548-550,576.
[8] Karin M,Cao Y,Greten FR,et al.NF-κB in cancer: from innocent bystander to major culprit[J].Nature Rev,2002,2301-2310.
[9] Liu SF,Ye X,Malik AB.Inhibition of NF-κB activation by pyrrolidine dithiocarbamate prevents in vivo expression of proinflammatory genes[J].Circulation,1999,21(7):1330-1337.
[10] Rodriguez FH,Nelson S,Kolls JK.Cytokine therapeutics for infectious diseases[J].Curr Pharm Des,2000,6(6): 665-680.
[11] Wang Q,Zou L,Liu W,et al.Inhibiting NF-κB activation and ROS production are involved in the mechanism of silibinin′s protection against D-galactose-induced senescence[J].Pharm Biochem Behav,2011,98:140-149.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
海峽科技與產業(2016年3期)2016-05-17 04:32:12
