川西北不同退化程度高山草甸和亞高山草甸的植被特征
2011-04-25 09:41:48干友民羅元佳馮國英
草業科學 2011年6期
關鍵詞:物種
紀 磊,干友民,羅元佳,成 平,馮國英,馬 超
(四川農業大學草業科學系,四川 雅安 625014)
青藏高原是我國主要的畜牧業基地之一,草地資源豐富。然而,目前青藏高原草地退化嚴重,據統計,約有0.45×108hm2退化草地,約占青藏高原草地面積的1/3[1]。川西北地區處于青藏高原東南緣,屬于海拔較高地區,高山草甸與亞高山草甸是該地區主要的植被類型之一,兩者普遍處于放牧退化過程中,而植被退化先于土壤退化,認識放牧干擾下草地退化的植被特征是采取合理的管理措施以防止草地退化、保證草地畜牧業可持續發展的基本前提。本研究分別以四川甘孜州理塘縣境內的高山草甸與阿壩州紅原縣境內的亞高山草甸退化草地為研究對象,對其退化演替過程中的植被特征進行了對比研究,以揭示兩者退化過程中植被特征變化的相似性與相異性。
1 材料與方法
1.1研究區概況 高山草甸退化草地研究區選在四川省甘孜藏族自治州理塘縣境內。地理位置為29°54′~30°14′ N, 99°52′~100°21′ E,平均海拔4 020 m,屬高原寒溫帶季風氣候,多年平均氣溫3 ℃,最高26.8 ℃,最低-30.5 ℃;年平均降水量725.8 mm;相對濕度70%;年平均日照時數2 500~2 650 h。土壤以高山草甸土和亞高山草甸土為主。高山嵩草(Kobresiapygmaea)和四川嵩草(K.setchwanensis)為植被群落主要建群種,伴生種有羊茅(Festucaovina)、西藏嵩草(K.tibetica)、異針茅(Stipaaliena)、垂穗披堿草(Elymusnutans)等[2]。
亞高山草甸退化草地研究區選在四川省阿壩藏族羌族自治州紅原縣境內。地理位置為31°51′~33°19′ N,101°51′~103°23′ E,平均海拔3 500 m左右。屬大陸性高原溫帶季風氣候,年均溫為0.6~1.1 ℃,年均降水量為650~730 mm,≥10 ℃年積溫865 ℃·d。土壤主要為亞高山草甸土和草甸潮土,植被類型主要為亞高山草甸,主要優勢植物有垂穗披堿草、四川嵩草、西藏嵩草、珠芽蓼(Polygonumviviparnm)等[2]。
1.2研究方法
1.2.1植被調查 樣地設置采用時空替換法,即利用空間的變換來代替時間的變換,用空間上不同草地放牧退化程度來研究草地在時間上的退化序列[3-4]。通過實地走訪與調查,根據不同放牧強度,結合紅原縣與理塘縣國家天然草地退牧還草工程點位置、歷史調查資料、牧民居住點位置以及草地放牧時間等因素,在高山草甸與亞高山草甸內分別按照植被蓋度及地表裸露程度選取輕度退化(light degradation,LD,地表少量裸露,植被蓋度75%~90%)、中度退化(medium degradation,MD,大量裸斑,植被蓋度65%~75%)、重度退化(heavy degradation,HD,裸地或少量毒害草,植被蓋度<60%)及無明顯退化(undegraded,UD,幾乎無裸斑,植被蓋度>90%)4個夏季天然放牧草地作為研究樣地,其中無明顯退化樣地從圍欄5年以上至今無放牧干擾的草地內選取,各樣地面積大于1 hm2。各樣地遠離公路,以減小交通運輸對草地的干擾。
2009年7月采用樣方法在上述各個樣地內分別隨機選擇5個樣方(1 m×1 m)進行植物群落調查,共計130個樣方,調查指標包括樣方內所有植物種分種蓋度、高度、多度、分種地上現存量和頻度,以及群落的高度和蓋度。
1.2.2數據處理 根據測定結果計算物種優勢度:SDR5=(相對高度+相對多度+相對蓋度+相對頻度+相對質量)/5。
生物多樣性指數的計算包括:
S=n
(1)
物種多樣性指數(Shannon-Wiener Index):
(2)
均勻度指數(Pielou Index)[5]:
(3)
式中,n為出現在樣方中的物種數目,Pi為第i個種的相對優勢度(SDR5i/100)。
以上數據的統計分析采用Excel和SPSS軟件,圖中數據結果均以平均數±標準差表示。
2 結果與分析
2.1物種豐富度與優勢種變化 不同退化程度的高山草甸草地物種豐富度變化從無明顯退化(UD)到重度退化(HD)呈高-低-高趨勢,與亞高山草甸(低-高-低)呈相反趨勢,說明放牧干擾對兩類退化草地物種豐富度的影響具有相異性(圖1)。高山草甸退化演替上草地豐富度值變化范圍為9.56~15.00種/m2(輕度退化最低),亞高山草甸變化范圍為11.80~25.08種/m2(輕度退化最高)。草地群落則由高大叢生的嵩草群落向低矮耐牧的匍匐型、蓮座狀植物組成的雜類草群落演變,其間以直立多年生雜類草群落為過渡。隨著放牧強度增加,高山草甸某些物種如四川嵩草從群落中消失,而另一些植物,如高山嵩草則通過形態變化或耐踐踏適應放牧等變化而得以生存。

圖1 不同退化程度的物種豐富度變化
高山草甸與亞高山草甸退化草地群落優勢種存在一定差異,高山嵩草在高山草甸退化草地大量出現,在亞高山草甸退化草地則較少出現;優良牧草老芒麥在高山草甸較少出現;雜類草種類及優勢度亦存在差別(表1)。
2.2物種多樣性及均勻度變化 高山草甸退化草地物種Shannon-Wiener多樣性指數變化不顯著,僅無明顯退化(2.49±0.02)與中度退化(2.35±0.10)間差異顯著(P<0.05);亞高山草甸退化草地物種多樣性變化呈“拋物線”趨勢,說明放牧干擾對高山草甸多樣性影響較小,而對亞高山草甸影響較顯著(表2)。
高山草甸均勻度指數(Pielou)變化無規律性,最大值出現在中度退化(0.95±0.03),而輕度退化草地(0.94)與中度退化草地的群落均勻度(0.92)較高,說明植被優勢度分配較均勻;亞高山草甸均勻度變化顯著,無明顯退化到重度退化呈明顯上升趨勢,說明放牧對高山草甸均勻度影響不及亞高山草甸的顯著(表2)。
2.3植被蓋度變化 高山草甸與亞高山草甸放牧退化草地蓋度均隨退化程度增加而下降。高山草甸中,重度退化與無明顯退化相比蓋度下降了32%,但重度退化蓋度仍然達64.00%;亞高山草甸蓋度變化梯度不明顯,輕度退化(96.00%)與中度退化(83.50%)和重度退化(88.50%)差異極顯著(P<0.01)(表2)。
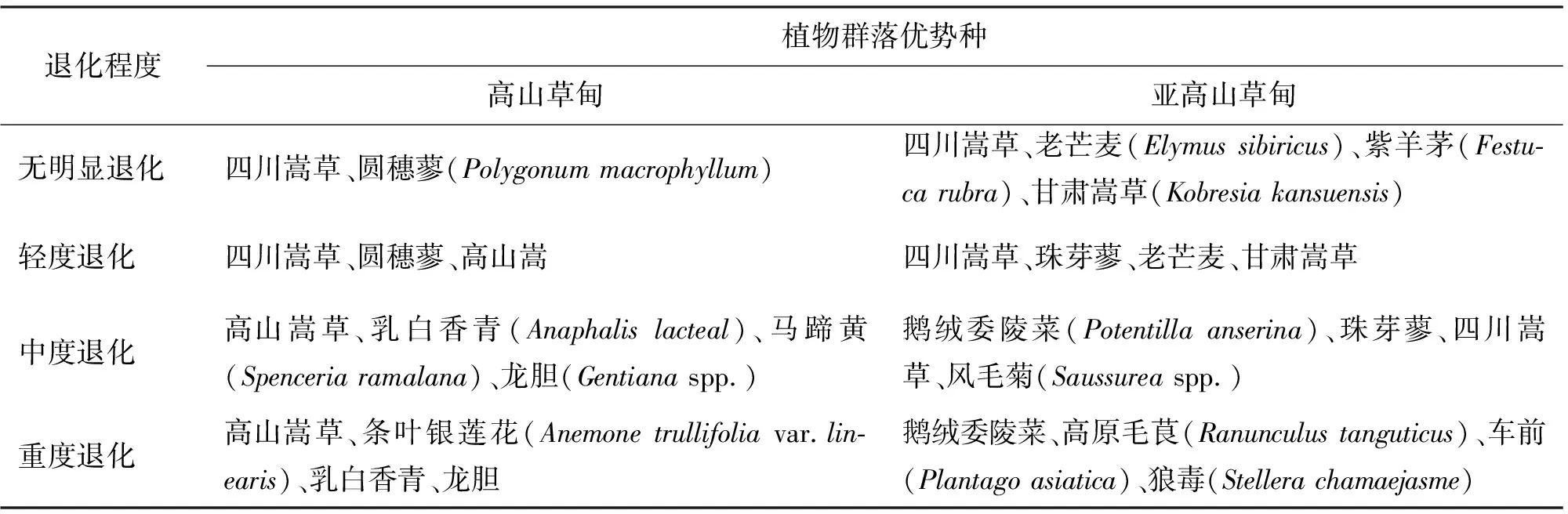
表1 川西北高山草甸與亞高山草甸不同退化程度草地優勢種
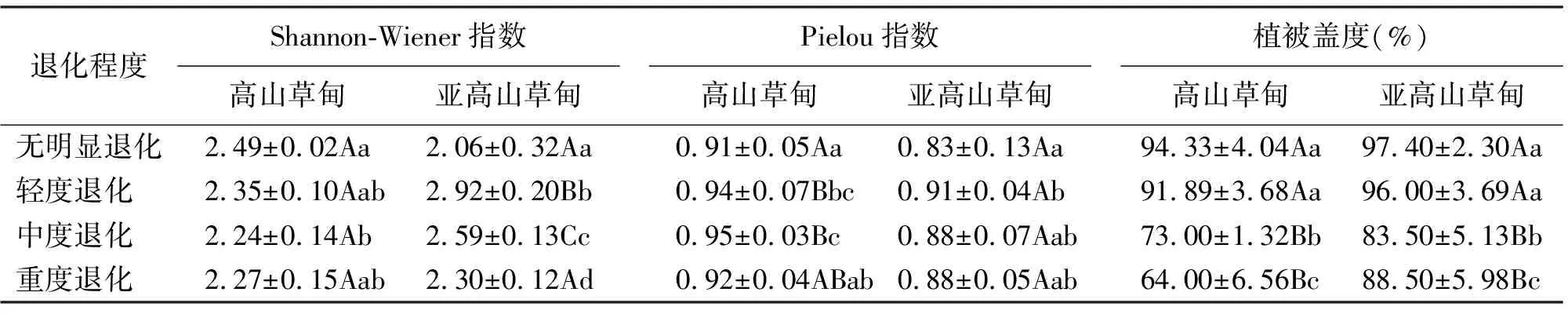
表2 不同退化程度草地Shannon-Wiener指數、Pielou指數和蓋度變化
2.4植被高度變化 高山草甸與亞高山草甸放牧退化草地植被高度隨退化程度加劇,呈下降趨勢。在無明顯退化階段的最高。高山草甸植被高度無明顯退化(14.0 cm)與中度退化(6.0 cm)和重度退化(5.0 cm)差異均極顯著(P<0.01),而與輕度退化差異不顯著(P>0.05),重度退化較無明顯退化下降了72%,處于圍欄封育(無明顯退化)的草地最高;亞高山草甸差異達極顯著(P<0.01),僅中度退化(10.0 cm)與重度退化(4.0 cm)差異不顯著(P>0.05),結果均說明放牧干擾降低了草地群落高度。高山草甸高度變化為3.98~13.98 cm,而亞高山草甸為3.26~41.42 cm(圖2)。
2.5地上現存量變化 放牧干擾不同程度地使地上現存量減少,除無明顯退化草地外,在輕度退化階段的地上現存量最高。高山草甸退化草地與亞高山草甸退化草地有相同變化規律,但亞高山草甸地上現存量的下降趨勢明顯高于高山草甸。亞高山草甸重度退化(236.06 g/m2)較無明顯退化(1 151.20 g/m2)的降幅達到79%,而處于圍欄封育(無明顯退化)的草地現存量最高,說明圍欄封育有利于現存量的積累,同時亞高山草甸地上現存量均高于高山草甸(圖3)。
2.6有毒有害植物地上現存量占群落地上現存量的比例 高山草甸有毒有害植物地上現存量占群落地上現存量的比例呈上升趨勢,差異均未達到極顯著水平(P>0.01),無明顯退化(15.97%)、輕度退化(16.37%)分別與中度退化(27.27%)、重度退化(28.40%)差異顯著(P<0.05);亞高山草甸有毒有害植物地上現存量占群落地上現存量的比例變化規律性不明顯,無明顯退化中比例最小,僅為0.37%,說明圍欄對草地優良物種的恢復起到了明顯作用,同時抑制了有毒有害植物的生長,降低了地上現存量(表3)。
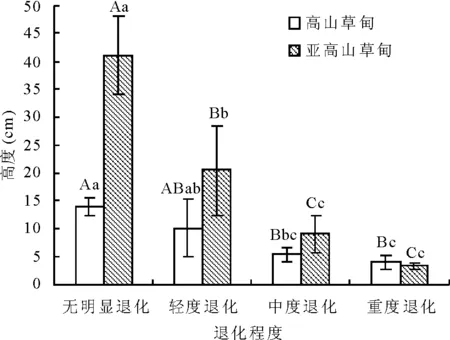
圖2 不同退化程度的植被高度變化

圖3 不同退化程度的植被地上現存量變化
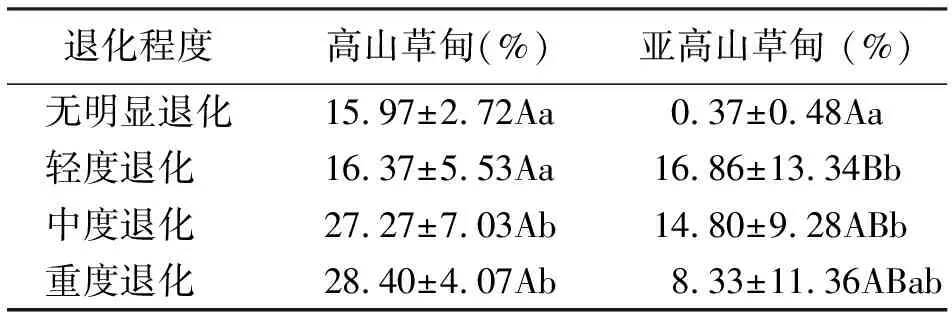
表3 有毒有害植物地上現存量比例變化
3 討論
在相同退化階段高山草甸草地的地上生物量和植被蓋度均低于亞高山草甸。造成這種差異主要原因是環境條件、植被類型與放牧歷史的不同。如亞高山草甸退化草地的植被高度變化極顯著的原因是禾本科植物占的比例較大且耐牧性較差。這些差異表明川西北退化草地植被變化特征與其他青藏高原地區的同類研究具有相似性[6-7]。此結果與公延明等[8]對巴音布魯克高寒草地退化演替階段植物群落特性研究分析結果相近。
隨著退化程度加劇,高山草甸植被蓋度呈下降趨勢,而亞高山草甸植被蓋度的變化規律性差,區分度較低。隨著放牧退化程度加劇使群落優勢植物種發生更替,耐牧植物增多,而有毒植物條葉銀蓮花、龍膽、狼毒以及乳漿大戟(Euphorbiaesula)等的大量出現擠占了其他優良牧草生存空間,降低草地質量,物種“小型化”現象明顯[9-10]。這種“小型化”現象與雜類草(如:龍膽)等蓮座狀植物的投影面積較大覆蓋地表的特點有關,說明植物種類不同直接影響草地植被蓋度的變化,而草地退化對蓋度的影響亦由于地區與植被類型差異而發生變化。
亞高山草甸草地的物種豐富度及植被分布均勻度均高于高山草甸草地。在高山草甸草地無明顯退化的物種多樣性最高則可能與用來計算物種多樣性的Shannon-Wienner指數有關,由于該指數為信息指數,以植物種優勢度為公式計算的來源,若某些植物種的優勢度較高則會使物種多樣性指數顯著升高。而無明顯退化中,由于優勢種如四川嵩草、圓穗蓼等植物占據了大量的群落優勢度,所以無明顯退化的物種多樣性指數值均高于其他退化草地;而輕度退化對亞高山草甸具有最高的物種多樣性,能促進草地植物群落的更新與穩定,該強度的放牧干擾有利于保持群落穩定性并提高群落生產力[11],與“中度干擾理論”的相關研究結果具有一致性[12]。
高山草甸退化有毒有害植物地上現存量占群落地上現存量比例高于亞高山草甸,其增加趨勢是由于家畜對其采食量的下降造成的,這與Lym和Kirby[13]報道乳漿大戟的出現可引起可食草產量下降75%以上而使其所占比例上升的結果一致。
研究區草地生態系統普遍受到人為活動干擾,偏離正常狀態[14-15]。對輕度退化草地,建議在合理利用和保護的前提下,維持其生態功能;對中度和重度退化草地,則在條件允許下盡量減少放牧活動干擾,以恢復并提高其生態功能。
4 結論
不同退化程度下的川西北高山草甸與亞高山草甸草地物種豐富度變化呈相反趨勢,說明放牧對2類退化草地物種豐富度的影響具有相異性。草地群落優勢種存在一定差異,高山嵩草在高山草甸退化草地大量出現而優良牧草老芒麥則較少出現,高山嵩草在亞高山草甸則出現較少。蓋度和高度在高山草甸和亞高山草甸上均隨放牧強度的增大而下降。高山草甸有毒有害植物地上現存量占群落地上現存量的比例呈上升趨勢且比亞高山草甸的比例高。
[1]馬玉壽,郎百寧,王啟基.“黑土型”退化草地研究工作的回顧與展望[J].草業科學,1999,16(2):5-9.
[2]周壽榮.四川草地資源[M].成都:四川民族出版社,1998:141-153.
[3]邱英,干友民,王欽,等.川西北放牧草地退化分類評價指標體系初探[J].湖北農業科學,2007,46(5):723-726.
[4]任繼周.草業科學研究方法[M].北京:中國農業出版社,1998:207-211.
[5]Hutchings M J,de Kroon H.Foraging in plants: the role of morphological plasticity in resource acquisition[J].Advanced Ecology Research,1994,25:159-238.
[6]周華坤,趙新全,周立,等.青藏高原高寒草甸的植被退化與土壤退化特征研究[J].草業學報,2005,14(3):31-40.
[7]汪詩平.青海省“三江源”地區植被退化原因及其保護策略[J].草業學報,2003,12(6):1-9.
[8]公延明,胡玉坤,阿德力·麥地,等.巴音布魯克高寒草地退化演替階段植物群落特性研究[J].干旱區資源與環境,2010,24(6):149-152.
[9]干友民,李志丹,王欽,等.川西北亞高山草甸放牧退化演替研究[J].草地學報,2005,13(S1):48-52.
[10]韓友吉,陳桂琛,周國英,等.青海湖地區高寒草原植物個體特征對放牧的響應[J].中國科學院研究生院學報,2006,23(1):118-122.
[11]王德利,祝廷成.不同種群密度狀態下羊草地上部生態場、生態勢、場梯度及其季節性變化規律研究[J].生態學報,1996,16(2):121-127.
[12]寶音陶格濤,劉美玲,李曉蘭.退化羊草草原在淺耕翻處理后植物群落演替動態研究[J].植物生態學報,2003,27(2):270-277.
[13]Lym R G,Kirby D R.Cattle foraging behavior in leafy spurge (Euphorbiaesula)-infested rangeland[J].Weed Technology,1987,1:314-318.
[14]王巖春,干友民,費道平,等.川西北退牧還草工程區圍欄草地植被恢復效果的研究[J].草業科學,2008,25(10):18-19.
[15]干友民,羅元佳,周家福,等.川西北沙化草地生態恢復工程對沙地植被群落的影響[J].草業科學,2009,26(6):51-56.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
學生天地(2019年36期)2019-08-25 08:59:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
