不同活力種子的萌發與出苗特性
2011-04-25 09:43:04胡小文何學青王彥榮
草業科學 2011年6期
王 娟,胡小文,何學青,王彥榮
(蘭州大學草地農業科技學院,甘肅 蘭州 730020)
北美官方種子分析家協會(AOSA)于1980年指出,種子活力是指在廣泛的田間條件下,決定種子迅速整齊出苗和長成正常幼苗潛在能力的總稱。作為評價種子質量的重要指標,活力與種子田間出苗狀況[1]、植物的抗逆、早熟、豐產特性密切相關[2]。 因而,正確評價種子活力對生產實踐具有重要意義。
標準發芽試驗是評估種子活力的一個重要方法[3],但是有關標準發芽試驗與田間出苗相關性的研究相對較少,且多數研究一般僅考慮萌發有關指標與田間出苗率的關系,對其他田間參數如出苗速率、幼苗生物量等關注不多。本研究通過對幾種禾本科牧草種子進行老化處理,獲得不同活力的種批后,測定其萌發(發芽率、發芽勢、發芽指數)、幼苗(胚根、胚芽、幼苗長度)以及盆栽條件下的出苗相關指標(出苗率、出苗速率),進一步對上述指標進行相關分析,探討標準發芽試驗測定活力方法用于評價種批田間表現的可行性。
1 材料與方法
1.1材料 供試種子燕麥(Avenasativa)、蘇丹草(Sorghumsudanense)、垂穗披堿草(Elymusnutans)、羊草(Leymuschinensis)、高羊茅(Festucaarundinacea)和多年生黑麥草(Loliumperenne)由農業部牧草與草坪草種子質檢中心(蘭州)提供,試驗前各樣品貯藏于4 ℃待用,樣品詳細信息見表1。
1.2方法
1.2.1老化處理 從各供試種子中隨機選取所需足夠數量的種子分為56份(每份50粒種子),參照王彥榮等[4]的方法調節種子含水量到20%,鋁箔袋包裝密封,置于42 ℃恒溫培養箱內,分別處理0、12、24、36、48、60、84 h 后取出,室溫條件晾干,待用。
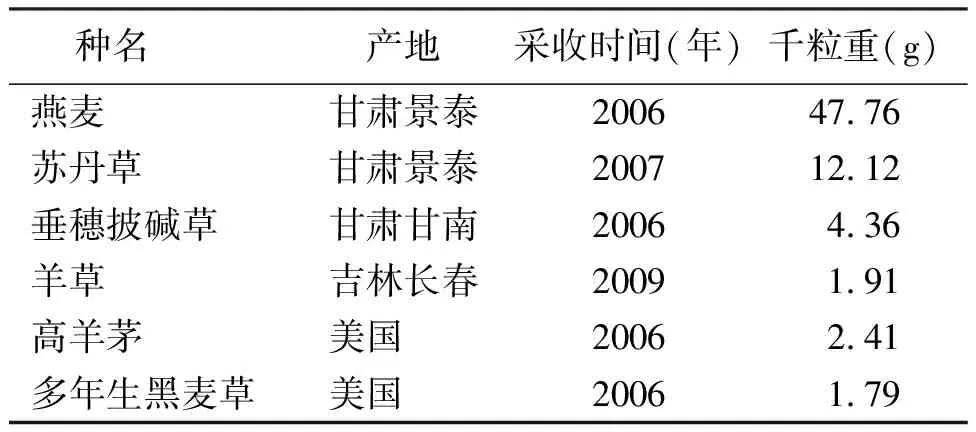
表1 供試種信息
1.2.2發芽試驗 標準發芽試驗參照文獻[5],4個重復,每重復50粒;以胚根突出種皮視為發芽,逐日統計發芽種子數,持續14 d;發芽至第7天,每個培養皿隨機取出7株幼苗,測定其胚根及胚芽長度。計算發芽勢、發芽率、發芽指數和活力指數。
發芽指數(GI)=∑(Gt/Dt);
活力指數(VI)=幼苗長度×發芽指數。
式中,Gt與Dt分別對應當天的發芽種子數與發芽的天數,t=0,1,2,3,…,n。
1.2.3盆栽試驗 對于任一參試物種,每處理設置4個重復,每個重復隨機取50粒種子,均勻播入花盆(直徑15 cm、高15 cm)中,覆蓋1 cm左右相同土質的浮土;每日澆水,以保持花盆內有充足的水分,試驗在蘭州大學草地農業科技學院榆中校區人工智能溫室中進行,溫室內平均日溫度為20 ℃(13~27 ℃);試驗期間逐日統計出苗數,統計時間持續21 d。
出苗速率(GR)=∑100Gi/nti。
式中,n為每個處理中使用的種子個數;Gi為ti天的出苗個數。
1.3數據統計與分析 試驗數據采用SPSS 13.0進行統計分析,ANOVA進行方差分析,采用Duncan法進行多重比較,Excel 2003作圖。
2 結果
2.1老化處理對種子萌發的影響 老化處理降低了除羊草外所有參試種子的發芽勢、發芽率、發芽指數和活力指數,而羊草種子發芽勢、發芽率、發芽指數、活力指數則呈現先升后降的趨勢(表2)。老化處理60 h時,燕麥種子即完全喪失生活力,隨著老化時間的延長(84 h),高羊茅、垂穗披堿草種子相繼喪失生活力,但羊草種子表現出相對較強的耐老化能力,老化84 h后,發芽率仍達34%。
以參試種子的發芽勢、發芽率、發芽指數以及活力指數作為評價指標,可將種批劃分成不同的活力批次:若以發芽率作為標準,可將燕麥劃分為5個批次;以活力指數為指標可劃分為4個批次;但以發芽指數與發芽勢來評價,只能劃分為3個批次。以發芽勢、發芽率、發芽指數以及活力指數任何一個指標均可將羊草劃分為4個批次,垂穗披堿草劃分為5個批次,多年生黑麥草劃分為6個批次。高羊茅種子以發芽率和發芽勢作為評價指標,可將種批劃分為4個批次;但以發芽指數和活力指數作為評價指標,只能劃分為3個批次。
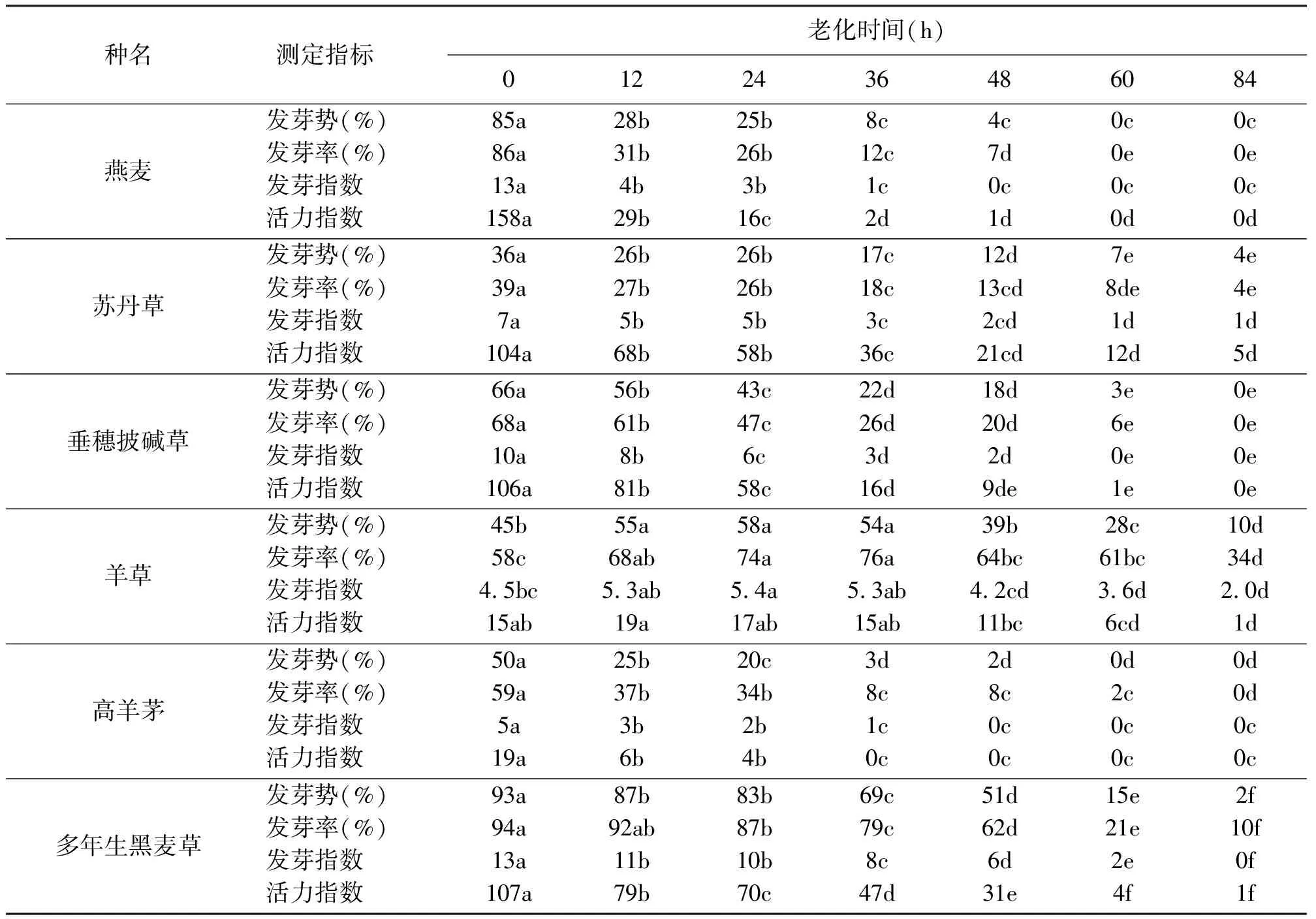
表2 老化處理對種子萌發的影響
2.2老化處理對幼苗的影響 隨老化時間的延長,燕麥、蘇丹草、垂穗披堿草、高羊茅、多年生黑麥草的胚根、胚芽以及幼苗長度呈現降低趨勢;而老化處理時間少于48 h時,羊草的胚根、胚芽以及幼苗長度基本不受老化時間的影響(圖1)。羊草種子老化處理12 h時胚根與幼苗長度最長,分別為1.97與3.53 cm;老化處理24 h胚芽最長,為1.66 cm(圖1)。
2.3老化處理對出苗的影響 燕麥、蘇丹草、垂穗披堿草、高羊茅、多年生黑麥草的出苗率與出苗速率隨著老化程度的加劇而降低(圖2)。但隨著老化時間的增加,羊草種子的出苗率以及出苗速率呈現先升后降的趨勢,老化處理36 h時出苗率達到最大,為94%。
不同參試牧草種子老化處理出苗狀況存在很大差異:燕麥種子老化處理48 h時,出苗率幾乎為0;多樣黑麥草老化處理48 h時,出苗率卻高達59%。參試牧草種子老化處理84 h時,蘇丹草、垂穗披堿草、高羊茅的出苗率幾乎為0,而羊草卻還有31%的出苗率。
2.4相關性分析 除羊草種子外,其他參試牧草種子出苗率和出苗速率均與其發芽勢、發芽率、發芽指數、活力指數以及胚根、胚芽、幼苗長度呈極顯著正相關(P<0.01)(表3)。羊草種子的出苗率與其發芽率存在極顯著正相關(P<0.01),與其發芽勢、發芽指數存在顯著正相關(P<0.05),與其活力指數以及胚根、胚芽、幼苗長度不存在顯著相關性(P>0.05);出苗速率與其發芽勢、發芽率、發芽指數呈極顯著正相關(P<0.01),與其活力指數以及胚根、胚芽、幼苗長度呈顯著正相關(P<0.05)。
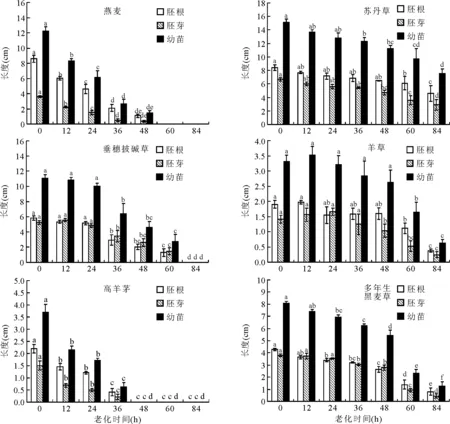
圖1 老化處理對幼苗的影響
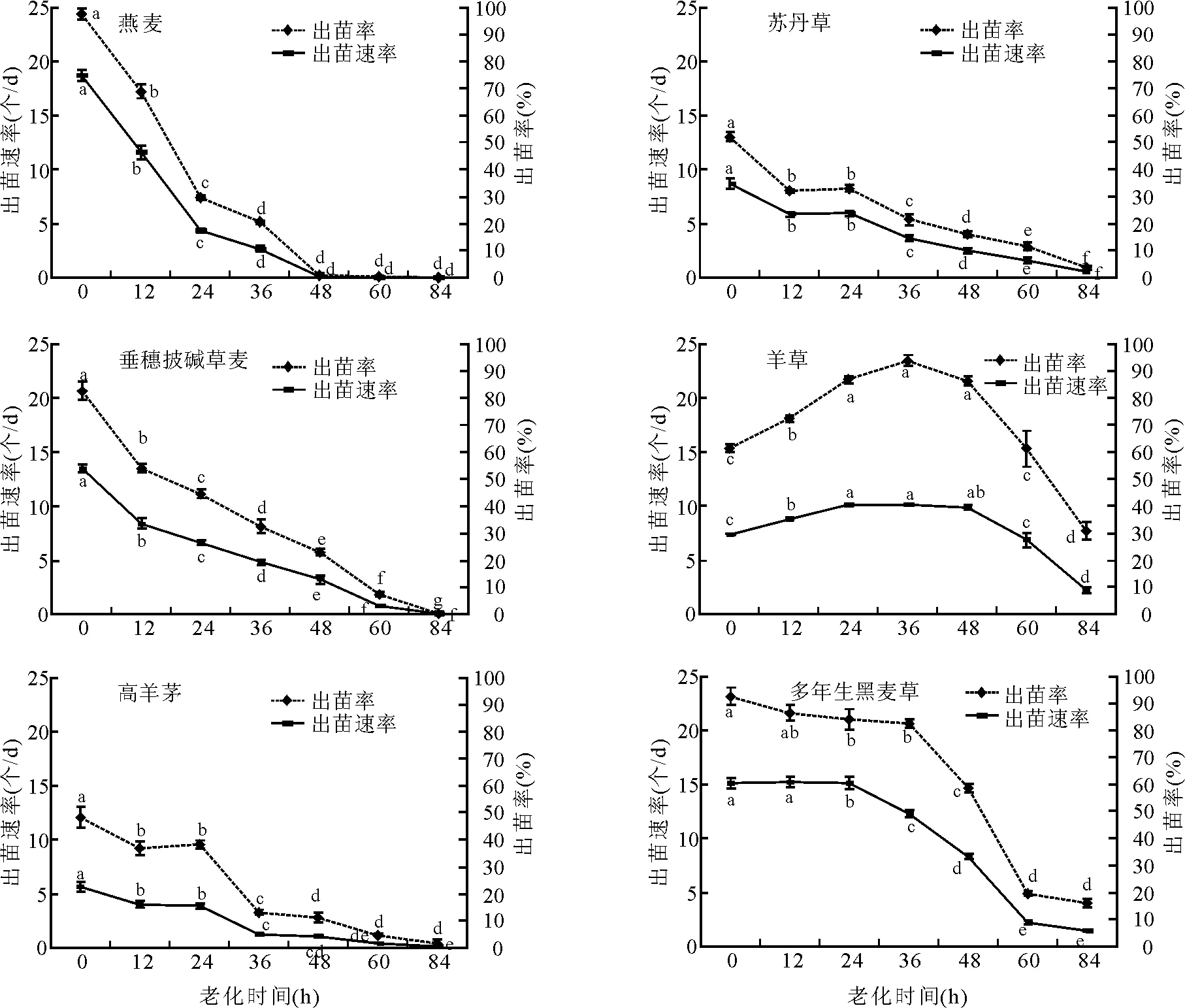
圖2 老化處理對出苗的影響
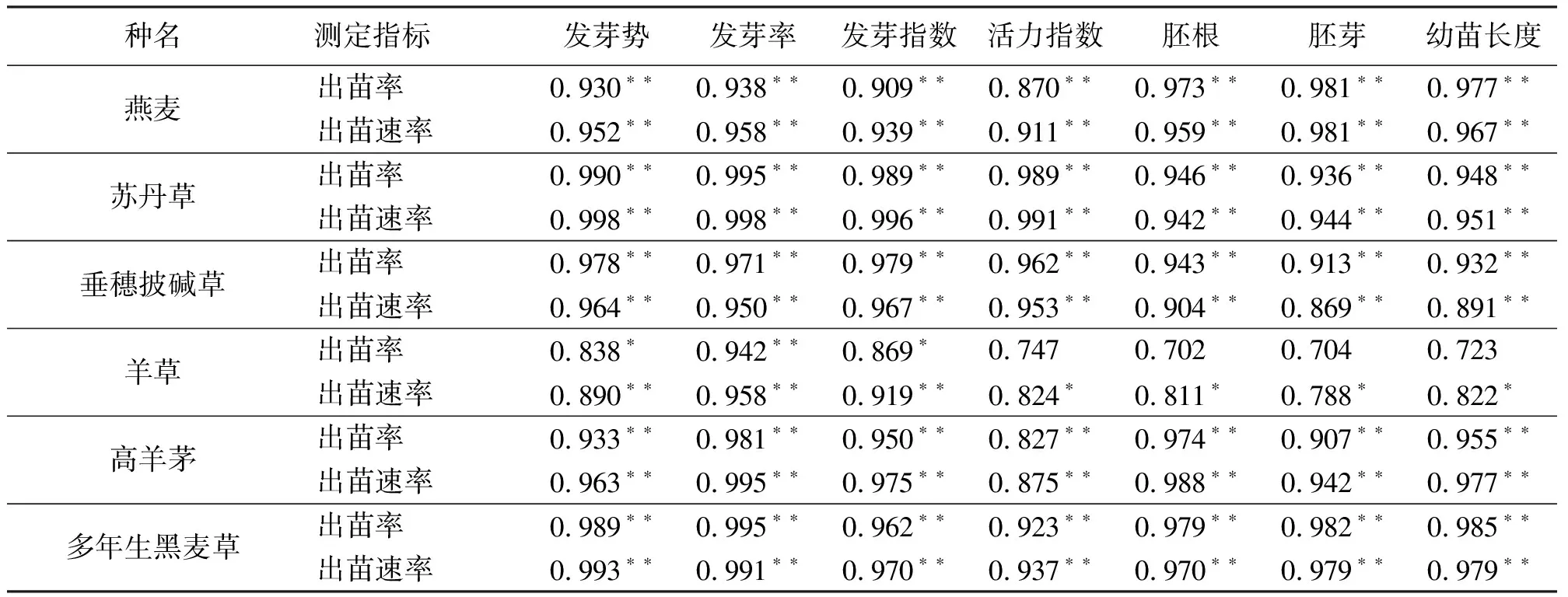
表3 出苗率、出苗速率與種子萌發以及幼苗特性的相關系數
3 討論與結論
大多數種子在儲存過程中發芽率、生活力都有不同程度的降低[6],這與種子儲存過程中的老化有密切的聯系[7]。研究發現,在-13~80 ℃的范圍內,種子的老化規律是一致的[8-10],用人工加速老化法可以模擬種子的自然老化[11-19]。因此利用人工老化處理的方法,可以在很短的時間內獲得不同活力的種批。
本研究通過老化處理幾種禾本科牧草種子,獲得不同活力的種批后,測定其萌發、幼苗以及出苗的相關指標。結果顯示,隨著老化程度的加劇,燕麥、多年生黑麥草、高羊茅、蘇丹草、垂穗披堿草種子的發芽勢、發芽率、發芽指數、活力指數、胚根、胚芽、幼苗長度以及出苗率、出苗速率呈現下降趨勢。這可能是老化處理加快了種子的劣變過程,致使其內部發生變化如膜結構破壞[20]、代謝能力下降[21]、遺傳物質結構變化[22]等,最終導致種子死亡[23]。但另一方面,隨老化的進行,羊草種子的萌發、幼苗以及出苗的相關指標呈現先升后降的趨勢,這可能是因為羊草種子自身具有休眠,一定程度的高溫高濕能夠釋放種子的部分休眠[23],從而提高種子的萌發指標。但隨著老化程度的加劇,劣變速率加快,發芽勢、發芽率、發芽指數、活力指數,胚根、胚芽、幼苗長度,出苗率、出苗速率表現出下降趨勢。毛培勝等[23]的研究亦有相似的觀點。
此外,不同植物種子耐老化程度存在較大差別。如燕麥與多年生黑麥草種子初始發芽率相似,但在老化處理48h時,燕麥種子發芽率只有7%,而多年生黑麥草種子的發芽率仍有62%;老化處理60h,燕麥種子生活力完全喪失,而垂穗披堿草種子發芽率還剩20.5%。這種差異可能與種子的大小、成熟度[24],膜結構,細胞內物質[23]等有關。這也說明在運用老化處理測定種子的活力和預測種子的耐貯性時,老化處理時間應因種而異[25]。
除羊草以外,其他參試種子的發芽勢、發芽率、發芽指數、活力指數,胚根、胚芽、幼苗長度與出苗率、出苗速率均存在極顯著正相關,表明標準發芽試驗相關指標可以很好地預測田間出苗狀況。這與趙光武和王建華[26]、金錫奎和張春慶[27]的研究結果一致,但陳士林的研究表明,標準發芽試驗不能很好地評價種子活力。這可能與研究采用的試驗條件有關,尤其是田間條件,如本研究所采用的田間條件脅迫相對比較溫和,而陳士林[28]則是在實際田間條件下開展的評價工作,環境更為嚴酷,具體原因有待進一步研究。羊草種子的出苗率以及出苗速率與其活力指數,胚根、胚芽、幼苗長度不存在顯著差異,是因為發芽試驗不適合高休眠的種子活力的測定,與王彥榮等[29]的研究一致。
對老化處理獲得的種批進行標準發芽試驗,根據發芽勢、發芽率、發芽指數、活力指數的差異顯著性分析,可以將種批進行分級,相比而言,發芽率能夠更加細致地對其進行分級。
[1]毛培勝,韓建國.貯藏處理對老芒麥種子活力的影響[J].草業科學,2003,20(4):16-19.
[2]王立軍,杜彥斌.利用標準發芽試驗法測定亞麻種子活力[J].種子科技,2010(7):28-29.
[3]WangYR,HamptonJG,HillMJ.Redclovervigourtesting-effectsofthreetestvariables[J].SeedScienceandTechnology,1994,22:99-105.
[4]王彥榮,孫建華,余玲,等.GB/T2930-2001 牧草種子檢驗規程 發芽試驗[S].北京:中國標準出版社,2001.
[5]InternationalSeedTestingAssociation.Internationalrulesforseedtesting[J].SeedScienceandTechnology,1999,27:27-32.
[6]管彥良.種子劣變與膜傷害的關系[J].種子,1991,51:46-47.
[7]朱世東,張志偉.蔬菜種子老化與膜脂過氧化作用[J].園藝學報,1995,22(4):394-396.
[8]EllisRH,HongTD,RobertsEH.ALow-Postire-contentlimittologarithmicrelationsbetweenseedmoisturecontentandlongevity[J].AnnalsofBotany,1988,61:405-408.
[9]WatersC,EngelsJ.Theeffectsofstoringseedsunderextremelydryconditions[J].SeedScienceResearch,1998,8:1-74.
[10]EllisRH,HongTD,RobertsEH.Moisturecontentandthelongevityofseedsofphaseolusvulgaris[J].AnnalsofBotany,1990,66:341-348.
[11]鄭光華.種子活力的原理及其應用[J].植物生理生化進展,1986(4):154-179.
[12]董鴻運.杉木,馬尾松種子人工老化過程中某些生理生化規律的研究[J].種子,1987(2):14-18.
[13]DeloucheJC,BaskinCC.Acceleratedagingtechniquesforpredictingtherelativestorabilityofseedslots[J].SeedScienceandTechnology,1973,1(2):427-452.
[14]PowellAA,MatthowsS.Evaluationofcontrolleddeterioration:anewvigortestforsmallseedvegetables[J].SeedScienceandTechnology,1981,9:633-640.
[15]PowellAA,MatthowsS.Applicationofthecontrolled:deteriorationvigortesttodetectseedlotsofBrussels sproutswithlowpotentialforstorageundercommercialconditions[J].SeedScienceandTechnology,1984,12:649-657.
[16]LikhatecherBS.Changingofviabilityofmaizeseedsinextremeconditionsofstorage[J].ProceedingsofVASXNIL,1977,4:38-68.
[17]LikhateherBS,MusorilaLJ,SchevchenkeZN,et al.Usageofextremeconditionsofseedstorageinmodelingprocessesoftheiraging[J].BulletinofVirus,1978,77:7-62.
[18]LikhatcherBS,ZelenskyGV,KiashkoYG,et al.Modelingofseedaging[J].SeedScienceandTechnology,1984,87:697-704.
[19]MatthewsS.Controlleddeterioration:anewvigortestforcropseeds[A].In:HebblethwaitePD.SeedProduction[M].London:Butterworth,1980:647-660.
[20]李馳,林鳳.種子老化機理的研究進展[J].安徽農業科學,2006,34(20):5176-5177.
[21]韓建國,浦心春,毛培勝.高羊茅種子老化過程中酶活性的變化[J].草地學報,1998,6(2):84-89.
[22]張晗,盧新雄,張志娥,等.種子老化誘導小麥染色體畸變及大麥醇溶蛋白帶型頻率變化的研究[J].植物遺傳資源學報,2004,5(1):56-61.
[23]毛培勝,常淑娟,王玉紅,等.人工老化處理對羊草種子膜透性的影響[J].草業學報,2008,17(6):66-70.
[24]周龍發.種子活力在種子質量評估中重要性的分析[J].農業與技術,2004,24(2):78,87.
[25]劉賀,毛培勝,孔令琪,等.馬棘種子發芽以及人工加速老化測定標準化研究[J].草業科學,2010,27(2):144-149.
[26]趙光武,王建華.甜玉米種子活力測定及其田間成苗能力的評估[J].植物生理學通訊,2005,41(4):444-448.
[27]金錫奎,張春慶.玉米種子活力實用測定方法初探[J].山東農業大學學報,1989(1):36-42.
[28]陳士林.玉米種子活力與田間苗期性狀相關性研究[J].種子,2003(4):35-37.
[29]王彥榮,余玲,劉友良,等.用活力指標評價幾種牧草的種批質量和田間出苗率[J].草業學報,2003,12(5):62-69.
