東亞飛蝗染色體C帶和銀染帶型分析
2011-04-25 09:46:22吳學芳何正波陶紅梅蔡志華
草業科學 2011年5期
關鍵詞:研究
吳學芳,何正波,陶紅梅,蔡志華
(1.重慶師范大學生命科學學院動物學重點實驗室,重慶 400047; 2.重慶師范大學資源昆蟲和分子生物研究所,重慶 400047)
近年來,我國蝗蟲災害頻繁爆發,嚴重影響了天然草地植被的正常生長發育[1],蝗蟲危害面積占草原蟲害危害面積的52.4%[2]。直翅目昆蟲東亞飛蝗(Locustamigratoriamanilensis)是一種洲際性農業害蟲,也是引發我國蝗災的最主要害蟲。目前,蝗總科的染色體已有不少的研究,但東亞飛蝗的染色體研究尚少。對東亞飛蝗染色體的研究將為生物治蝗奠定基礎[3]。蝗蟲染色體的研究普遍采用壓片染色顯帶,進行C帶和銀染核仁組織區(Ag-nucleolus organizer region,Ag-NOR)定位研究。本試驗采用哺乳動物常用的懸浮滴片法,研究了東亞飛蝗C帶和銀染顯帶,并對其帶型進行分析。
1 材料與方法
本研究所用東亞飛蝗由重慶師范大學生命科學學院資源昆蟲分子生物研究所提供。
1.1染色體標本制備 根據陶紅梅等[4]滴片法略做修改。東亞飛蝗活體注射0.04%秋水仙素溶液0.3 μL,8 h后活體解剖取出精巢。于0.75%的生理鹽水中剔凈脂肪和筋膜后,放入37℃預溫的1%檸檬酸鈉溶液中低滲30 min[5]。轉至新鮮配制的3∶1甲醇冰醋酸固定液中30 min,換固定液重復固定數次,直至精巢發白,取出剪碎,新鮮固定液懸浮固定,1 000 r/min離心,棄上清液,反復4次,滴片。
1.2C帶 C帶處理采用BSG法,具體過程參照Summer[6]方法稍加修改,玻片65℃烘箱內老化24 h,0.2 mol/L鹽酸溶液室溫處理1 h,60℃氫氧化鋇110 s,2×SSC在60℃恒溫水浴鍋內處理60 min。10%Gimesa染色8 min,鏡檢。
1.3銀染 按Howell和Black[7]的方法,略做改進。玻片于65℃水浴,加50%硝酸銀溶液4滴, 2%明膠顯影液2滴,覆以蓋片,直至玻片標本呈茶褐色(一般為3 min)。蒸餾水沖洗,10% Gimesa復染8 min,鏡檢。
1.4染色體核型 取30只蝗蟲按照上述方法制得染色體玻片標本,每個個體選取10個分散較好的分裂中期細胞顯微照相,沖洗放大。用Image-Pro Plus 5.1軟件測量照片上染色體和C帶帶紋長度等相關數據,并對染色體進行分組、編號,分析核型、C帶和銀染帶型,計算染色體的相對長度和異染色質含量[8]。
2 結果與分析
2.1核型 東亞飛蝗的染色體數目為2N(♂)=23。性別決定機制為XO。染色體組式為4L+4M+3S+XO,包括大型染色體4對,相對長度RL值13.99~11.11;中型染色體4對,相對長度RL值8.16~5.16;小型染色體3對,相對長度RL值3.22~2.44;X染色體為大型染色體,RL值17.37,染色體長度在整個染色體組中位居第一(表1)。本試驗顯示的染色體核型和組式與賈瀟凌和馬恩波[9]、馬恩波和歐曉紅[10]的研究結果一致,可確定此為東亞飛蝗的基本核型。

表1 東亞飛蝗染色體和C帶數據統計
2.2C帶 東亞飛蝗所有染色體具有明顯的端著絲粒帶,與已進行染色體研究的大多數蝗總科種類一致。早前期細胞著絲粒深染點附著在細胞核膜上,減數分裂Ⅰ和減數分裂Ⅱ中期染色體都具有著絲粒C帶。其中,第1、4、7對和X染色體著絲粒較淺,僅隱約可見;第2、3、5、6、9、10、11對染色體著絲粒呈現大小不同的深染區;第8對染色體著絲粒為中度著色,第2對染色體端粒深染;未見居間帶(圖1)。染色體組中異染色質總含量為(15.88±0.71)%。
2.3銀染 間期細胞銀染深染點為3~6個不等,多為5個,有的深染點緊貼于細胞核膜上。減數分裂Ⅰ前期觀察到黑色團塊Ag-NOR,多為3個。減數分裂Ⅰ中期,觀察到2號染色體近端部次縊痕處呈較規則圓形Ag-NOR,較為恒定。極少數細胞除2號染色體的深染點外,1號染色體近著絲粒初縊痕處有形狀不規則Ag-NOR。 其他各期未發現明顯的Ag-NOR(圖2)。
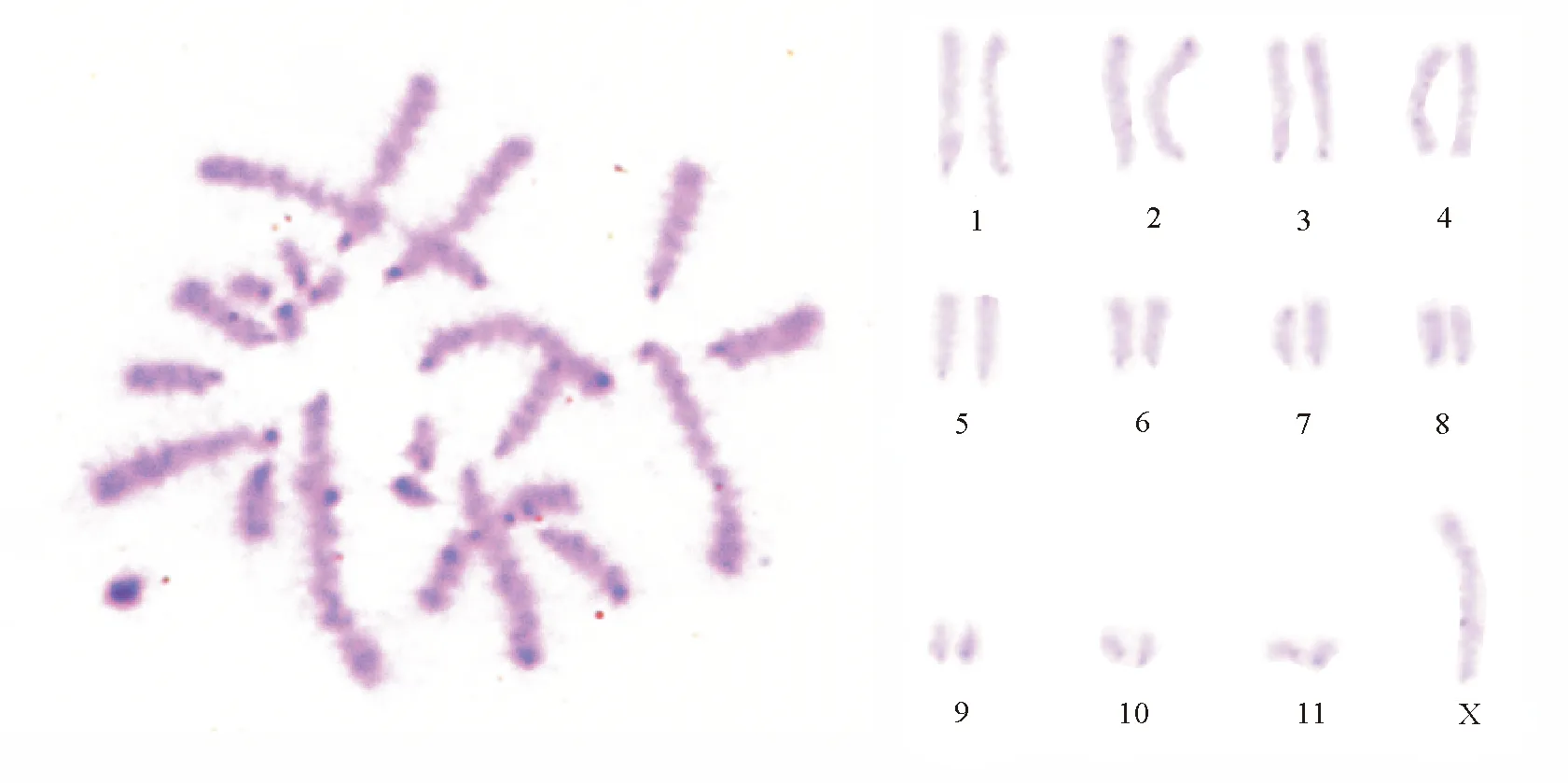
圖1 東亞飛蝗中期染色體C帶及排序

圖2 東亞飛蝗中期銀染及排序
3 討論與結論
以往蝗蟲染色體研究多采用冰凍揭片法或壓片法,本試驗采用改進后的懸浮滴片法[4],玻片較其他方法更為干凈,染色體形態良好,細胞質抽提干凈,利于觀察和顯帶研究。
制片過程中,秋水仙素的注射量和低滲時間是影響東亞飛蝗染色體標本形態主要因素。秋水仙素注射量過大,染色體凝縮,甚至可呈點狀;注射量過小,則無法起到阻斷細胞分裂的作用,中期細胞少。經反復試驗,每只蟲體注射3 μL 0.04%秋水仙素,處理8 h,效果最好。低滲時間也會影響染色體形態。低滲時間過短,染色體伸展不好,呈緊縮狀;時間過長,染色體疏松,崩解,甚至出現燈刷染色狀外觀,若繼續低滲會導致細胞破裂,染色體丟失。1%檸檬酸鈉溶液低滲30 min可達到較好的效果。
3.1C帶 近年來,人們利用分子生物學研究手段證實C帶主要是AT與GC高度重復序列[11]。C帶標本中深染部分即為結構異染色質部分,著色深淺程度與異染色質含量高低有直接聯系,可因此進行種間的比較。根據Webb[12]的觀點,深黑色為結構異染色質密集區域,灰黑色帶則為結構異染色質與常染色體夾雜的區域。第2對染色體端粒處出現深染,說明較其他染色體,該染色體端粒處結構緊密且非組蛋白含量也更為豐富。東亞飛蝗均為端部著絲粒染色體,且某些染色體出現了端帶,而以往研究的大多數蝗總科[9,12-15]的種類也出現了此特點。對比發現,東亞飛蝗與其他種類C帶差異表現在端帶、居間帶的有無,個別染色體上端帶的大小、著色強弱,結構異染色質含量多少等方面。
從遺傳學角度來看,C帶所顯示的結構異染色質區域在遺傳上轉錄活性低,在種內水平上具有一定穩定性,低級階元分類上是一個有用的指標[11,16]。但與銀染帶型相比,C 帶的進化速率較快,往往在相近種間C 帶異染色質的結構及組成都具有差異[13]。就異染色質含量來看,進行C帶研究的蝗蟲異染色質含量普遍在20%左右[8,11,13-17]。而姚世鴻[13]研究的同科的疣蝗(Trilophidiaannulata)異染色質含量為21.15%,韋仕珍和鄧維安[17]研究的隆叉小車蝗(Oedaleusabruptus)和紅脛小車蝗(O.manjius)異染色質含量分別為22.36%和22.07%,相比之下,東亞飛蝗的含量較低,僅為15.88%,與傅鵬和鄭哲民[8]研究的癩蝗科友誼華癩蝗(Sinotmethisamicus)含量(15.18%)較為接近。說明異染色質含量在同科不同種間含量存在較大差異。一般情況下,C帶處理的結果往往只能顯示一個物種或種群結構異染色質主要的分布和含量特征[18]。而賈瀟凌和馬恩波[9]的東亞飛蝗C帶結果顯示其異染色質含量為18.87%,未見端帶。根據Max和Bernard[19]報道,同物種的C帶帶型會隨著如酸堿處理時間不同和染色體伸展程度等因素的影響而變化。因此,只有在同一細胞分裂時期并在基本相同的分帶處理條件下才可能得出較為一致的結果。
3.2銀染核仁組織區 銀染是中期染色體的核仁組織區(NOR),Howell[20]和Hsu等[21]的研究顯示,深染區域是具有轉錄活性的18S+28S rDNA。因此理論上深染點應該為偶數,但是本試驗卻觀察到深染點大多呈奇數,且僅在同源染色體的其中1條上出現,陳曉光和何麟[22]在研究褐斑大蠊(Periplanetabrunnea)時也出現此現象。可推測東亞飛蝗只有一條18S+28S rDNA基因功能上有活性。間期及早前期深染點多,到減數分裂Ⅰ中期,只可見1處或2處深染點,可見,rDNA轉錄活性降低。減數分裂Ⅰ后期、末期以及減數分裂Ⅱ未見深染。2處深染的情況并不多見,另一處出現在1號染色體近著絲粒端的深染可能僅是2號染色體rDNA未充分表達時,作為補償性的開放。此種核仁組織區的多態性,在以往研究的多種動物中也有發現,如靈長類[23]、兩棲類[24]、魚類[25]等。18S+28S rDNA轉錄酸性蛋白,而酸性蛋白又與染料中銀離子特異性結合而著色[26]。
與C帶、G帶所不同的是,核仁組織區的定位以及其活性,具有較慢的進化速率和相對較大的保守性,一般較為恒定。正是由于核仁組織區定位在遺傳學上的這種特點,使得該項技術成為揭示一個大分類單元(高級階元)中種系發生關系的一個有用指標[27]。 本研究的結果顯示,東亞飛蝗的核仁組織區定位于2號染色體端部次縊痕處(圖2),經反復試驗,發現其具有恒定特點。而賈瀟凌和馬恩波[9]研究的東亞飛蝗銀染定位在M8上,產生差異的原因尚不明白,需做進一步研究。
馬恩波和郭亞平[14]在研究斑腿蝗科發現銀染核仁組織區與C帶有相關性,出現Ag深染區域也有C帶帶紋,而在稻蝗屬、卵翅蝗屬、偽稻蝗屬[28]中也發現此對應關系。但是本試驗的結果卻未顯示出這種關聯,東亞飛蝗的核仁組織區位于2號染色體靠近端粒區域,C帶在次縊痕區域未有帶型,僅出現端粒深染的情況。在有關飛蝗屬的文獻中也未發現有此種聯系,這種聯系并不存在于飛蝗屬中。
東亞飛蝗核仁組織區定位于2號染色體,且2號染色體的C帶帶型豐富,因此可作為東亞飛蝗的遺傳標記。
[1] 孫濤,趙景學,田莉華.草地蝗蟲發生原因及可持續管理對策[J].草業學報,2010,19(3):220-227.
[2] 王俊梅.生物技術對草原蝗蟲的控制效果及應用前景[J].草業科學,2009,26(9):206-211.
[3] 王俊梅,豆衛,譚成虎.綠韁菌復合油劑對草原蝗蟲的防治試驗[J].草業科學,2009,26(10):155-159.
[4] 陶紅梅,蔡志華,何正波,等.東亞飛蝗染色體的減數分裂觀察[J].重慶師范大學學報(自然科學版),2010,27(2):20-22.
[5] 趙建,何孟元,郝水.紅翅皺膝蝗減數分裂染色體的螺旋與軸結構[J].實驗生物學報,1990,23(3):261-271.
[6] Summer A T.A simple technique for demonstrating centromeric heterochromatin[J].Experimental Cell Research,1972,75:304-306.
[7] Howell W M,Black D A.Controlled silver-staining of nucleolus organizer regions with a protective colloidal developer: a 1-step method[J].Experientia,1980,36:1014-1015.
[8] 傅鵬,鄭哲民.兩種華癩蝗的染色體C-帶核型[J].遺傳,1989,11(3):26-28.
[9] 賈瀟凌,馬恩波.斑翅蝗科部分種屬細胞分類學的研究[J].山西大學學報(自然科學版),1998,21(3):291-296.
[10] 馬恩波,歐曉紅.蝗總科染色體研究及科級綜合比較(直翅目)[J].昆蟲分類學報,2002,22(1):6-10.
[11] 李新江,張道川,王文強.蝗蟲染色體C-帶核型研究進展(昆蟲綱:直翅目)[J].河北大學學報(自然科學版),2005,24(3):332-336.
[12] Webb G C.Cytogenetics of the parthenogenetic grosshopper warrmba (formerly noraba) virgo and its bisexual relatives.Ⅳ Chromosome banding studies[J].Chromosoma,1980,67:309-339.
[13] 姚世鴻.貴州蝗總科5種蝗蟲的核型和C-帶[J].貴州師范大學學報,2006,24(2):18-22.
[14] 馬恩波,郭亞平.四種斑腿蝗科昆蟲染色體帶型的比較[J].昆蟲學報,2001,44(3):268-275.
[15] 馬恩波,鄭哲民.五種稻蝗染色體核型和C帶帶型的比較[J].昆蟲學報,1989,32(4):399-404.
[16] 霍滿鵬,王文強,蒲力群,等.菊蝗和黃佛蝗三個種染色體C帶帶型分析[J].陜西師范大學學報(自然科學版),1997,25:48-52.
[17] 韋仕珍,鄧維安.兩種小車蝗染色體C帶核型研究(蝗總科:斑翅蝗科)[J].四川動物,2009,28(1):7-10.
[18] Sants J L,Giraldz R.C-heterochromatin polymorphism and variation in chiasma localization inEuchorthippuspulvinatusgallicus(Acrididae,Orthoptera)[J].Chromosoma,1982,85:507-508.
[19] Max K,Bernard J.Regularities and restrictions governing C-band variation in acridoid grasshoppers[J].Chromosoma,1980,76:123-150.
[20] Howell W M.Chromosome core structure revealed by sliver staining[J].Chromosoma,1979,73:61-66.
[21] Hsu T C,Spirito S E, Pardue M L.The distribution of 18+28S ribosomal gene in mammalian Chromosomes[J].Chromosoma,1975,53(1):25-36.
[22] 陳曉光,何麟.褐斑大蠊的銀染核仁形成區及染色體骨架研究[J].第一軍醫大學學報,1988,8(4):343-339.
[23] 曹莜梅,陳宜峰,羅麗華.十一種靈長類染色體核仁組織者(NORs)的比較研究[J].動物學研究,1983,2(2):119-123.
[24] 尚克岡,李士鵬,金昊.花背蟾蜍的核型分析Ⅱ:銀染核仁組成區在數目上的多態及在劑量上的補償效應[J].遺傳學報,1984,11(1):52-60.
[25] 王蕊芳,施立明,賀維顧.幾種鯉魚染色體核仁組織者的銀染觀察[J].動物學研究,1985,6(4):391-398.
[26] 馬恩波,郭亞平,張冰.九種蝗蟲核仁組成區定位及細胞分類學意義[J].山西大學學報(自然科學版),1993,16(2):217-222.
[27] 馬恩波,郭亞平,鄭哲民.斑腿蝗科10屬NOR定位及其細胞分類學意義[J].動物學研究,1992,14(3):271-276.
[28] 馬恩波,郭亞平.偽稻蝗及其近緣屬的細胞學分類[J].昆蟲學報,1995,38(1):54-60.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
