低溫馴化過程中大青楊葉片差異蛋白質分析1)
2011-05-31 08:58:30李春明
東北林業大學學報 2011年10期
李春明
(林木育種國家工程實驗室(北京林業大學),北京,10083)
白 卉
(黑龍江省林業科學研究所)
于文喜
(黑龍江省林業科學院)
溫度是重要的環境因子之一,主要影響植物分布以及生長、發育等多個生命過程[1]。溫帶木本植物通過休眠可以避開冬季極端低溫,但早春與晚秋季節仍面臨低溫脅迫風險。通過低溫馴化增強抗寒能力是木本植物應對秋季低溫脅迫的重要方式之一。該過程主要表現為生長減緩或停止,伴隨大量活性氧的生成[2],含水量降低[3]等。另外低溫馴化過程會改變基因表達[4-7],主要表現蛋白表達上調,如抗凍蛋白[8],熱激蛋 白 (HSP)[9],穩定蛋白[10],晚期胚胎富集蛋白(LEA)[11]等。
大青楊(P.ussuriensis Kom),又稱憨大楊、哈達楊。自然分布于我國東北的長白山、小興安嶺林區,材質優良,抗寒性強,是東北三省東部山區森林更新的主要樹種之一[12]。在長期進化過程中,大青楊形成了特有的低溫適應機制,但對其低溫適應機制研究不夠深入,暫未見有關大青楊低溫分子適應機制方面的研究。蛋白質是生命活動的執行者,直接影響植物的生長發育,通過對大青楊休眠前低溫馴化過程中蛋白質差異表達的研究,尋找其低溫馴化過程中起作用的重要蛋白質,能夠從蛋白質水平研究其低溫適應機制,對全面研究大青楊抗寒機理具有重要的理論及實踐意義。
1 材料與方法
1.1 植物材料
大青楊為同一無性系,通過硬枝扦插繁殖,于2010年5月初定植于規格為20 cm×16 cm×20 cm的塑料花盆中,置于黑龍江省林業科學研究所溫室外,培養基質為V(腐殖土)∶V(河沙)=3∶1,每周澆水2次,10~15 d噴施1次10%MS溶液,培養4個月后開始試驗。2010年9月8日、9月21日、10月6日,于上午9:00時至10:00時,選擇3株長勢一致幼苗,每株取功能葉5~8片,混合后放于冰箱中-80℃保存,用于蛋白質分析。
1.2 實驗儀器
Ettan IPGphor II等電聚焦系統、Ettan DALT twelve垂直電泳系統、SE600電泳儀掃描儀、自動染膠儀(美國通用)、高速冷凍離心機(德國Sigma公司)、紫外分光光度計、Powerlook 2100XL UMAX掃描儀(臺灣)。
1.3 主要試劑
固相 pH 值 4.0~7.0,24 cm 梯度干膠條、2D-Quant試劑盒、丙烯酰胺(Acrylamide)、甲叉雙丙烯酰胺(Bis-Acrylamide),均購自 Amersham Biosciences公司。苯甲基磺酰氟(PMSF)、碘乙酰胺、二硫蘇糖醇(DTT)、N'N'N'N'-四甲基乙二胺(TEMED)購自Sigma公司;Trizol購自Invitrogen公司,Tris-base、溴酚藍、蛋白 Marker、三氯乙酸(TCA)、聚乙烯吡咯烷酮(PVPP)、牛血清白蛋白(BSA)、考馬斯亮藍R-350、低熔點瓊脂糖、β-巰基乙醇(β-ME)購自 BBI(Bio Basic Inc,Canada)。胰蛋白酶、酶抑制劑購自德國Roche公司。其它試劑為國產分析純。所有溶液均用Milli-Q制備的純水配制。
1.4 實驗方法
樣品制備:蛋白質提取方法采用三氯醋酸/丙酮沉淀法[13]。
蛋白質樣本的定量:應用Amersham Biosciences公司的2D-Quant試劑盒,進行蛋白質樣本的定量。具體操作步驟見Amersham Biosciences公司提供的雙向電泳原理和方法。
雙向電泳:①第一向等電聚焦(IEF)。初始IEF:100 V 1 h,200 V 1 h,500 V 1 h,1 000 V 1 h,1 000~8 000 V 1 h;IEF 到穩定狀態:8 000 V 10 h。②IPG膠條平衡和第二向 SDSPAGE電泳。IPG膠條平衡參照Amersham Biosciences公司提供的雙向電泳原理和方法,平衡后進行聚丙烯酰胺凝膠,將平衡好的IPG膠條用電泳緩沖液沖洗后膠面朝外小心置于第二向PAGE膠上,在膠的上面加入低熔點瓊脂糖封閉液(0.5%瓊脂糖,0.002%溴酚藍溶于SDS電泳緩沖液中),待瓊脂糖凝固后即可進行第二向SDS凝膠電泳。電泳條件見表1。
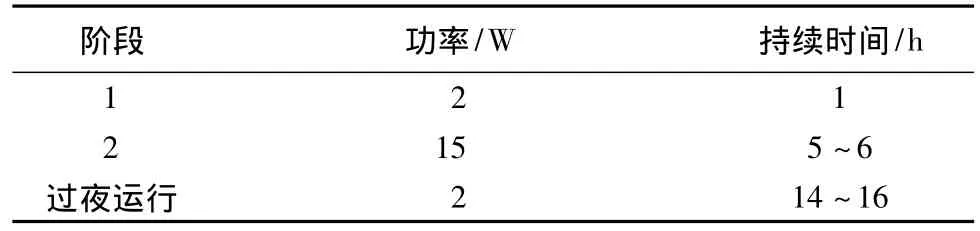
表1 第二向Ettan DALT twelve系統電泳參數
染色:采用考馬斯亮藍染色法[14]。
凝膠掃描:凝膠顯色后用UMAX 2100XL掃描儀進行圖像掃描,圖像掃描儀經強度矯正后,透射掃描2-DE凝膠(光學分辨率300像素,象素深度8bite)。
圖像分析:所得2-DE圖譜利用PDQuest V 8.0圖像分析軟件進行分析處理。
蛋白質的膠內酶解:方法參見畢影東方法[15],略有改動,加入酶液后,改水浴為37℃空氣浴倒置酶解12~14 h。
蛋白質的質譜分析:Ultraflex TM MALDI-TOF-MS質譜儀檢測,獲得蛋白點的肽指紋圖譜(PMF)信息。
數據分析及Mascot數據庫搜索:應用本地服務器上的Matrix server 2.0模塊選擇毛果楊蛋白庫(網站http://www.ncbi.nlm.nih.gov)對 PMF信息進行檢索。蛋白選擇胰蛋白酶(Trypsin),可變修飾選擇(Oxidation),固定修飾選擇(Carbamidomethyl),肽段質量標準偏差:±10-4。
蛋白質的生物信息學分析:使用本地Perl程序根據蛋白GI號從毛果楊蛋白數據庫(http://genome.jgipsf/poptrl_1/poptrl_1.home.html)中抽提目的序列。應用Blast2Go程序進行Go(gene ontology)分類。
2 結果與分析
2.1 雙向電泳圖譜
大青楊在9月8日、9月21日、10月6日葉片中蛋白質的雙向電泳圖譜見圖1,在pH值4.0~7.0的范圍內3個樣品經R-350染色后分別得到了853、988、1192個清晰可分辨的蛋白點。用分析軟件PDQuest 8.0對蛋白點進行比較,共獲得538個匹配的蛋白點,蛋白點多分布在膠的中上部。

圖1 低溫脅迫下楊樹葉片中蛋白質的雙向電泳圖譜
2.2 差異蛋白點的選擇
使用PDQuest8.0軟件,對大青楊3個時期蛋白質點的表達豐度(由軟件根據蛋白質點的灰度生成的數量值)進行差異檢測,選擇相對表達豐度大于200%的蛋白質點作為差異蛋白點,共94個(見圖2)。
2.3 差異表達蛋白點質譜鑒定結果
對94個差異蛋白點從膠上回收、酶解后進行質譜鑒定,78個蛋白點得到了完整的肽指紋圖譜,在毛果楊與植物蛋白庫中進行搜索,其中45個蛋白質得到了鑒定,結果見表2。
已鑒定的45個蛋白中有40個蛋白表達量上調2倍以上,其中有4個蛋白點(19、37、58、90)第1個時期未見表達,后兩個時期表達量大增;只有5個蛋白點(53、60、67、72、92)表達量下調2倍以上,蛋白點67在最后1個時期未見表達。另外,出現同一蛋白質出現在蛋白圖譜不同位置的現象,蛋白點 4、5、6、7,蛋白點 23、25,蛋白點 39、41,蛋白點 9、67 4 組蛋白肽指紋圖譜信息相同,但蛋白點可以通過雙向電泳分開(見表2),蛋白點39、41相對分子質量與等電點均有較大差異,其它3組蛋白相對分子質量幾乎相同,由于等電點差異而被分開(見圖2)。
2.4 蛋白功能分類
植物低溫馴化是一個復雜的生理過程,眾多蛋白參與這一過程,一個蛋白可能執行多個功能。對已鑒定的45個蛋白進行功能分類,結果表明22個(42%)是脅迫響應蛋白,12個(23%)與呼吸作用相關,9個(17%)與光合作用相關,5個(10%)參與其它生理過程,還有4個(8%)功能未知。
3 討論
3.1 利用肽指紋圖譜鑒定蛋白質
PMF是利用測量酶解肽段相對分子質量并與現有數據庫進行比對,進行蛋白質鑒定的一項技術。蛋白點4、5、6、7的PMF信息相同,相對分子質量基本相同,等電點不同,推測這4個蛋白點可能為同一蛋白經翻譯后修飾(磷酸化、甲基化、糖基化等)而造成的等電點差異。由于PMF提供的信息有限,不能對翻譯后修飾進行鑒定,所以這4個蛋白被鑒定為同一蛋白。類似情況還蛋白點23、25,蛋白點9、67。蛋白點39、41的PMF信息相同,相對分子質量、等電點均不同,推測這兩個蛋白可能是由同一基因家族控制的旁系同源基因產物,或者為同一蛋白攜帶不同信號分子或目標靶序列而形成異構體[16]。
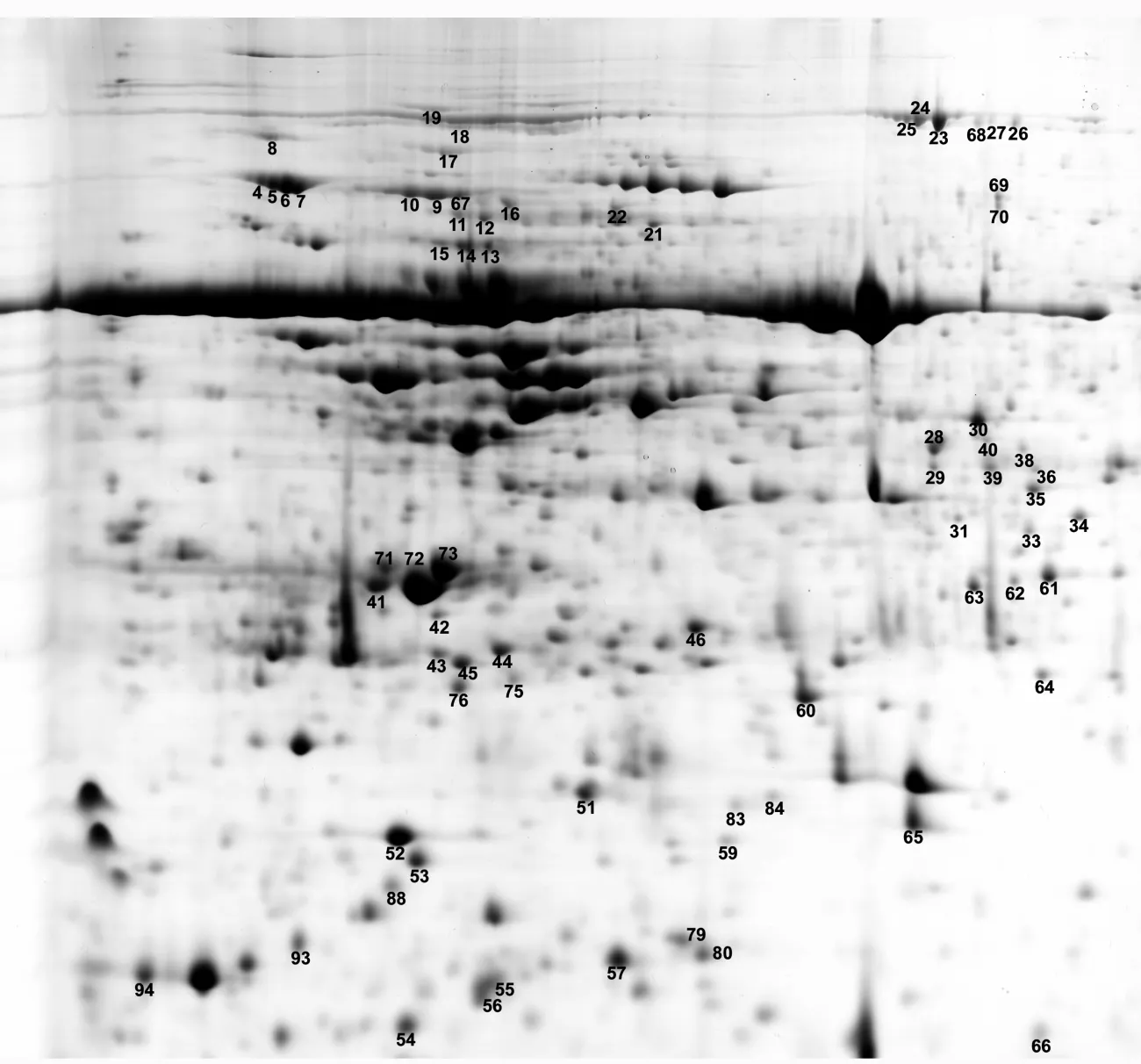
圖2 差異蛋白點在膠圖上的分布(以大青楊9月8日膠圖為例)
3.2 與脅迫相關蛋白
熱激蛋白(HSPs)是一類在生物體遭受高溫脅迫時合成的保守多肽序列蛋白質[17],又被稱為脅迫相關分子伴侶[18]。大量研究表明,低溫[19-21]、干旱、鹽漬、水淹等多種逆境脅迫也能誘導熱激蛋白的積累[22]。熱激蛋白參與變性蛋白的重新折疊,防止變性蛋白聚合,參與穩定膜系統等多個生命過程[23-24]。本研究所鑒定的45個蛋白中13個屬于熱激蛋白/分子伴侶類,10 個屬于熱激蛋白 70 家族(點 4、5、6、7、9、10、16、39、41、67),1 個屬于熱激蛋白 90 家族(點 8),2 個屬于伴侶蛋白前體(點1、45)。除點67表達量下調外,其它12個蛋白在自然低溫馴化過程中表達量均顯著上調。表明熱激蛋白/分子伴侶蛋白在大青楊低溫馴化過程中起到重要作用。
其它與脅迫響應蛋白 9 個(11、12、13、21、22、46、58、90、92),在大青楊自然低溫馴過程中,除抗病蛋白(點92)表達量下調外,其余表達量均上調,與前人研究結論基本一致。其中蛋白點21(磷酸甘油酸變位酶)[1]與蛋白點90(ATP合酶)是三羧酸循環關鍵酶[25],低溫馴化過程中,三羧酸循環關鍵酶表達量上調,會促進下游產物及ATP生成,為大青楊應對低溫脅迫提供必需的物質及能量。蛋白點46(抗壞血酸過氧化物酶)與蛋白點58(過氧化還原酶)等抗氧化酶表達量上調,有助于清除大青楊低溫脅迫下產生的過量活性氧,該結果與Renaut研究相似[20]。
3.3 與呼吸作用相關蛋白
呼吸作用是植物細胞最基本的生命活動,環境脅迫下,能量代謝加強是植物抵御逆境的基本特征。本研究中,低溫脅迫條件下共鑒定出12個與呼吸作用相關蛋白,包括溫度敏感絲狀脅迫響應蛋白(點11),細胞分裂蛋白(點12),Rubisco β亞基(點13、14),翻譯延伸因子G(點17),磷酸甘油酸變位酶(點21),羧化酶前體(點23、25),蘋果酸脫氫酶(36),脫氫酶亞基(37),酯酶D(點62),ATP合酶(點90)等在低溫脅迫下表達量都顯著上調。

表2 低溫脅迫下楊樹葉片蛋白點質譜鑒定
能量代謝是各種生命活動的基礎代謝,不僅為各種生命活動提供必需的ATP,還為眾多代謝過程提供中間代謝產物。蛋白點36為蘋果酸脫氫酶是三羧酸循環中起重要作用的酶[26],其表達豐度上調會促進能量生成及下游物質的合成,為大青楊低溫馴化及抵御低溫脅迫提供必要的能量及物質。
3.4 與光合作用相關蛋白
光合作用是綠色植物將光能固定及物質合成的過程,也是各種生命活動的起點。葉綠體是進行光合作用的細胞器,由于低溫等逆境脅迫會對葉綠體類囊體膜造成傷害,因此,必將影響植物的光合代謝,從而影響植物的正常生長發育[19]。本研究從大青楊葉片中鑒定與光合作用相關的蛋白包括溫度敏感絲狀脅迫響應蛋白(點11),細胞分裂蛋白(點12),葉綠體蛋白酶(點22),Rubisco大亞基(點52、61),葉綠體銅鋅超氧化物歧化酶(點57),葉綠體放氧蛋白(點71)等7個蛋白在自然低溫馴化過程中蛋白表達量顯著上調;ATP合酶δ鏈(點53),葉綠體放氧蛋白(點72)等2個蛋白表達量顯著下調。
Xiao等[27]研究表明,抗旱能力差的楊樹葉片中Rubisco大亞基在干旱脅迫下表達量下調。Zhao Yan等[25]發現狗牙根葉片在干旱脅迫下,葉綠素結合蛋白、葉綠體放氧蛋白、ATP合酶與Rubisco大亞基表達均下調,直接影響光合作用捕光過程、電子傳遞及碳同化3個主要過程。本研究中,只有ATP合酶δ鏈、葉綠體放氧蛋白2個蛋白表達量下調,表明大青楊在自然低溫馴化過程中光反應的光合放氧及光合磷酸化過程受到影響。
3.5 其它蛋白及功能未知蛋白
除以上3類主要蛋白外,本研究還鑒定了翻譯延伸因子G(點17),前序列蛋白酶(點19)、細胞質動力蛋白重鏈(點44)、酯酶D(點62)、抗病蛋白(點92)以及4個未知功能蛋白(34、51、60、70)。這些蛋白質在自然低溫馴化過程中表達量變化明顯,可能執行某種生物學功能。由于目前對這些蛋白功能信息了解有限,它們在大青楊自然低溫馴化中的表達變化與其在適應低溫脅迫的調節機制和功能還需要進一步的研究。
3.6 蛋白質在大青楊低溫馴化過程中的作用
大青楊是黑龍江省鄉土樹種,在長期進化過程中形成了特有低溫適應機制。通過本研究可以推測,蛋白質表達量變化與大青楊低溫馴化關系密切。大青楊在低溫及短光周期的誘導下,熱激蛋白表達量顯著上調。熱激蛋白一方面作為功能蛋白參與變性蛋白的重新折疊,防止變性蛋白聚合,穩定新生成蛋白質結構等;另外熱激蛋白也起到轉導脅迫信號并激活相關基因表達的作用。在熱激蛋白調節下,大青楊葉片中呼吸作用相關蛋白、抗氧化酶表達量上調,而參與光反應的蛋白表達量則顯著下調,進而調整生理狀態,逐步完成低溫馴化過程,為抵御低溫脅迫做好準備,見圖3。
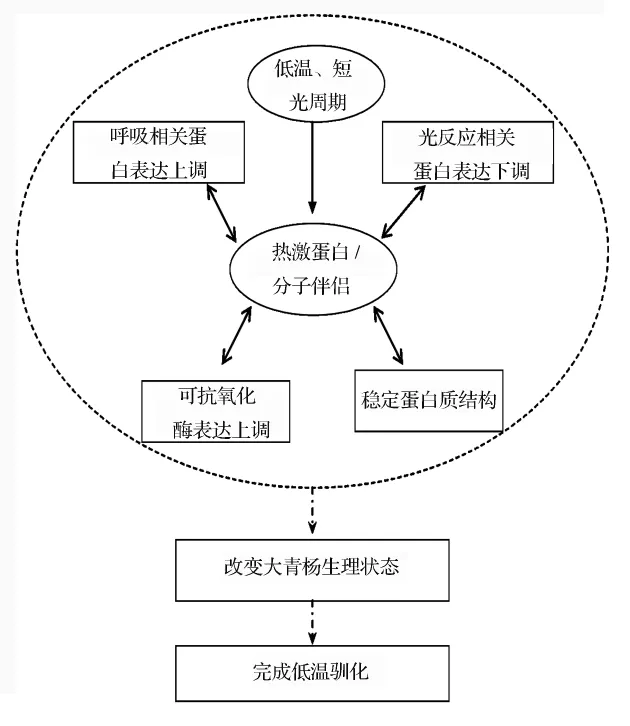
圖3 蛋白質豐度變化與大青楊低溫馴化關系
[1]J Renaut.Proteomics and low-temperature studies:bridging the gap between gene expression and metabolism[J].Physiologia Plantarum,2005,126:97-109.
[2]Prasad T K,Anderson M D,Martin B A,et al.Evidence for chilling-induced oxidative stress in maize seedlings and a regulatory role for hydrogen peroxide[J].Plant Cell,1994,6:65-74.
[3]Imanishi H T,Suzuki T,Masuda K,et al.Accumulation of raffinose and stachyose in shoot apices of Lonicera caerulea L.during cold acclimation[J].Sci Hort,1998,72:255-263.
[4]Chen jing,Dennis Doucet.Expression profile matrix of Arabidopsis transcription factor genes suggests their putative functions in response to environmental stresses[J].Plant Cell,2002,14:559 -574.
[5]Guy C.Cold acclimation and freezing stress tolerance:role of protein metabolism[J].Ann Rev Plant Physiol Plant Mol Biol,1990,41:187-223.
[6]Pearce R S.Molecular analysis of acclimation to cold[J].Plant Growth Regul,1999,29:47-76.
[7]Seki M,Narusaka M,Abe H,et al.Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray[J].Plant Cell,2001,13:61-72.
[8]Hon W C,Griffith M,Mlynarz A,et al.Antifreeze proteins in winter rye[J].Physiol Plant,1997,100:327-332.
[9]Wisniewski M E.Seasonal patterns of dehydrins and 70-kDa heatshock proteins in bark tissues of eight species of woody plants[J].Physiol Plant,1996,99:496-505.
[10]Wang W.Characterization of SP1,a stress-responsive,boiling soluble,homo-oligomeric protein from aspen[J].Plant Physiol,2002,130:865-875.
[11]Antikainen M,Griffith M.Antifreeze protein accumulation in freezing-tolerant cereals[J].Physiol Plant,1997,100:423-432.
[12]王冰,王建光,姜秀煜,等.闊葉紅松林的伴生鄉土楊樹-大青楊[J].林業科技,2003,28(6):12-15.
[13]Damerval C,de Vienne D,Zivy M,et al.Technical improvments in two-dimensional electrophoresis increase the level of genetic variation detected in wheat-seedling proteins[J].Electrophoresis,1986,7:52-54.
[14]Rabilloud T.Detecting Proteins separated by 2-D gel electrophoresis[J].Anal Chem,2000,72:48A-55A.
[15]畢影東.樟子松頂芽休眠與萌發轉換的蛋白質組學研究[D].哈爾濱:東北林業大學林學院,2010.
[16]Christophe.Mapping the proteome of poplar and application to the discovery of drought-stress responsive proteins[J].Proteomics,2006(24):6509-6527.
[17]Vierling E.The role of heat shock proteins in plants[J].Ann Rev Plant Physiol Plant Mol Biol,1991,42:579-620.
[18]Miernyk J A.The 70 kDa stress-related proteins as molecular chaperones[J].Trends Plant Sci,1997,2:180-187.
[19]Lopez Matas M A.Protein cryoprotective activity of a cytosolic small heat shock protein that accumulates constitutively in chestnut stems and is up-regulated by low and high temperatures[J].Plant Physiol,2004,132:1708-1717.
[20]Renaut J.Responses of Poplar to Chilling Temperatures:Proteomic and Physiological Aspects[J].Plant Biology,2004,6:81-90.
[21]Taylor N L,Heazlewood J L,Day D A,et al.Differential impact of environmental stresses on the pea mitochondrial proteome[J].Mol Cell Proteomics,2005,4:1122-1133.
[22]Sabehat A,Lurie S,Weiss D.Expression of small heat shock proteins at low temperatures-A possible role in protecting against chilling injuries[J].Plant Physiol,1998,117:651-658.
[23]Sung D Y,Vierling E,Guy C L.Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family[J].Plant Physiol,2001,129:789-800.
[24]Tsvetkova N.Small heat-shock proteins regulate membrane lipid polymorphism[J].Proc Natl Acad Sci,2002,99:13504-13509.
[25]Yan Zhao.Identification of proteins associated with water-deficit tolerance in C4 perennial grass species,Cynodon dactylon× Cynodon transvaalensis and Cynodon dactylon[J].Physiologia Plantarum,2010,141:40-55.
[26]Jiang Yuanqing.Comparative proteomic analysis of NaCl stress-responsive proteins in Arabidopsis roots[J].Journal of Experimental Botany,2007,58:3591-3607.
[27]Xiao X,Yang F,Zhang S,et al.Physiological and proteomic responses of two contrasting Populus cathayana populations to drought stress[J].Physiol Plant,2009,136:150-168.
